
Typical Plantation Water Use Strategies Are Determined by Environmental Conditions and Plant Eco-Physiology in Beijing, China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
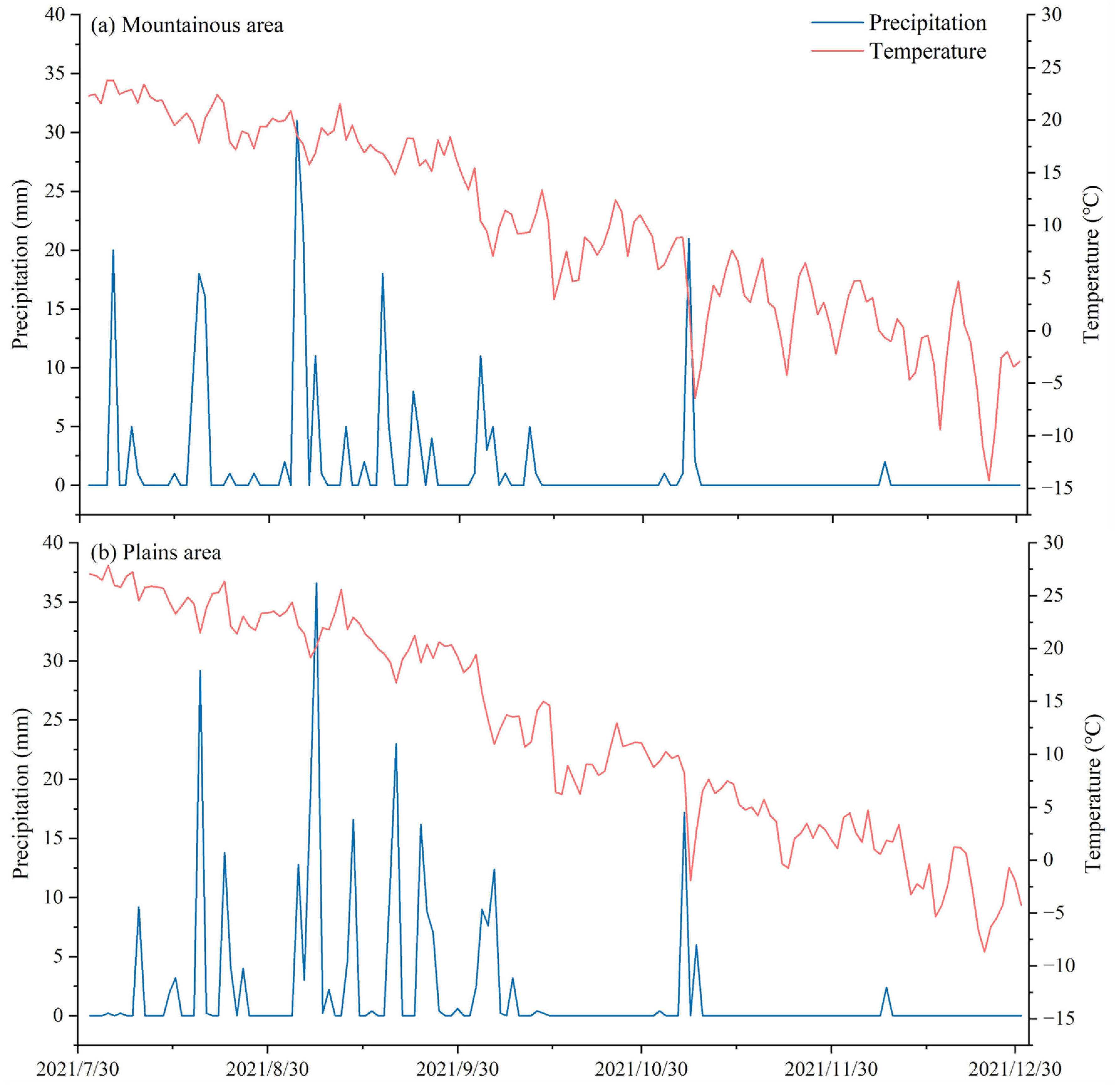
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Laboratory Analysis
2.4. Data Analysis
3. Results
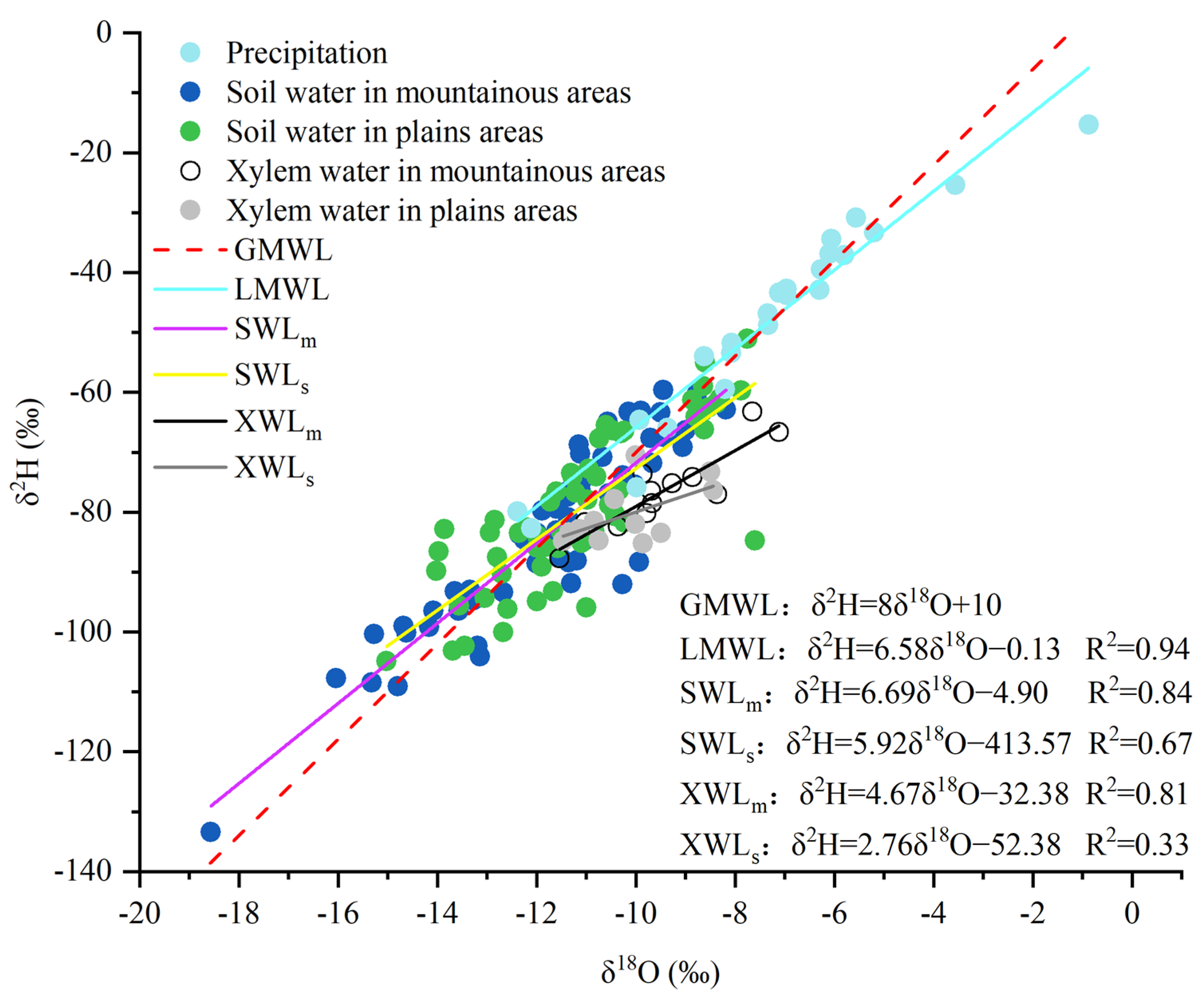
3.1. Isotopic Characteristics of Different Water Bodies
3.2. Variation in Soil Water Content in Different Areas
3.3. Isotopic Characteristics of Soil Water
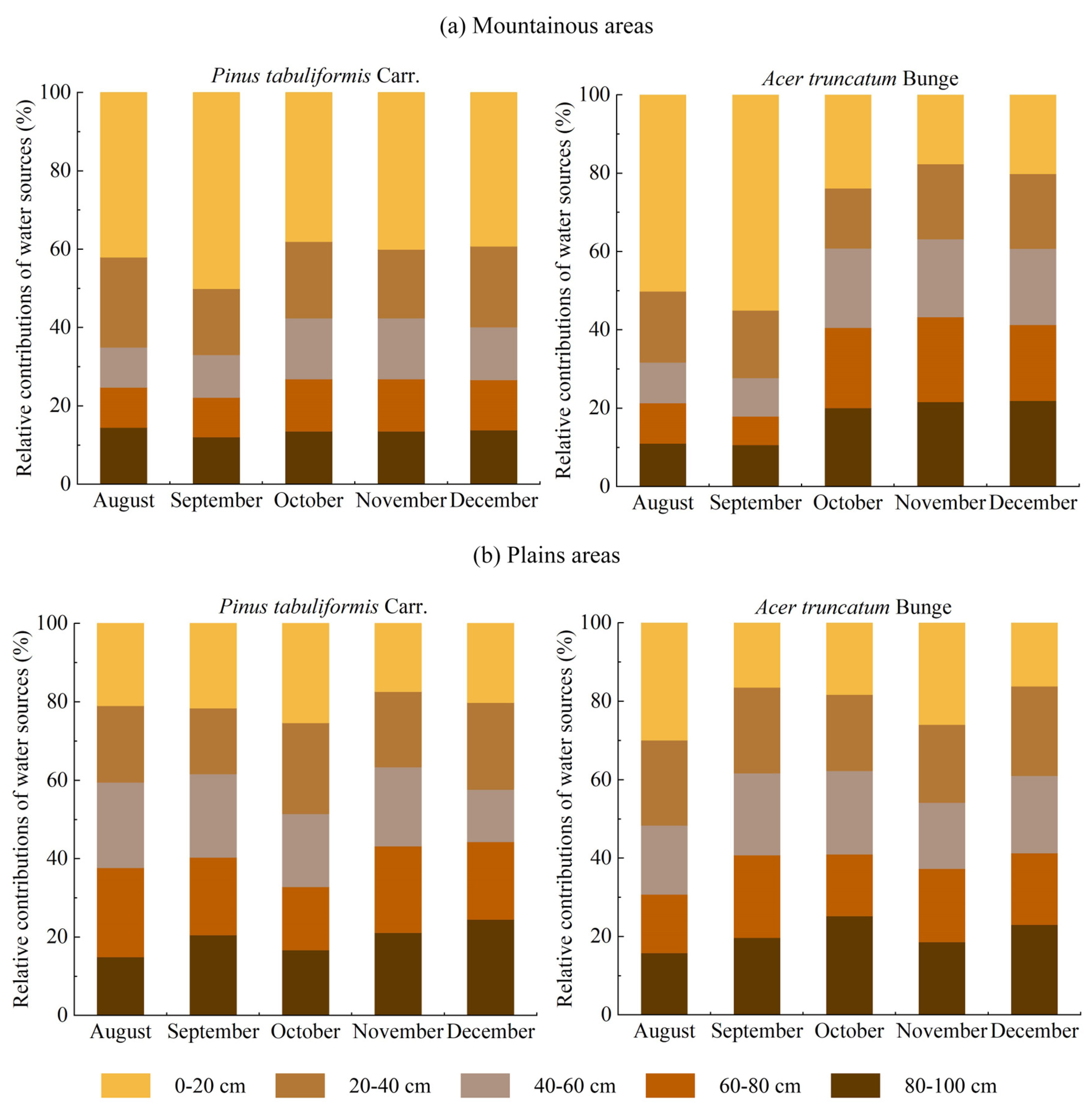
3.4. Water Sources for Typical Tree Species in Different Areas
4. Discussion
4.1. Soil Water Content Variation and Stable Isotope Distribution in Different Areas
4.2. Water Sources of Typical Tree Species in Different Areas
4.3. Implications for Forest Management in the Mountainous and Plains Areas of Beijing
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, K.Q.; Wang, B.T.; Wang, B.R.; Gao, H.P. Studies on the growth of forests with different density in the system of afforestation by water-harvesting. Sci. Silvae Sin. 2002, 38, 54–60. (In Chinese) [Google Scholar]
- Xu, Q.; Ji, C.L.; Wang, H.Y.; Li, Y. Use of stable isotopes of hydrogen, oxygen and carbon to identify water use strategy by plants. World For. Res. 2009, 22, 41–46. (In Chinese) [Google Scholar]
- Yu, Y.; Hua, T.; Chen, L.D.; Zhang, Z.Q.; Paulo, P. Divergent Changes in Vegetation Greenness, Productivity, and Rainfall Use Efficiency Are Characteristic of Ecological Restoration towards High-Quality Development in the Yellow River Basin, China. Engineering 2023. [Google Scholar] [CrossRef]
- Grant, G.E.; Dietrich, W.E. The frontier beneath our feet. Water Resour. Res. 2017, 53, 2605–2609. [Google Scholar] [CrossRef]
- Penna, D.; Hopp, L.; Scandellari, F.; Allen, S.T.; Benettin, P.; Beyer, M.; Geris, J.; Klaus, J.; Marshall, J.D.; Schwendenmann, L.; et al. Ideas and perspectives: Tracing terrestrial ecosystem water fluxes using hydrogen and oxygen stable isotopes—Challenges and opportunities from an interdisciplinary perspective. Biogeosciences 2018, 15, 6399–6415. [Google Scholar] [CrossRef]
- Rothfuss, Y.; Javaux, M. Reviews and syntheses: Isotopic approaches to quantify root water uptake: A review and comparison of methods. Biogeosciences 2017, 14, 2199–2224. [Google Scholar] [CrossRef]
- Duan, D.Y.; Ouyang, H. Application of stable hydrogen and oxygen isotope in analyzing plant water use sources. Ecol. Environ. 2007, 16, 22–30. (In Chinese) [Google Scholar]
- Grossiord, C.; Sevanto, S.; Dawson, T.E.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; Newman, B.D.; Stockton, E.A.; McDowell, N.G. Warming combined with more extreme precipitation regimes modifies the water sources used by trees. New Phytol. 2017, 213, 584–596. [Google Scholar] [CrossRef]
- Yang, B.; Wen, X.F.; Sun, X.M. Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agric. For. Meteorol. 2015, 201, 218–228. [Google Scholar] [CrossRef]
- Antunes, C.; Diaz-Barradas, M.C.; Zunzunegui, M.; Vieira, S.; Maguas, C. Water source partitioning among plant functional types in a semi-arid dune ecosystem. J. Veg. Sci. 2018, 29, 671–683. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Sun, H.Y.; Shen, Y.J.; Qi, Y.Q. Application of Hydrogen and Oxygen Stable Isotopes Technique in the Water Depletion of Ecosystems. Sci. Geogr. Sin. 2012, 32, 289–293. (In Chinese) [Google Scholar]
- Wei, F.L.; Li, S.C.; Yu, W.S.; Xie, A.L.; Hou, W.Y. A review of stable isotope in precipitation studies based on bibliometric and social network analysis. Acta Ecol. Sin. 2019, 39, 2634–2643. (In Chinese) [Google Scholar]
- Dawson, T.E.; Ehleringer, J.R. Streamside trees that do not use stream water. Nature 1992, 350, 335–337. [Google Scholar] [CrossRef]
- Zimmermann, U.; Munnich, K.O.; Roether, W.; Kreutz, W.; Schubach, K.; Siegel, O. Tracers determine movement of soil moisture and evapotranspiration. Science 1966, 152, 346–347. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Yu, X.X.; Jia, G.D.; Jia, J.B.; Lou, Y.H.; Lu, W.W. Contrasting water sources of evergreen and deciduous tree species in rocky mountain area of Beijing, China. Catena 2017, 150, 108–115. [Google Scholar] [CrossRef]
- Chen, Z.X.; Wang, G.H.; Pan, Y.H.; Yang, X.L.; Shen, Y.Y. Water use patterns differed notably with season and slope aspect for Caragana korshinskii on the Loess Plateau of China. Catena 2021, 198, 105028. [Google Scholar] [CrossRef]
- Ma, X.F.; Zhu, J.T.; Wang, Y.; Yan, W.; Zhao, C.Y. Variations in water use strategies of sand-binding vegetation along a precipitation gradient in sandy regions, northern China. J. Hydrol. 2021, 600, 126539. [Google Scholar] [CrossRef]
- Zhang, F.H.; Jia, W.X.; Zhu, G.F.; Zhang, Z.Y.; Shi, Y.; Yang, L.; Xiong, H.; Zhang, M.M. Using stable isotopes to investigate differences of plant water sources in subalpine habitats. Hydrol. Process. 2022, 36, e14518. [Google Scholar] [CrossRef]
- Beijing Municipal and Parks Bureau. Available online: https://yllhj.beijing.gov.cn/ztxx/lhysh/st/202302/t20230221_2920979.shtml (accessed on 25 September 2023).
- Christensen, C.W.; Hayashi, M.; Bentley, L.R. Hydrogeophysical survey of groundwater flow pathways in an alpine headwater basin. In Proceedings of the Near Surface Geoscience 2016—22nd European Meeting of Environmental and Engineering Geophysics, Barcelona, Spain, 4–8 September 2016. [Google Scholar]
- Xia, J.; Qiu, B.; Li, Y.Y. Water resources vulnerability and adaptive management in the Huang, Huai and Hai river basins of China. Water Int. 2012, 37, 523–536. [Google Scholar] [CrossRef]
- Wang, X.; Jia, G.D.; Deng, W.P.; Liu, Z.Q.; Liu, Z.H.; Qiu, G.F.; Li, W.L. Long-term water use characteristics and patterns of typical tree species in seasonal drought regions. Chin. J. Appl. Ecol. 2021, 32, 1943–1950. (In Chinese) [Google Scholar]
- West, A.G.; Patrickson, S.J.; Ehleringer, J.R. Water extraction times for plant and soil materials used in stable isotope analysis. Rapid Commun. Mass Spectrom. 2006, 20, 1317–1321. [Google Scholar] [CrossRef]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source Partitioning Using Stable Isotopes: Coping with Too Much Variation. PLoS ONE 2010, 5, e9672. [Google Scholar] [CrossRef] [PubMed]
- Stock, B.C.; Semmens, B.X. MixSIAR GUI User Manual, Version 3.1. 2013. Available online: https://github.com/brianstock/MixSIAR (accessed on 1 December 2022).
- Bowen, G.J.; Putman, A.; Brooks, J.R.; Bowling, D.R.; Oerter, E.J.; Good, S.P. Inferring the source of evaporated waters using stable H and O isotopes. Oecologia 2018, 187, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, E.B.; Bedrick, E.J. A Bayesian framework for stable isotope mixing models. Environ. Ecol. Stat. 2013, 20, 377–397. [Google Scholar] [CrossRef]
- Moore, J.W.; Semmens, B.X. Incorporating uncertainty and prior information into stable isotope mixing models. Ecol. Lett. 2008, 11, 470–480. [Google Scholar] [CrossRef]
- Parnell, A.C.; Phillips, D.L.; Bearhop, S.; Semmens, B.X.; Ward, E.J.; Moore, J.W.; Jackson, A.L.; Grey, J.; Kelly, D.J.; Inger, R. Bayesian stable isotope mixing models. Environmetrics 2013, 24, 387–399. [Google Scholar] [CrossRef]
- Craig, H. Isotopic Variations in Meteoric Waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef]
- Rossatto, D.R.; Silva, L.D.R.; Villalobos-Vega, R.; Sternberg, L.D.L.; Franco, A.C. Depth of water uptake in woody plants relates to groundwater level and vegetation structure along a topographic gradient in a neotropical savanna. Environ. Exp. Bot. 2012, 77, 259–266. [Google Scholar] [CrossRef]
- Wu, H.W.; Li, X.Y.; Jiang, Z.Y.; Li, J.; Zheng, X.R.; Zhao, D.Z. Variations in water use for Achnatherum splendens in Lake Qinghai watershed, based on δD and δ18O. Acta Ecol. Sin. 2015, 35, 8174–8183. (In Chinese) [Google Scholar]
- Qiu, X.; Zhang, M.; Wang, S. Preliminary research on hydrogen and oxygen stable isotope characteristics of different water bodies in the Qilian Mountains, northwestern Tibetan Plateau. Environ. Earth Sci. 2016, 75, 1491. [Google Scholar] [CrossRef]
- Che, C.W.; Zhang, M.J.; Argiriou, A.A.; Wang, S.J.; Du, Q.Q.; Zhao, P.P.; Ma, Z.Z. The Stable Isotopic Composition of Different Water Bodies at the Soil-Plant-Atmosphere Continuum (SPAC) of the Western Loess Plateau, China. Water 2019, 11, 1742. [Google Scholar] [CrossRef]
- Cheng, X.L.; An, S.Q.; Li, B.; Chen, J.Q.; Lin, G.H.; Liu, Y.H.; Luo, Y.Q.; Liu, S.R. Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China. Plant Ecol. 2006, 184, 1–12. [Google Scholar] [CrossRef]
- Gazis, C.; Feng, X. A stable isotope study of soil water: Evidence for mixing and preferential flow paths. Geoderma 2004, 119, 97–111. [Google Scholar] [CrossRef]
- Amin, A.; Zuecco, G.; Geris, J.; Schwendenmann, L.; McDonnell, J.J.; Borga, M.; Penna, D. Depth distribution of soil water sourced by plants at the global scale: A new direct inference approach. Ecohydrology 2020, 13, e2177. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Munoz-Villers, L.E.; Holwerda, F.; McDonnell, J.J.; Asbjornsen, H.; Dawson, T.E. Stable isotopes reveal linkages among ecohydrological processes in a seasonally dry tropical montane cloud forest. Ecohydrology 2012, 5, 779–790. [Google Scholar] [CrossRef]
- February, E.C.; Higgins, S.I. The distribution of tree and grass roots in savannas in relation to soil nitrogen and water. South Afr. J. Bot. 2010, 76, 517–523. [Google Scholar] [CrossRef]
- Deng, W.P.; Yu, X.X.; Jia, G.D.; Li, Y.J.; Liu, Y.J. Water Sources of Three Typical Plants in the Beijing Mountain Areas in Rainy Season. Arid. Zone Res. 2014, 31, 649–657. (In Chinese) [Google Scholar]
- Eggemeyer, K.D.; Awada, T.; Harvey, F.E.; Wedin, D.A.; Zhou, X.; Zanner, C.W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C-4 grasses in a semiarid grassland. Tree Physiol. 2009, 29, 157–169. [Google Scholar] [CrossRef]
- Chen, X.L.; Chen, Y.N.; Chen, Y.P. Relationship among water use of different plants in Heihe River riparian forests. Chin. J. Eco-Agric. 2014, 22, 972–979. (In Chinese) [Google Scholar]
- Mooney, H.A.; Gulmon, S.L.; Rundel, P.W.; Ehleringer, J. Further observations on the water relations of Prosopis tamarugo of the northern Atacama desert. Oecologia 1980, 44, 177–180. [Google Scholar] [CrossRef]
- Poot, P.; Lambers, H. Shallow-soil endemics: Adaptive advantages and constraints of a specialized root-system morphology. New Phytol. 2008, 178, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhao, W.Z.; Zheng, X.J.; Li, S.J. Root distribution of Nitraria sibirica with seasonally varying water sources in a desert habitat. J. Plant Res. 2015, 128, 613–622. [Google Scholar] [CrossRef]
- Dawson, T.E.; Pate, J.S. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: A stable isotope investigation. Oecologia 1996, 107, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R.; Dawson, T.E. Water uptake by plants: Perspectives from stable isotope composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Yu, X.X.; Jia, G.D.; Jia, J.B.; Lou, Y.H.; Zhang, K. Water Use Characteristics of Platycladus orientalis and Quercus variabilis in Beijing Mountain Area. Sci. Silvae Sin. 2016, 52, 22–30. (In Chinese) [Google Scholar]
- Liu, Z.Q.; Yu, X.X.; Jia, G.D.; Li, H.Z.; Lu, W.W. Water source of typical plants in rocky mountain area of Beijing, China. Chin. J. Appl. Ecol. 2017, 28, 2135–2142. (In Chinese) [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Zha, T.; Zhang, H.; Zhao, L.; Wang, A.; Cui, Q.; Song, J.; Zhang, H.; Yu, Y.; Wang, D. Typical Plantation Water Use Strategies Are Determined by Environmental Conditions and Plant Eco-Physiology in Beijing, China. Forests 2023, 14, 2277. https://doi.org/10.3390/f14122277
Chen Y, Zha T, Zhang H, Zhao L, Wang A, Cui Q, Song J, Zhang H, Yu Y, Wang D. Typical Plantation Water Use Strategies Are Determined by Environmental Conditions and Plant Eco-Physiology in Beijing, China. Forests. 2023; 14(12):2277. https://doi.org/10.3390/f14122277
Chicago/Turabian StyleChen, Yujia, Tonggang Zha, Hongli Zhang, Lili Zhao, Ao Wang, Qingzhou Cui, Junling Song, Hengshuo Zhang, Yang Yu, and Dongmei Wang. 2023. "Typical Plantation Water Use Strategies Are Determined by Environmental Conditions and Plant Eco-Physiology in Beijing, China" Forests 14, no. 12: 2277. https://doi.org/10.3390/f14122277