Screening and Regulatory Mechanisms of Inter-Root Soil Nematicidal Bacteria of Pinus massoniana

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Treatment
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Analysis of Sequence Data
2.4. Screening and Identification of Bacteria for Controlling Pine Wood Nematode
2.4.1. Soil Samples
2.4.2. Pine Wood Nematode
2.4.3. Isolation of Soil Bacteria
2.4.4. Bacteria Screening for Nematocidal Activity
2.4.5. Re-screening of Nematocidal Bacteria
2.4.6. Evaluation of Nematocidal Activity in Fermentation Filtrates and Bacterial Suspensions of Four Bacterial Strains
2.5. Molecular Biology Identification
2.6. Inoculation of Pine Wood Nematode
2.7. Determination of Pine Peroxidase (POD) Activity
2.8. Determination of Pine Phenylalanine Ammonia-Lyase (PAL) Activity
2.9. Statistical Analysis
3. Results
3.1. Differential Analysis of Rhizosphere Microbial Diversity in Samples with Varying Disease Severity
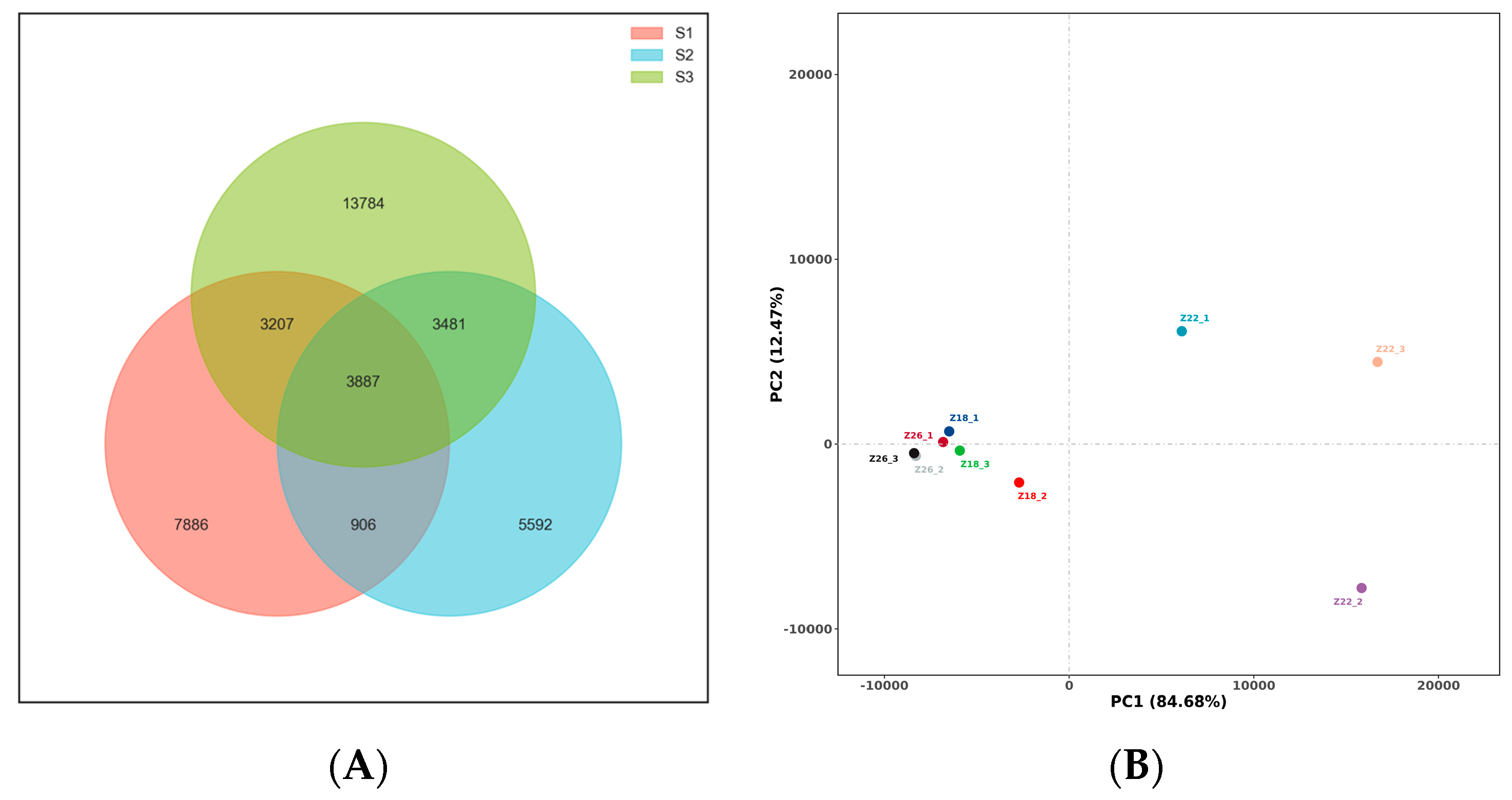
3.1.1. Three-Sample OTU Analysis and Correlation
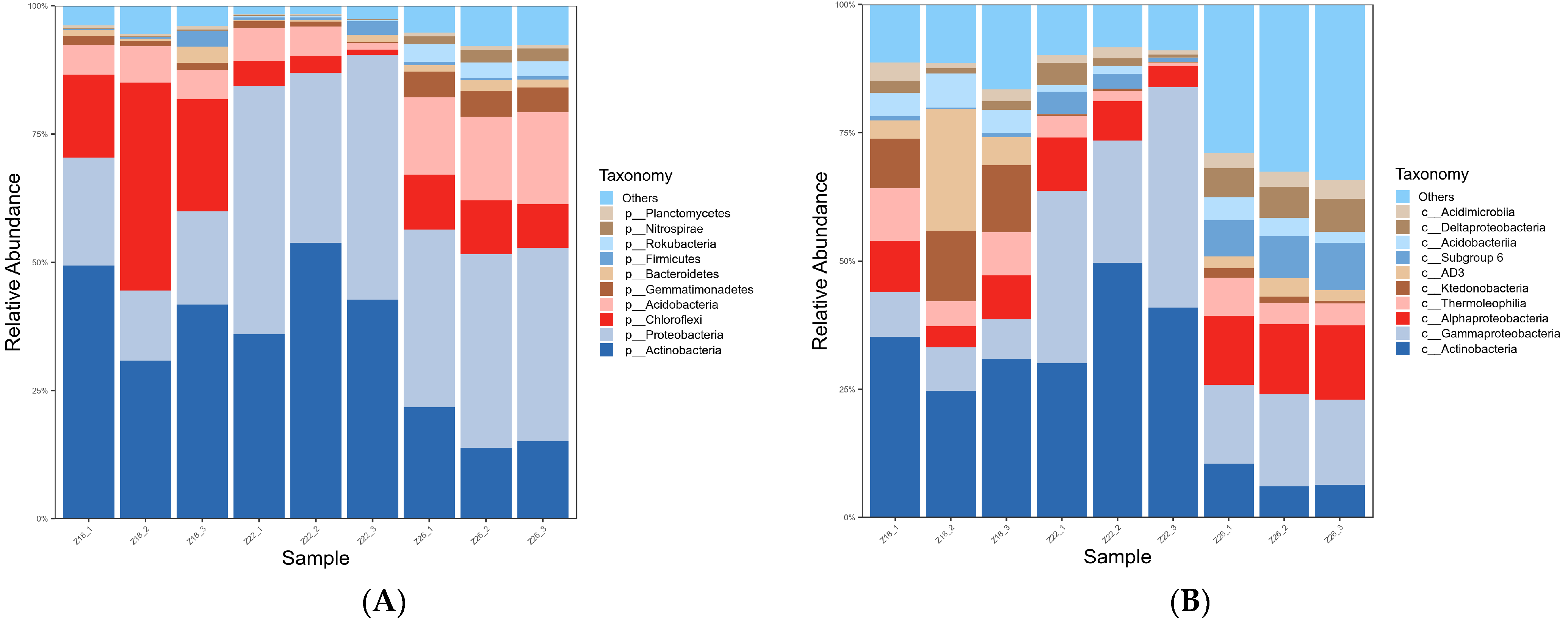
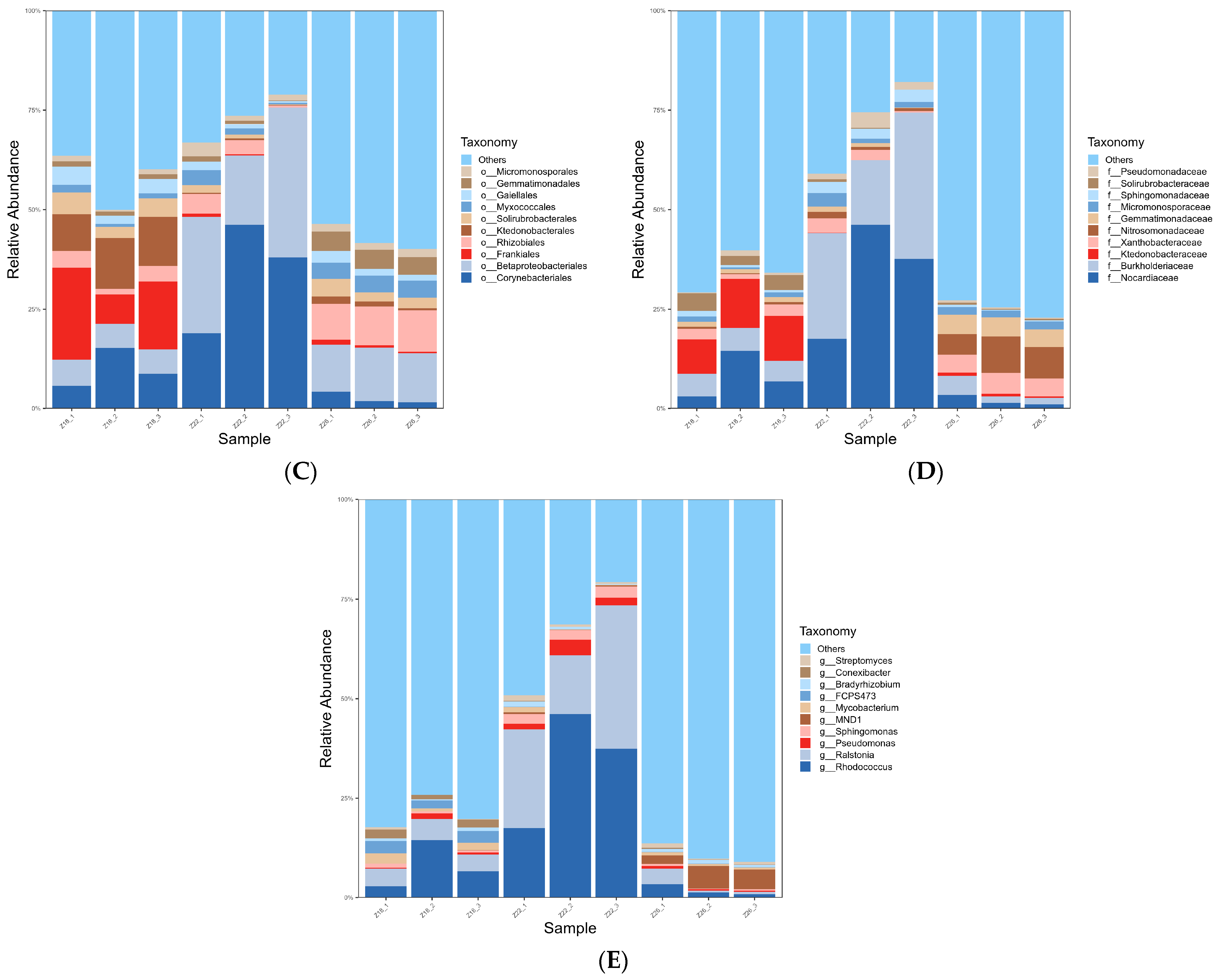
3.1.2. Analysis of Rhizosphere Microbial Species Composition in Three Sample Groups
3.2. Isolation and Screening of Threadkilling Bacteria
3.3. Determination of Nematocidal (Nematicidal) Activity of Fermentation Filtrate and Bacterial Suspension of 3 Bacterial Strains
3.4. Molecular Biological Identification of Three Bacterial Strains
3.5. Effects of Three Strains on PAL Content of Pinus Massoniana
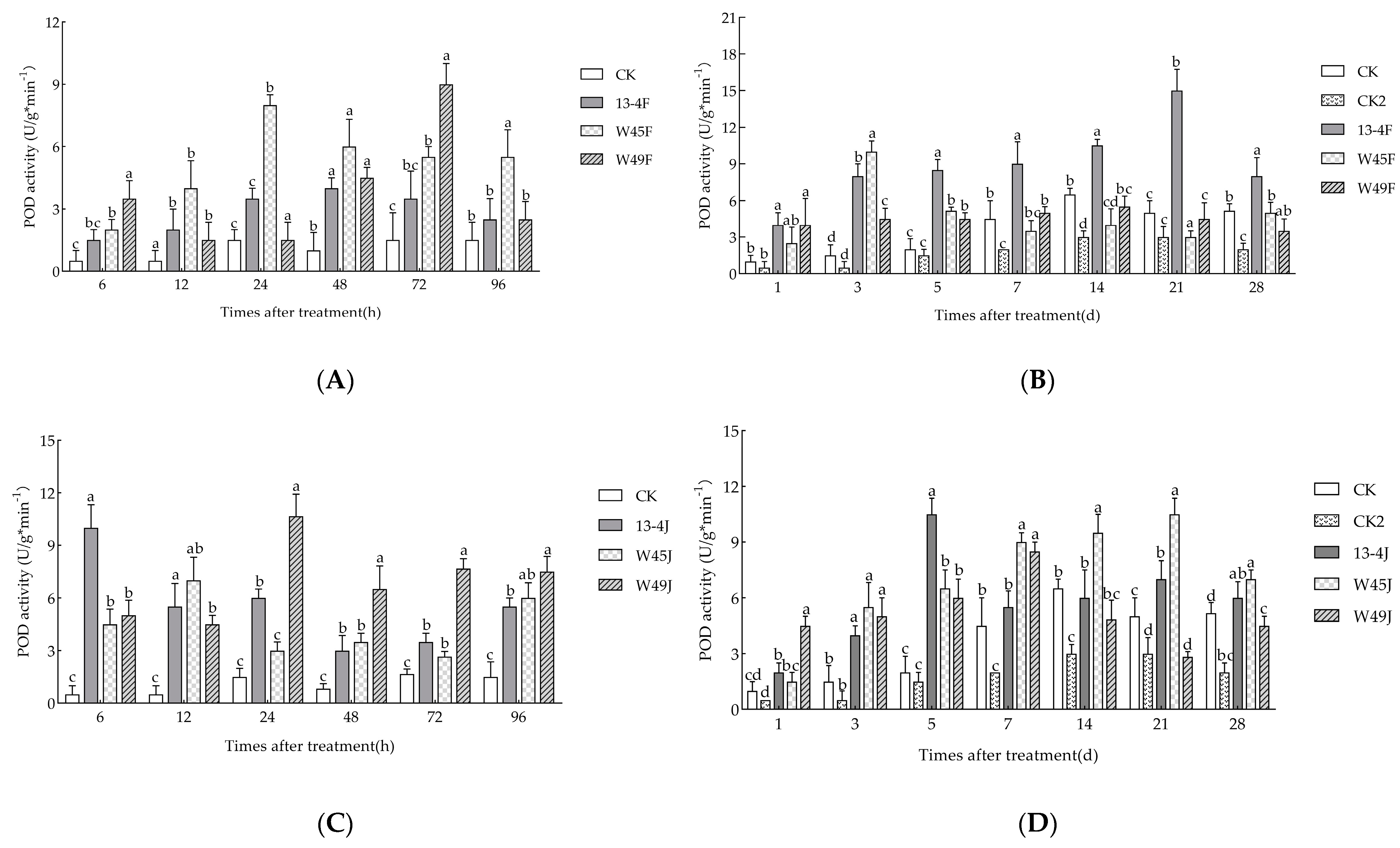
3.6. Effects of Three Strains on POD Content of Pinus massoniana
3.7. Screening and Control Effect of Pinus massoniana against Pine Wood Nematode Disease Induced by Linicide Strain
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nickle, W.R.; Golden, A.M.; Mamiya, Y.; Wergin, W. On the taxonomy and morphology of the pine wood nematode, Bursaphelenchus xylophilus (Steiner &Buhrer 1934) Nickle 1970. J. Nematol. 1981, 13, 385–392. [Google Scholar] [PubMed]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Zamora, P.; Rodríguez, V.; Renedo, F.; Sanz, A.V.; Domínguez, J.C.; Pérez-Escolar, G.; Miranda, J.; Alvarez, B.; Gonzalez-Casas, A.; Mayor, E.; et al. First report of Bursaphelenchus xylophilus causing pine wilt disease on Pinus radiata in Spain. Plant Dis. 2015, 99, 1449. [Google Scholar] [CrossRef]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.N.; Kim, J.H.; Ahn, J.Y.; Kim, S.; Cho, B.K.; Kim, Y.H.; Min, J. A short review of the pinewood nematode, Bursaphelenchus xylophilus. Toxicol. Environ. Health Sci. 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Chan, S.Y.; Liu, S.Y.; Seng, Z.; Chua, S.L. Biofilm matrix disrupts nematode motility and predatory behavior. ISME J. 2021, 15, 260–269. [Google Scholar] [CrossRef]
- Cheng, C.; Qin, J.; Wu, C.; Lei, M.; Wang, Y.; Zhang, L. Suppressing a plant-parasitic nematode with fungivorous behavior by fungal transformation of a Bt cry gene. Microb. Cell Factories 2018, 17, 116. [Google Scholar] [CrossRef]
- Jansen-Girgan, C.; Claassens, S.; Fourie, H. In vitro evaluations to determine the effect of Bacillus firmus strains on the motility of Meloidogyne incognita second-stage juveniles. Trop. Plant Pathol. 2016, 41, 320–324. [Google Scholar] [CrossRef]
- Paiva, G.; Proença, D.N.; Francisco, R.; Verissimo, P.; Santos, S.S.; Fonseca, L.; Morais, P.V. Nematicidal bacteria associated to pinewood nematode produce extracellular proteases. PLoS ONE 2013, 8, e79705. [Google Scholar] [CrossRef]
- Wang, Y.; Mei, L.; Wu, J.; Zhang, L. Detection and characterisation of a Bacillus thuringiensis crystal protein with nematicidal activity against the pinewood nematode Bursaphelenchus xylophilus. Biocontrol Sci. Technol. 2012, 22, 1143–1153. [Google Scholar] [CrossRef]
- Yu, J.; Du, G.; Li, R.; Li, L.; Li, Z.; Zhou, C.; Guo, D. Nematicidal activities of bacterial volatiles and components from two marine bacteria, Pseudoalteromonas marina strain H-42 and Vibrio atlanticus strain S-16, against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2015, 17, 1011–1025. [Google Scholar] [CrossRef]
- Wu, J.; Yin, Y.; Tan, J.; Hao, D. A preliminary study on resistance of Pinus massoniana induced by Bacillus cereus NJSZ-13 strain to Bursaphelenchus xylophilus. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 46, 4. [Google Scholar] [CrossRef]
- Han, G.; Mannaa, M.; Kim, N.; Jeon, H.W.; Jung, H.; Lee, H.H.; Kim, J.; Park, J.; Park, A.R.; Kim, J.C.; et al. Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease. Microorganisms 2021, 9, 688. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, L.; Zhou, J.; Yu, H.; Zhang, C.; Lv, Y.; Sun, J. Enhancement of oxidative stress contributes to increased pathogenicity of the invasive pine wood nematode. Philos. Trans. R. Soc. B 2019, 374, 20180323. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Liu, Z.; Zhang, X. Research advance of pathogenic mechanism of pine wood nematode. For. Pest. Dis. 2022, 41, 11–20. [Google Scholar]
- Cao, Y.; Wang, L.; Wang, X.; Fan, J. Pathogenicity of Bursaphelenchus xylophilus to Larix olgensis Seedlings. Sci. Silv. Sin. 2020, 56, 108–115. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Y.; Zhon, L.; Shen, D.; Zhu, L.; Fan, B. Inhibition of pine wood nematode Bursaphelenchus xylophilus by rhizobacterium Bacillus velezensis FZB42. Acta Microbiol. Sin. 2021, 61, 1287–1298. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, X.; Luan, Q.; Feng, J.; Wang, J.; Liu, M. Effects of Bursaphelenchus xylophilus Infestation on Physiological Indexes of Larix kaempferi. J. Southwest. For. Univ. 2023, 43, 6. [Google Scholar]
- Ren, W.; Ren, G.; Teng, Y.; Li, Z.; Li, L. Time-dependent effect of grapheneon the structure, abundance, and function of the soil bacterial community. J. Hazard. Mater. 2015, 297, 286–294. [Google Scholar] [CrossRef]
- Han, Z.M.; Hong, Y.D.; Zhao, B.G. A study on pathogenicity of bacteria carried by pine wood nematodes. J. Phytopathol. 2003, 151, 683–689. [Google Scholar] [CrossRef]
- Liu, M.J.; Hwang, B.S.; Jin, C.Z.; Li, W.J.; Park, D.J.; Seo, S.T.; Kim, C.J. Screening, isolation and evaluation of a nematicidal compound from actinomycetes against the pine wood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2018, 75, 1585–1593. [Google Scholar] [CrossRef]
- Elamary, R.; Salem, W. Optimizing and purifying extracellular amylase from soil bacteria to inhibit clinical biofilm-forming bacteria. PeerJ 2020, 8, e10288. [Google Scholar] [CrossRef]
- Maehara, N.; Futai, K. Effect of fungal interactions on the numbers of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), carried by the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Fundam. Appl. Nematol. 1997, 20, 611–617. [Google Scholar] [CrossRef]
- Xiong, Y.; Xia, F.; Huang, Z.; Liu, L.; Weng, X.; Wu, S. Isolation and Identification of Bacillus sp. Strain BRC-XYT1 and its Toxicity to Monochamus alternatus Hope. Curr. Biotechnol. 2016, 6, 5. [Google Scholar] [CrossRef]
- Vicente, C.S.; Ikuyo, Y.; Mota, M.; Hasegawa, K. Pinewood nematode-associated bacteria contribute to oxidative stress resistance of Bursaphelenchus xylophilus. BMC Microbiol. 2013, 13, 299. [Google Scholar] [CrossRef]
- Fukuda, K. Physiological process of the symptom development and resistance mechanism in pine wilt disease. J. For. Res. 1997, 2, 171–181. [Google Scholar] [CrossRef]
- Mabuhay, J.A.; Nakagoshi, N. Response of soil microbial communities to changes in a forest ecosystem brought about by pine wilt disease. Landsc. Ecol. Eng. 2012, 8, 189–196. [Google Scholar] [CrossRef]
- Liu, Y.; Ponpandian, L.N.; Kim, H.; Jeon, J.; Hwang, B.S.; Lee, S.K.; Park, S.-C.; Bae, H. Distribution and diversity of bacterial endophytes from four Pinus species and their efficacy as biocontrol agents for devastating pine wood nematodes. Sci. Rep. 2019, 9, 12461. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, K.; Yrjälä, K.; Liu, H.; Tong, Z.; Zhang, J. Introduction of broadleaf species into monospecific Cunninghamia lanceolata plantations changed the soil Acidobacteria subgroups composition and nitrogen-cycling gene abundances. Plant Soil 2021, 467, 29–46. [Google Scholar] [CrossRef]
- Pérez-García, A.; Romero, D.; De Vicente, A. Plant protection and growth stimulation by microorganisms: Biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Friman, V.P.; Gu, S.H.; Wang, X.F.; Eisenhauer, N.; Yang, T.J.; Ma, J.; Shen, Q.R.; Xu, Y.C.; et al. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. MBio 2016, 7, 10–1128. [Google Scholar] [CrossRef]
- Leclère, V.; Béchet, M.; Adam, A.; Guez, J.S.; Wathelet, B.; Ongena, M.; Thonat, P.; Gancel, F.; Cholletimbeert, M.; Jacques, P. Mycosubtilin overproduction by Bacillus subtilis BBG100 enhances the organism’s antagonistic and biocontrol activities. Appl. Environ. Microbiol. 2005, 71, 4577–4584. [Google Scholar] [CrossRef]
- Han, L.; Zhang, G.; Miao, G.; Zhang, X.; Feng, J. Streptomyces kanasensis sp. nov., an antiviral glycoprotein producing actinomycete isolated from forest soil around kanas lake of China. Curr. Microbiol. 2015, 71, 627–631. [Google Scholar] [CrossRef]
- Ballhausen, M.B.; Vandamme, P.; De Boer, W. Trait differentiation within the fungus-feeding (mycophagous) bacterial genus Collimonas. PLoS ONE 2016, 11, e0157552. [Google Scholar] [CrossRef]
- Oku, H.; Shiraishi, T.; Ouchi, S.; Kurozumi, S.; Ohta, H. Pine wilt toxin, the metabolite of a bacterium associated with a nematode. Naturwissenschaften 1980, 67, 198–199. [Google Scholar] [CrossRef]
- Le Dang, Q.; Son, S.W.; Cheon, H.M.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Kim, J.C. Pyochelin isolated from Burkholderia arboris KRICT1 carried by pine wood nematodes exhibits phytotoxicity in pine callus. Nematology 2011, 13, 521–528. [Google Scholar] [CrossRef]
- Zhao, B.G.; Wang, H.L.; Han, S.F.; Han, Z.M. Distribution and pathogenicity of bacteria species carried by Bursaphelenchus xylophilus in China. Nematology 2003, 5, 899–906. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Wang, C.; Wang, Y.; Sung, C. Esteya vermicola controls the pinewood nematode, Bursaphelenchus xylophilus, in pine seedlings. J. Nematol. 2017, 49, 86. [Google Scholar] [CrossRef]
- Maehara, N. Reduction of Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae) population by inoculating Trichoderma spp. into pine wilt-killed trees. Biol. Control 2008, 44, 61–66. [Google Scholar] [CrossRef]
- Jeong, M.H.; Yang, S.Y.; Lee, Y.S.; Ahn, Y.S.; Kim, K.Y. Selection and characterization of bacillus licheniformis mh48 for the biocontrol of pine wood nematode (Bursaphelenchus xylophilus). J. Korean For. Soc. 2015, 104, 512–518. [Google Scholar] [CrossRef]
- Yuan, Y.; Yan, Z.; Chen, Y.; Ye, J.; Tan, J. Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode. Diversity 2023, 15, 566. [Google Scholar] [CrossRef]
- Jähne, J.; Le Thi, T.T.; Blumenscheit, C.; Schneider, A.; Pham, T.L.; Le Thi, P.T.; Blom, J.; Vater, J.; Schweder, T.; Lasch, P.; et al. Novel plant-associated Brevibacillus and Lysinibacillus genomospecies harbor a rich biosynthetic potential of antimicrobial compounds. Microorganisms 2023, 11, 168. [Google Scholar] [CrossRef]
- Zhao, D.; Jiao, J.; Du, B.; Liu, K.; Wang, C.; Ding, Y. Volatile organic compounds from Lysinibacillus macroides regulating the seedling growth of Arabidopsis thaliana. Physiol. Mol. Biol. Plants 2022, 28, 1997–2009. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Anees, M.; Park, Y.S.; Kim, S.B.; Jung, W.J.; Kim, K.Y. Purification and properties of a Meloidogyne-antagonistic chitinase from Lysobacter capsici YS1215. Nematology 2014, 16, 63–72. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Kusajima, M.; Shima, S.; Fujita, M.; Minamisawa, K.; Che, F.-S.; Yamakawa, H.; Nakashita, H. Involvement of ethylene signalingin Azospirillum sp. B510-induced disease resistance in rice. Biosci. Biotechnol. Biochem. 2018, 82, 1522–1526. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Wang, J.; Jin, P.; Liu, H.; Zheng, Y. Bacillus cereus AR156-Induced Resistance to Colletotrichum acutatum Is Associated with Priming of Defense Responses in Loquat Fruit. PLoS ONE 2014, 9, e112494. [Google Scholar] [CrossRef]
- Khalifa, E.A.; Khalifa, R.K.M.; Haggag, H.E. Effect of Trichoderma species on damping off diseases incidence, some plant enzymes activity and nutritional status of bean plants. Am. J. Sci. 2010, 6, 10–57. [Google Scholar] [CrossRef]
- Jamal, Q.M.S.; Ahmad, V. Lysinibacilli: A biological factories intended for bio-insecticidal, bio-control, and bioremediation activities. J. Fungi 2022, 8, 1288. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | 24 h Mortality % | Adjusted Mortality % |
|---|---|---|
| 13-3 | 84.67 ± 0.73 e | 83.64 ± 0.78 e |
| W73 | 88.53 ± 1.54 cde | 87.77 ± 1.64 cde |
| W45 | 91.42 ± 0.76 abc | 90.85 ± 0.81 abc |
| 13-4 | 92.72 ± 0.93 ab | 92.23 ± 0.99 ab |
| W3 | 88.93 ± 1.19 bcd | 88.20 ± 1.27 bcd |
| W2 | 85.5 ± 1.04 de | 84.53 ± 1.11 de |
| W60 | 90.18 ± 1.18 abc | 89.52 ± 1.26 abc |
| W49 | 94.02 ± 0.87 a | 93.62 ± 0.93 a |
| W61 | 88.49 ± 1.17 cde | 87.73 ± 1.24 cde |
| W76 | 89.36 ± 1.14 bcd | 88.65 ± 1.21 bcd |
| W77 | 88.47 ± 1.02 cde | 87.70 ± 1.85 cde |
| W80 | 85.58 ± 0.70 de | 84.62 ± 1.09 de |
| W84 | 88.93 ± 0.50 bcd | 88.19 ± 0.75 bcd |
| Strain Number | Latin Name |
|---|---|
| 13-3 | Bacillus cereus |
| W73 | Bacillus siamensis |
| W45 | Bacillus paramycoides |
| 13-4 | Lysinibacillus capsici |
| W3 | Bacillus siamensis |
| W2 | Bacillus zanthoxyli |
| W60 | Lelliottia jeotgali |
| W49 | Delftia tsuruhatensis |
| W61 | Bacillus siamensis |
| W76 | Lysinibacillus |
| W77 | Pseudomonas geniculata |
| W80 | Bacillus siamensis |
| W84 | Bacillus siamensis |
| Treatment | Strain Number | Dilution Times | 24 h Mortality % | Adjusted Mortality % | 48 h Mortality % | Adjusted Mortality % |
|---|---|---|---|---|---|---|
| CK | 0 | 1.00 | 8.00 | |||
| Fermentation filtrate | 13-4 | 0 | 90 ± 0.64 b | 89.93 ± 1.11 b | 96.33 ± 0.75 ab | 95.74 ± 0.87 ab |
| W45 | 0 | 87.1 ± 0.67 b | 87.84 ± 1.17 b | 95.56 ± 0.83 ab | 94.84 ± 0.97 ab | |
| W49 | 0 | 94.74 ± 0.41 a | 94.63 ± 0.71 a | 98 ± 0.26 a | 97.68 ± 0.30 a | |
| Bacterial suspension | 13-4 | 0 | 81.98 ± 1.19 c | 83.22 ± 2.08 c | 98.33 ± 0.91 a | 98.068 ± 1.05 a |
| W45 | 0 | 71.51 ± 0.91 d | 70.36 ± 1.58 d | 94.33 ± 1.74 b | 93.42 ± 2.02 b | |
| W49 | 0 | 80.11 ± 1.40 c | 80.54 ± 2.45 c | 90.19 ± 0.81 c | 88.61 ± 0.94 c |
| Treatment | Strain Number | Dilution Times | 24 h Mortality % | Adjusted Mortality % | 48 h Mortality % | Adjusted Mortality % |
|---|---|---|---|---|---|---|
| CK | 0 | 1.00 | 8.00 | |||
| Fermentation filtrate | 13-4 | 2 | 81.04 ± 1.15 a | 79.77 ± 1.23 a | 95.26 ± 0.67 ab | 91.02 ± 0.77 ab |
| 4 | 75.84 ± 1.04 abc | 74.23 ± 1.11 abc | 88.5 ± 0.76 bbc | 86.65 ± 0.89 abc | ||
| W45 | 2 | 70 ± 1.47 d | 68 ± 1.57 d | 84.58 ± 1.85 cd | 82.1 ± 2.15 cd | |
| 4 | 42.78 ± 1.47 ef | 38.96 ± 1.57 ef | 81.64 ± 1.08 d | 78.68 ± 1.25 d | ||
| W49 | 2 | 80 ± 1.15 ab | 78.67 ± 1.23 ab | 94.01 ± 0.75 a | 93.04 ± 0.87 a | |
| 4 | 70.33 ± 1.57 d | 68.36 ± 1.68 d | 83.88 ± 1.25 d | 81.27 ± 1.46 d | ||
| Bacterial suspension | 13-4 | 2 | 75.73 ± 1.34 bc | 74.12 ± 1.43 bc | 84.86 ± 1.94 cd | 82.41 ± 2.26 cd |
| 4 | 35.94 ± 1.31 g | 31.67 ± 1.40 g | 67.04 ± 1.62 ef | 61.72 ± 1.88 ef | ||
| W45 | 2 | 72.43 ± 2.41 cd | 70.59 ± 2.57 cd | 80.54 ± 1.42 d | 77.4 ± 1.65 d | |
| 4 | 45.65 ± 1.55 e | 42.03 ± 1.65 e | 64.81 ± 1.44 f | 59.13 ± 1.68 f | ||
| W49 | 2 | 67.78 ± 1.49 d | 65.63 ± 1.59 d | 82.94 ± 0.93 d | 80.192 ± 1.08 d | |
| 4 | 40.01 ± 1.21 fg | 36.01 ± 1.29 fg | 70.83 ± 1.11 e | 66.12 ± 1.28 e |
| 13-4F | W45F | W49F | 13-4J | W45J | W49J | |
|---|---|---|---|---|---|---|
| POD | + | + | - | + | + | + |
| PAL | + | + | - | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Wang, Y.; Zhu, J.; Zhou, G.; Liu, J. Screening and Regulatory Mechanisms of Inter-Root Soil Nematicidal Bacteria of Pinus massoniana. Forests 2023, 14, 2230. https://doi.org/10.3390/f14112230
Liu M, Wang Y, Zhu J, Zhou G, Liu J. Screening and Regulatory Mechanisms of Inter-Root Soil Nematicidal Bacteria of Pinus massoniana. Forests. 2023; 14(11):2230. https://doi.org/10.3390/f14112230
Chicago/Turabian StyleLiu, Manman, Yating Wang, Jiacheng Zhu, Guoying Zhou, and Junang Liu. 2023. "Screening and Regulatory Mechanisms of Inter-Root Soil Nematicidal Bacteria of Pinus massoniana" Forests 14, no. 11: 2230. https://doi.org/10.3390/f14112230
APA StyleLiu, M., Wang, Y., Zhu, J., Zhou, G., & Liu, J. (2023). Screening and Regulatory Mechanisms of Inter-Root Soil Nematicidal Bacteria of Pinus massoniana. Forests, 14(11), 2230. https://doi.org/10.3390/f14112230