
Dendrometric Relationships and Biomass in Commercial Plantations of Dipteryx spp. in the Eastern Amazon

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
Study and Data Collection Locations
3. Statistical Analysis
R2adj. = 0.891
R2adj. = 0.996
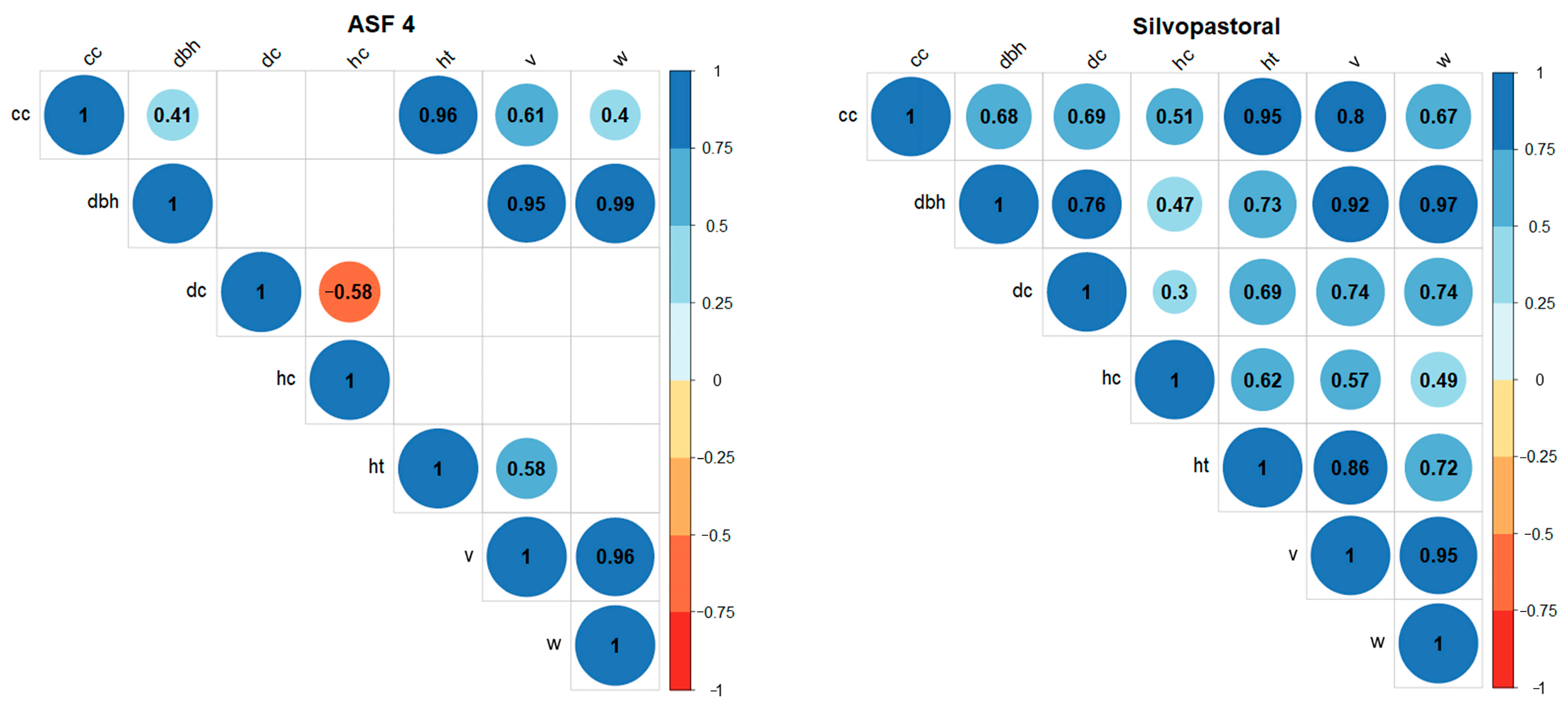
4. Results
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Instituto Brasileiro de Geografia e Estatística (IBGE). Mapa da Amazônia Legal 2022; IBGE: Rio de Janeiro, Brazil, 2021.
- Instituto Brasileiro de Geografia e Estatística (IBGE). Sistema IBGE de Recuperação Automática—SIDRA; IBGE: Rio de Janeiro, Brazil, 2021.
- Peres, C.A.; Campos-Silva, J.; Ritter, C.D. Environmental policy at a critical junction in the Brazilian Amazon. Trends Ecol. Evol. 2023, 38, 113–116. [Google Scholar] [CrossRef]
- Fearnside, P.M. Valoração do estoque de serviços ambientais como estratégia de desenvolvimento no Estado do Amazonas. Inclusão Soc. 2018, 12, 141–151. [Google Scholar]
- Homma, A.K.O. Amazônia: Venda de Serviços Ambientais ou de Atividades Produtivas? Terceira Margem Amaz. 2021, 6, 23–34. [Google Scholar] [CrossRef]
- Lapola, D.M.; Pinho, P.; Barlow, J.; Aragão, L.E.O.C.; Berenguer, E.; Carmenta, R.; Liddy, H.M.; Seixas, H.; Silva, C.V.J.; Silva-Junior, C.H.L.; et al. The drivers and impacts of Amazon forest degradation. Science 2023, 379, eabp8622. [Google Scholar] [CrossRef] [PubMed]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef]
- da Cruz, D.C.; Ferreira, G.C.; Ribeiro, S.S.; Schwartz, G.; Monteiro, A. Priority areas for restoration in permanent preserrmedvation areas of rural properties in the Brazilian Amazon. Land Use Policy 2022, 115, 106030. [Google Scholar] [CrossRef]
- Nunes, C.A.; Berenguer, E.; Frana, F.; Ferreira, J.; Lees, A.C.; Louzada, J.; Sayer, E.; Solar, R.; Smth, C.C.; Aragao, L.E.O.C.; et al. Linking land-use and land-cover transitions to their ecological impact in the Amazon. Proc. Natl. Acad. Sci. USA 2022, 119, e2202310119. [Google Scholar] [CrossRef]
- Sonter, L.J.; Herrera, D.; Barrett, D.J.; Galford, G.L.; Moran, C.J.; Soares-Filho, B.S. Mining drives extensive deforestation in the Brazilian Amazon. Nat. Commun. 2017, 8, 1013. [Google Scholar] [CrossRef]
- Vasconcelos, A.Í.T.; Garcia, E.A.d.R.; Furtado, C.F.C.; Cabral, J.E.D.O. As dimensões da sustentabilidade dos Sistemas Agroflorestais—SAFs: Um estudo no Projeto de Reflorestamento Consorciado e Adensado—RECA, Ponta do Abunã—RO. Desenvolv. Meio Ambiente 2016, 36, 73–93. [Google Scholar] [CrossRef]
- da Cruz, D.C.; Benayas, J.M.R.; Ferreira, G.C.; Santos, S.R.; Schwartz, G. An overview of forest loss and restoration in the Brazilian Amazon. New For. 2021, 52, 1–16. [Google Scholar] [CrossRef]
- BRASIL. Lei nº 12.651, de 25 de maio de 2012. Dispõe sobre a proteção da vegetação nativa; altera as Leis nºs 6.938, de 31 de agosto de 1981, 9.393, de 19 de dezembro de 1996, e 11.428, de 22 de dezembro de 2006; revoga as Leis nºs 4.771, de 15 de setembro de 1965, e 7.754, de 14 de abril de 1989, e a Medida Provisória nº 2.166-67, de 24 de agosto de 2001; e dá outras providências. Available online: https://www.planalto.gov.br/ccivil_03/_ato2011-2014/2012/lei/l12651.htm (accessed on 20 June 2023).
- Villa, P.M.; Martins, S.V.; de Oliveira Neto, S.N.; Rodrigues, A.C.; Hernández, E.P.; Kim, D.G. Policy forum: Shifting cultivation and agroforestry in the Amazon: Premises for REDD+. For. Policy Econ. 2020, 118, 102217. [Google Scholar] [CrossRef]
- de Vasconcellos, R.C.; Beltrão, N.E.S. Avaliação de prestação de serviços ecossistêmicos em sistemas agroflorestais através de indicadores ambientais. Interações 2018, 19, 209–220. [Google Scholar] [CrossRef]
- da Silva, J.A.G.; Pauletto, D.; da Silva, A.F.; Carvalho, C.D.S.d.S.; Nascimento, H.G.G.D. Morfometria de Plantios de Dipteryx odorata Aubl Willd (Cumaru) no Oeste do Pará. Adv. For. Sci. 2020, 7, 1171–1180. [Google Scholar] [CrossRef]
- Capucho, H.L.V.; Silva, A.F.d.S.; Rebelo, A.G.d.M.; Pauletto, D.; Silva, A.R. Phenology of Dipteryx odorata and Dipteryx punctata in agroforestry systems in the eastern Amazon. Amaz. J. Agric. Environ. Sci. 2021, 64, 1–16. [Google Scholar]
- Román-Dañobeytia, F.; Cabanillas, F.; Lefebvre, D.; Farfan, J.; Alferez, J.; Polo-Villanueva, F.; Llacsahuanga, J.; Vega, C.M.; Velasquez, M.; Corvera, R.; et al. Survival and early growth of 51 tropical tree species in areas degraded by artisanal gold mining in the Peruvian Amazon. Ecol. Eng. 2021, 159, 106097. [Google Scholar] [CrossRef]
- da Mota, C.G.; Pauletto, D.; Capucho, H.L.V.; da Silva, S.U.P.; Ponte, M.X. O cultivo do cumaru como alternativa econômica para agricultores familiares: Estudo de caso na região oeste do Pará. Res. Soc. Dev. 2022, 11, e46511326732. [Google Scholar] [CrossRef]
- de Melo, R.R.; Dacroce, J.M.F.; Junior, F.R.; Lisboa, G.d.S.; França, L.C.d.J. Lumber yield of four native forest species of the Amazon Region. Floresta E Ambiente 2019, 26, e20160311. [Google Scholar] [CrossRef]
- de Sousa, B.C.M.; de Castro, S.P.; Lourido, K.A.; Kasper, A.A.M.; Paulino, G.d.S.; Delarmelina, C.; Duarte, M.C.T.; Sartoratto, A.; Vieira, T.A.; Lustosa, D.C.; et al. Identification of Coumarins and Antimicrobial Potential of Ethanolic Extracts of Dipteryx odorata and Dipteryx punctata. Molecules 2022, 27, 5837. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.C.C.; Leite, H.G. Mensuração Florestal: Perguntas e Respostas, 5th ed.; Editora UFV: Viçosa, Brazil, 2017. [Google Scholar]
- Binoti, D.H.B.; Da Binoti, M.L.M.S.; Leite, H.G.; Silva, A. Redução dos custos em inventário de povoamentos equiâneos. Rev. Bras. Cienc. Agrar. 2013, 8, 125–129. [Google Scholar] [CrossRef]
- Lopes, L.S.S.; Rode, R.; Pauletto, D.; Baloneque, D.D.; dos Santos, F.G.; Silva, A.R.; Binoti, D.H.B.; Leite, H.G. Uso de regressão e redes neurais artificiais na estimativa do volume de Khaya ivorensis. Rev. Ciência Madeira—RCM 2020, 11, 74–84. [Google Scholar] [CrossRef]
- Phillips, O.L.; Brienen, R.J.W. Carbon uptake by mature Amazon forests has mitigated Amazon nations carbon emissions. Carbon Balance Manag. 2017, 12, 2–9. [Google Scholar] [CrossRef]
- Heinrich, V.H.A.; Dalagnol, R.; Cassol, H.L.G.; Rosan, T.M.; de Almeida, C.T.; Silva Junior, C.H.L.; Campanharo, W.A.; House, J.I.; Sitch, S.; Hales, T.C.; et al. Large Carbon Sink Potential of Secondary Forests in the Brazilian Amazon to Mitigate Climate Change. Nat. Commun. 2021, 12, 1785. [Google Scholar] [CrossRef]
- Araujo, E.C.G.; Sanquetta, C.R.; Dalla Corte, A.P.; Pelissari, A.L.; Orso, G.A.; Silva, T.C. Global Review and State-of-the-Art of Biomass and Carbon Stock in the Amazon. J. Environ. Manag. 2023, 331, 117251. [Google Scholar] [CrossRef]
- Ministério do Meio Ambiente (MMA). ENREDD+ Estratégia Nacional para Redução das Emissões Provenientes do Desmatamento e da Degradação Florestal, Conservação dos Estoques de Carbono Florestal, Manejo Sustentável de Florestas e Aumento de Estoques de Carbono Florestal. 2016. Available online: http://redd.mma.gov.br/en/redd-and-brazil-s-ndc (accessed on 7 May 2023).
- Romero, F.M.B.; Jacovine, L.A.G.; Ribeiro, S.C.; Neto, J.A.F.; Ferrante, L.; da Rocha, S.J.S.S.; Torres, C.M.M.E.; Junior, V.T.M.d.M.; Gaspar, R.d.O.; Velasquez, S.I.S.; et al. Stocks of carbon in logs and timber products from forest management in the southwestern Amazon. Forests 2020, 11, 1113. [Google Scholar] [CrossRef]
- Isinkaralar, O.; Isinkaralar, K.; Yilmaz, D. Climate-related spatial reduction risk of agricultural lands on the Mediterranean coast in Türkiye and scenario-based modelling of urban growth. Environ. Dev. Sustain. 2023, 25, 13199–13217. [Google Scholar] [CrossRef]
- Isinkaralar, O. Bioclimatic comfort in urban planning and modeling spatial change during 2020–2100 according to climate change scenarios in Kocaeli, Türkiye. Int. J. Environ. Sci. Technol. 2023, 20, 7775–7786. [Google Scholar] [CrossRef]
- Zilli, M.; Scarabello, M.; Soterroni, A.C.; Valin, H.; Mosnier, A.; Leclère, D.; Havlík, P.; Kraxner, F.; Lopes, M.A.; Ramos, F.M. The impact of climate change on Brazil’s agriculture. Sci. Total Environ. 2020, 740, 139384. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.A.; de Paiva, H.N.; Nogueira, G.S.; Leite, H.G. Planejamento de Experimentos Com Espécies Florestais, 1st ed.; Editora UFV: Viçosa, Brazil, 2022. [Google Scholar]
- Lima, Q.T.; Cunha, T.A.; Amaro, M.A.; Figueiredo, E.O.; Parente, P.R.F. Volume estimate for three timber species with commercial interest from the diameter of the stump. Floresta 2021, 51, 776–784. [Google Scholar] [CrossRef]
- Fetzer, D.E.F.; Kanda, L.R.S.; Xavier, L.A.; Cruz, P.N.; Errico, M.; Corazza, M.L. Lipids and coumarin extraction from cumaru seeds (Dipteryx odorata) using sequential supercritical CO2+solvent and pressurized etanol. J. Supercrit. Fluids 2022, 188, 105688. [Google Scholar] [CrossRef]
- Moraes, M.L.L.; Silva, S.A.; Silva, H.D.T. Experimental mixture design as a tool to evaluate coumarin (1,2-benzopyrone) extraction from Dipteryx odorata seeds. J. Pharm. Biomed. Anal. 2022, 210, 114586. [Google Scholar] [CrossRef]
- Lameira, M.K.d.S.; Silva, H.K.M.; Gomes, K.M.A.; Cândido, A.C.T.F.; Martorano, L.G.; Ribeiro, R.B.d.S.; Silva, A.R.; Gama, J.R.V. Capacidade produtiva de Dipteryx odorata (Aubl.) Willd. em um sistema de produção de pecuária integrada no Baixo Amazonas. Bras. Product. 2022, 19, 56–65. [Google Scholar]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.d.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Martorano, L.G.; Soares, W.B.; Moraes, J.R.d.S.C.d.; Nascimento, W.; Aparecido, L.E.d.O.; Villa, P.M. Climatology of air temperature in Belterra: Thermal regulation ecosystem services provided by the Tapajós National Forest in the amazon. Rev. Bras. De Meteorol. 2021, 36, 327–337. [Google Scholar] [CrossRef]
- Brasil, E.C.; Cravo, M.S.; Viégas, I.J.M. Recomendações de adubação para o estado do Pará. In Recomendações de Calagem e Adubação Para o Estado do Pará; EMBRAPA: Brasília, Brazil, 2020. [Google Scholar]
- Instituto Brasileiro de Geografia e Estatística (IBGE). Mapa de Vegetação do Brasil; IBGE: Rio de Janeiro, Brazil, 2004.
- Baloneque, D.D.; Pauletto, D.; Lopes, L.S.S.; Rode, R.; Oliveira, T.G.d.S. Variáveis morfométricas de quatro espécies florestais em sistema silvipastoril no município de Belterra, Pará. Res. Soc. Dev. 2022, 11, e43511326761. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Araújo, S.O.; Vital, B.R.; Oliveira, B.; Carneiro, A.C.O.; Lourenço, A.; Pereira, H. Physical and mechanical properties of heat treated wood from Aspidosperma populifolium, Dipteryx odorata and Mimosa scabrella. Maderas. Cienc. Tecnol. 2016, 18, 143–156. [Google Scholar]
- Zau, M.D.L.; De Vasconcelos, R.P.; Giacon, V.M.; Lahr, F.A.R. Chemical, physical and mechanical properties of particleboard produced with Amazon wood waste—Cumaru (Dipteryx odorata)—and Castor oil based polyurethane adhesive. Polimeros 2014, 24, 726–732. [Google Scholar] [CrossRef]
- Romero, F.M.B.; Jacovine, L.A.G.; Ribeiro, S.C.; Torres, C.M.M.E.; da Silva, L.F.; Gaspar, R.d.O.; da Rocha, S.J.S.S.; Staudhammer, C.L.; Fearnside, P.M. Allometric equations for volume, biomass, and carbon in commercial stems harvested in a managed forest in the southwestern amazon: A case study. Forests 2020, 11, 874. [Google Scholar] [CrossRef]
- de Souza, A.L.; Soares, C.P.B. Florestas Nativas: Estrutura, Dinâmica e Manejo, 1st ed.; Editora UFV: Viçosa, Brazil, 2013. [Google Scholar]
- Santos, A.C.d.A.; Fardin, L.P.; de Oliveira Neto, R.R. Teste de hipótese em análise de regressão. In Jurnal Penelitian Pendidikan Guru Sekolah Dasar, 1st ed.; Novas Edições Acadêmicas: São Paulo, Brazil, 2017. [Google Scholar]
- Binkley, D.; Stape, J.L.; Bauerle, W.L.; Ryan, M.G. Explaining growth of individual trees: Light interception and efficiency of light use by Eucalyptus at four sites in Brazil. For. Ecol. Manag. 2010, 259, 1704–1713. [Google Scholar] [CrossRef]
- Wink, C.; Monteiro, J.S.; Reinert, D.J.; Liberalesso, E.E. Parâmetros da copa e a sua relação com o diâmetro e altura das árvores de eucalipto em diferentes idades. Sci. For./For. Sci. 2012, 40, 57–67. [Google Scholar]
- Li, Q.; Liu, Z.; Jin, G. Impacts of stand density on tree crown structure and biomass: A global meta-analysis. Agric. For. Meteorol. 2022, 326, 109181. [Google Scholar] [CrossRef]
- de Oliveira, T.M.; Madi, J.P.S.; Cerqueira, C.L.; Millikan, P.H.K.; Chaves, M.P.d.L.; Carvalho, S.d.P.C. Relações morfométricas para árvores de Tectona grandis. Adv. For. Sci. 2018, 5, 461–465. [Google Scholar]
- Resende, R.T.; Soares, A.A.V.; Forrester, D.I.; Marcatti, G.E.; dos Santos, A.R.; Takahashi, E.K.; e Silva, F.F.; Grattapaglia, D.; Resende, M.D.V.; Leite, H.G. Environmental uniformity, site quality and tree competition interact to determine stand productivity of clonal Eucalyptus. For. Ecol. Manag. 2018, 410, 76–83. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Chen, Q.; Duan, G.; Liu, Q.; Ye, Q.; Sharma, R.P.; Chen, Y.; Liu, H.; Fu, L. Estimating crown width in degraded forest: A two-level nonlinear mixed-effects crown width model for Dacrydium pierrei and Podocarpus imbricatus in tropical China. For. Ecol. Manag. 2021, 497, 119486. [Google Scholar] [CrossRef]
- Ricken, P.; Hess, A.F.; de Mattos, P.P.; Braz, E.M.; Nakajima, N.Y.; Hosokawa, R.T. Morfometria de Araucaria angustifolia em diferentes altitudes no Sul do Brasil. Pesqui. Florest. Bras. 2020, 40, 1–11. [Google Scholar] [CrossRef]
- Chai, G.; Zheng, Y.; Lei, L.; Yao, Z.; Chen, M.; Zhang, X. A novel solution for extracting individual tree crown parameters in high-density plantation considering inter-tree growth competition using terrestrial close-range scanning and photogrammetry technology. Comput. Electron. Agric. 2023, 209, 107849. [Google Scholar] [CrossRef]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; del Río, M.; Drössler, L.; Heym, M.; et al. Effects of crown architecture and stand structure on light absorption in mixed and monospecific Fagus sylvatica and Pinus sylvestris forests along a productivity and climate gradient through Europe. J. Ecol. 2018, 106, 746–760. [Google Scholar] [CrossRef]
- Qin, Y.; Wu, B.; Lei, X.; Feng, L. Prediction of tree crown width in natural mixed forests using deep learning algorithm. For. Ecosyst. 2023, 10, 100109. [Google Scholar] [CrossRef]
- Mildrexler, D.J.; Berner, L.T.; Law, B.E.; Birdsey, R.A.; Moomaw, W.R. Large Trees Dominate Carbon Storage in Forests East of the Cascade Crest in the United States Pacific Northwest. Front. For. Glob. Chang. 2020, 3, 594274. [Google Scholar] [CrossRef]
- Shoda, T.; Imanishi, J.; Shibata, S. Growth characteristics and growth equations of the diameter at breast height using tree ring measurements of street trees in Kyoto City, Japan. Urban For. Urban Green. 2020, 49, 126627. [Google Scholar] [CrossRef]
- Qin, Y.; He, X.; Lei, X.; Feng, L.; Zhou, Z.; Lu, J. Tree size inequality and competition effects on nonlinear mixed effects crown width model for natural spruce-fir-broadleaf mixed forest in northeast China. For. Ecol. Manag. 2022, 518, 120291. [Google Scholar] [CrossRef]
- Hu, L.; Xu, X.; Wang, J.; Xu, H. Individual tree crown width detection from unmanned aerial vehicle images using a revised local transect method. Ecol. Inform. 2023, 75, 102086. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Banin, L.; Phillips, O.L.; Baker, T.R.; Lewis, S.L.; Quesada, C.A.; Affum-Baffoe, K.; Arets, E.J.M.M.; Berry, N.J.; Bird, M.; et al. Height-diameter allometry of tropical forest trees. Biogeosciences 2011, 8, 1081–1106. [Google Scholar] [CrossRef]
- Nascimento, R.G.M.; Vanclay, J.K.; Filho, A.F.; Machado, S.D.A.; Ruschel, A.R.; Hiramatsu, N.A.; de Freitas, L.J.M. The tree height estimated by non-power models on volumetric models provides reliable predictions of wood volume: The Amazon species height modelling issue. Trees For. People 2020, 2, 100028. [Google Scholar] [CrossRef]
- Leites, L.P.; Zubizarreta-Gerendiain, A.; Robinson, A.P. Modeling mensurational relationships of plantation-grown loblolly pine (Pinus taeda L.) in Uruguay. For. Ecol. Manag. 2013, 289, 455–462. [Google Scholar] [CrossRef]
- Gujarati, D.N.; Porter, D.C. Econometria Básica, 5th ed.; AMGH Editora: Porto Alegre, Brazil, 2011. [Google Scholar]
- Hair, J.F., Jr.; Anderson, R.E.; Tatham, R.L.; Blanck, W.C. Análise Multivariada de Dados, 5th ed.; Bookman: Los Angeles, CA, USA, 2005. [Google Scholar]
- Voss, D.S. Multicollinearity. In Encyclopedia of Social Measurement; Elsevier: Amsterdam, The Netherlands, 2004; Volume 2, pp. 1–3000. [Google Scholar]
- de Oliveira, M.L.R.; Leite, H.G.; Garcia, S.L.R.; Campos, J.C.C.; Soares, C.P.B.; Santana, R.C. Estimação do volume de árvores de clones de eucalipto pelo método da similaridade de perfis. Rev. Árvore 2009, 33, 133–141. [Google Scholar] [CrossRef]
- Reis, T.; Silva, S.; Junior, I.T.; Leite, R.; Taquetti, M.; Atanazio, K.; Fardin, L.; Cruz, J.; Leite, H. Assessing alternatives for growth and yield modelling in eucalypt stands. South. For. 2022, 84, 34–43. [Google Scholar] [CrossRef]
- Gomes, G.S.L.; Neto, S.N.d.O.; Leite, H.G.; da Silva, M.L.; Lopes, L.S.d.S.; Said Schettini, B.L. Relationships between spacing, productivity and profitability of eucalypt plantations in a small rural property in south-eastern Brazil. South. For. 2022, 84, 206–214. [Google Scholar] [CrossRef]
- Araujo, H.J.B. Inventário florestal a 100% em pequenas áreas sob manejo florestal madeireiro. Acta Amaz. 2006, 36, 447–464. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, S.; Xu, A.; Chen, B. Passive measurement method of tree diameter at breast height using a smartphone. Comput. Electron. Agric. 2019, 163, 104875. [Google Scholar] [CrossRef]
- Curto, R.D.A.; Pinto, M.F.; Wink, C.; de Araújo, E.J.G.; Kohler, S.V. Aplicativos e smartphones para mensuração da altura de árvores em plantio florestal. Pesqui. Florest. Bras. 2022, 42, 1–12. [Google Scholar] [CrossRef]
- Chisholm, R.A.; Muller-Landau, H.C.; Abdul Rahman, K.; Bebber, D.P.; Bin, Y.; Bohlman, S.A.; Bourg, N.A.; Brinks, J.; Bunyavejchewin, S.; Butt, N.; et al. Scale-dependent relationships between tree species richness and ecosystem function in forests. J. Ecol. 2013, 101, 1214–1224. [Google Scholar] [CrossRef]
- Castro-Izaguirre, N.; Chi, X.; Baruffol, M.; Tang, Z.; Ma, K.; Schmid, B.; Niklaus, P.A. Tree diversity enhances stand carbon storage but not leaf area in a subtropical forest. PLoS ONE 2016, 11, e0167771. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.P.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G.; et al. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef]
- Liu, X.; Trogisch, S.; He, J.S.; Niklaus, P.A.; Bruelheide, H.; Tang, Z.; Erfmeier, A.; Scherer-Lorenzen, M.; Pietsch, K.A.; Yang, B.; et al. Tree species richness increases ecosystem carbon storage in subtropical forests. Proc. R. Soc. B Biol. Sci. 2018, 285, 20182090. [Google Scholar]
- Celentano, D.; Rousseau, G.X.; Paixão, L.S.; Lourenço, F.; Cardozo, E.G.; Rodrigues, T.O.; Silva, H.R.; Medina, J.; Sousa, T.M.C.; Rocha, A.E.; et al. Carbon sequestration and nutrient cycling in agroforestry systems on degraded soils of Eastern Amazon, Brazil. Agrofor. Syst. 2020, 94, 1781–1792. [Google Scholar] [CrossRef]
- Chen, Q.; Lu, D.; Keller, M.; Dos-Santos, M.N.; Bolfe, E.L.; Feng, Y.; Wang, C. Modeling and Mapping Agroforestry Aboveground Biomass in the Brazilian Amazon Using Airborne Lidar Data. Remote Sens. 2016, 8, 21. [Google Scholar] [CrossRef]
- Lenci, L.H.V.; Tsukamoto Filho, A.A.; Weber, O.L.S.; Honoré, E.A.D.; Caldeira, S.F.; Bertolini, J. Estoque de carbono da biomassa arbórea, serapilheira e solo em sistemas agroflorestais com seringueira. Rev. Bras. Ciências Agrárias 2022, 17, e2406. [Google Scholar]
- Neuman, A.D.; Belcher, K.W. The contribution of carbon-based payments to wetland conservation compensation on agricultural landscapes. Agric. Syst. 2011, 104, 75–81. [Google Scholar] [CrossRef]
- Manzatto, C.V.; Skorupa, L.A.; de Araújo, L.S.; Vicente, L.E.; Assad, E.D. Estimativas de redução de emissões de gases de efeito estufa pela adoção de sistemas ILPF no Brasil. In Sistemas de Integração Lavoura-Pecuária-Floresta no Brasil: Estratégias Regionais de Transferência de Tecnologia, Avaliação da Adoção e de Impactos; EMBRAPA: Brasília, Brazil, 2019; pp. 400–424. [Google Scholar]
- Cardozo, E.G.; Celentano, D.; Rousseau, G.X.; e Silva, H.R.; Muchavisoy, H.M.; Gehring, C. Agroforestry systems recover tree carbon stock faster than natural succession in Eastern Amazon, Brazil. Agrofor. Syst. 2022, 96, 941–956. [Google Scholar] [CrossRef]
- Müller, M.D.; Fernandes, E.N.; Castro, C.R.T.; Paciullo, D.S.C.; Alves, F.D.F. Estimativa de Acúmulo de Biomassa e Carbono em Sistema Agrossilvipastoril na Zona da Mata Mineira. Pesqui. Florest. Bras. 2010, 11–18. [Google Scholar] [CrossRef]
- dos Santos, S.R.M.; Miranda, I.d.S.; Tourinho, M.M. Análise florística e estrutural de sistemas agroflorestais das várzeas do rio Juba, Cametá, Pará. Acta Amaz. 2004, 34, 251–263. [Google Scholar] [CrossRef]
- Montagnini, F.; Nair, P.K.R. Carbon sequestration: An underexploited environmental benefit of agroforestry systems. Agrofor. Syst. 2004, 61–62, 281–295. [Google Scholar]
- BRASIL. Lei nº 14.119, de 13 de Janeiro de 2021. Institui a Política Nacional de Pagamento por SA; e Altera as Leis nos 8.212, de 24 de Julho de 1991, 8.629, de 25 de Fevereiro de 1993, e 6.015, de 31 de Dezembro de 1973, Para Adequá-Las à Nova Política. Available online: http://www.planalto.gov.br/ccivil_03/_ato2019-2022/2021/lei/L14119.htm (accessed on 24 June 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Stand | City | Age (Years) | Spacing (m) | Species Cultivated/Raised in Consortium * |
|---|---|---|---|---|
| Homogeneous | Alenquer | 10 | 6 × 6 | - |
| AFS 1 | Alenquer | 6 | 5.5 × 4.5 | Ab |
| AFS 2 | Belterra | 7 | 8 × 6 | C; Aç; Gr; Bn; P |
| AFS 3 | Alenquer | 8 | 10 × 8.5 | Lm |
| AFS 4 | Mojuí dos Campos | 9 | 4 × 8 | Lj |
| Silvopastoral | Belterra | 11 | 10 × 10 | Bovines |
| Forest Stand | n | ht (m) | hc (m) | cc (m) | dbh (cm) | dc (m) |
|---|---|---|---|---|---|---|
| Homogeneous | 25 | 7.04 ± 1.43 | 1.9 ± 0.73 | 4.48 ± 1.34 | 8.79 ± 2.86 | 4.1 ± 1.26 |
| AFS 1 | 25 | 8.08 ± 0.78 | 0.64 ± 0.47 | 6.03 ± 0.93 | 8.88 ± 1.51 | 5.15 ± 0.77 |
| AFS 2 | 25 | 9.68 ± 1.94 | 2.78 ± 1.1 | 6.22 ± 2.03 | 11.95 ± 3.31 | 4.7 ± 1.57 |
| AFS 3 | 25 | 5.26 ± 1.4 | 1.26 ± 0.84 | 2.74 ± 1.42 | 6.18 ± 2.78 | 3.15 ± 1.54 |
| AFS 4 | 25 | 9.62 ± 1.38 | 0.7 ± 0.35 | 7.83 ± 1.2 | 14.09 ± 2.6 | 7.25 ± 0.76 |
| AFS 1 and 4 | 50 | 8.85 ± 1.35 | 0.67 ± 0.41 | 6.93 ± 1.4 | 11.48 ± 3.36 | 6.2 ± 1.3 |
| Silvopastoral | 50 | 14.35 ± 4.55 | 5.45 ± 2.58 | 10.45 ± 3.77 | 16.57 ± 5.31 | 7.97 ± 3.16 |
| Total sample | 175 | 9.77 ± 4.2 | 2.6 ± 2.45 | 6.88 ± 3.56 | 11.86 ± 5.22 | 5.76 ± 2.68 |
| Model | System | R2adj. | F | ||||
|---|---|---|---|---|---|---|---|
| Homogeneous | 0.857056 * | 0.50536 * | 0.589 | 2.500 | 34.556 | ||
| AFS 1 | 1.382878 * | 0.32745 * | 0.358 | 1.131 | 28.310 | ||
| AFS 2 | 0.839736 * | 0.57845 * | 0.696 | 2.924 | 73.442 | ||
| AFS 3 | 0.857707 * | 0.446987 * | 0.698 | 2.159 | 35.696 | ||
| AFS 4 | 1.382878 * | 0.32745 * | 0.358 | 1.131 | 28.310 | ||
| Silvopastoral | 0.762366 * | 0.671219 * | 0.516 | 23.599 | 50.384 | ||
| General model | −0.584886 * | 1.010811 * | 0.713 | 9.497 | 359.300 | ||
| Homogeneous | 0.921099 * | 0.353371 * | 0.19068 ns | 0.617 | 1.139 | 20.344 | |
| AFS 1 | 1.303266 * | 0.219919 * | 0.187483 ns | 0.378 | 1.129 | 15.880 | |
| AFS 2 | 1.0934 * | 0.370159 * | 0.172467 ns | 0.767 | 1.117 | 40.406 | |
| AFS 3 | 0.922075 * | 0.204625 ns | 0.33709 * | 0.708 | 1.170 | 30.078 | |
| AFS 4 | 1.303266 * | 0.219919 * | 0.187483 ns | 0.378 | 1.129 | 15.880 | |
| Silvopastoral | 1.030821 * | 0.251216 ns | 0.447024 * | 0.636 | 1.265 | 43.872 | |
| General model | 0.617836 * | 0.458415 * | 0.300522 * | 0.760 | 1.231 | 275.800 |
| Model | Sistem | R2adj. | F | ||||
|---|---|---|---|---|---|---|---|
| Homogeneous | −0.335868 ns | 0.797088 * | 0.507 | 1.282 | 25.666 | ||
| AFS 1 | 0.42463 * | 0.57355 * | 0.544 | 1.161 | 59.440 | ||
| AFS 2 | −1.470793 * | 1.207716 * | 0.793 | 1.236 | 92.693 | ||
| AFS 3 | −0.190952 ns | 0.718981 * | 0.573 | 1.364 | 33.147 | ||
| AFS 4 | 0.42463 * | 0.57355 * | 0.544 | 1.161 | 59.440 | ||
| Silvopastoral | −0.600536 ns | 0.939553 * | 0.582 | 1.390 | 69.203 | ||
| General model | −0.4364 * | 0.8743 * | 0.699 | 1.331 | 404.400 | ||
| Homogeneous | −0.883262 ns | 0.474319 ns | 0.638691 ns | 0.547 | 1.269 | 15.503 | |
| AFS 1 | 0.045817 ns | 0.483848 * | 0.273933 ns | 0.558 | 1.158 | 31.950 | |
| AFS 2 | −1.970995 * | 0.863153 * | 0.595666 ns | 0.805 | 1.228 | 50.667 | |
| AFS 3 | −0.996543 * | 0.299154 ns | 0.939239 * | 0.695 | 1.300 | 28.293 | |
| AFS 4 | 0.045817 ns | 0.483848 * | 0.273933 ns | 0.558 | 1.158 | 31.950 | |
| Silvopastoral | −1.086688 * | 0.511525 * | 0.637688 * | 0.695 | 1.325 | 56.760 | |
| General model | −0.674451 * | 0.521555 * | 0.489129 * | 0.741 | 1.303 | 250.500 |
| Sistem | N ha−1 | V | W | C | CO2 | CO2 yr−1 | ||
|---|---|---|---|---|---|---|---|---|
| (m3 tree−1) | (m3 ha−1) | (kg tree−1) | (t ha−1) | (t C ha−1) | (t CO2 ha−1) | (t CO2 ha−1 yr−1) | ||
| Homogeneous | 278 | 0.0252 | 7.0163 | 50.820 | 14.128 | 7.064 | 25.925 | 2.592 |
| AFS 1 | 405 | 0.0254 | 10.2899 | 45.121 | 18.274 | 9.137 | 33.533 | 5.589 |
| AFS 2 | 209 | 0.0479 | 10.0087 | 107.739 | 22.517 | 11.259 | 41.320 | 5.903 |
| AFS 3 | 118 | 0.0163 | 1.9288 | 23.661 | 2.792 | 1.396 | 5.123 | 0.640 |
| AFS 4 | 313 | 0.0578 | 18.0926 | 154.792 | 48.450 | 24.225 | 88.906 | 9.878 |
| Silvopastoral | 100 | 0.1261 | 12.6057 | 267.135 | 26.714 | 13.357 | 49.019 | 4.456 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa Lopes, L.S.; Pauletto, D.; Gomes, E.S.C.; da Silva, Á.F.; de Sousa Oliveira, T.G.; da Silva, J.A.G.; Baloneque, D.D.; Martorano, L.G. Dendrometric Relationships and Biomass in Commercial Plantations of Dipteryx spp. in the Eastern Amazon. Forests 2023, 14, 2167. https://doi.org/10.3390/f14112167
de Sousa Lopes LS, Pauletto D, Gomes ESC, da Silva ÁF, de Sousa Oliveira TG, da Silva JAG, Baloneque DD, Martorano LG. Dendrometric Relationships and Biomass in Commercial Plantations of Dipteryx spp. in the Eastern Amazon. Forests. 2023; 14(11):2167. https://doi.org/10.3390/f14112167
Chicago/Turabian Stylede Sousa Lopes, Lucas Sérgio, Daniela Pauletto, Emeli Susane Costa Gomes, Ádria Fernandes da Silva, Thiago Gomes de Sousa Oliveira, Jéssica Aline Godinho da Silva, Diego Damázio Baloneque, and Lucieta Guerreiro Martorano. 2023. "Dendrometric Relationships and Biomass in Commercial Plantations of Dipteryx spp. in the Eastern Amazon" Forests 14, no. 11: 2167. https://doi.org/10.3390/f14112167
APA Stylede Sousa Lopes, L. S., Pauletto, D., Gomes, E. S. C., da Silva, Á. F., de Sousa Oliveira, T. G., da Silva, J. A. G., Baloneque, D. D., & Martorano, L. G. (2023). Dendrometric Relationships and Biomass in Commercial Plantations of Dipteryx spp. in the Eastern Amazon. Forests, 14(11), 2167. https://doi.org/10.3390/f14112167