
Quantitative Assessment of Forest–Tundra Patch Dynamics in Polar Urals Due to Modern Climate Change



Abstract
:1. Introduction
2. Materials and Methods
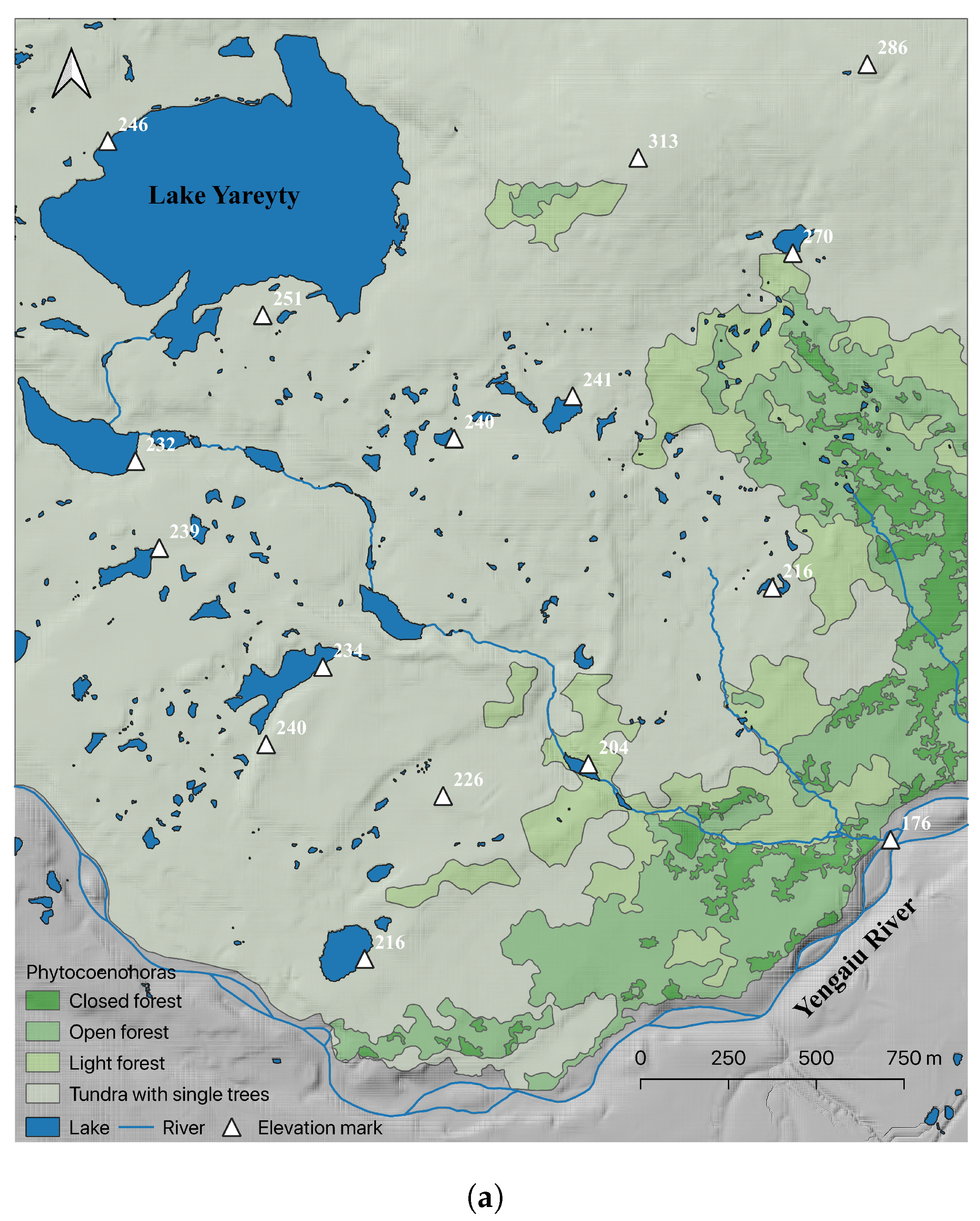
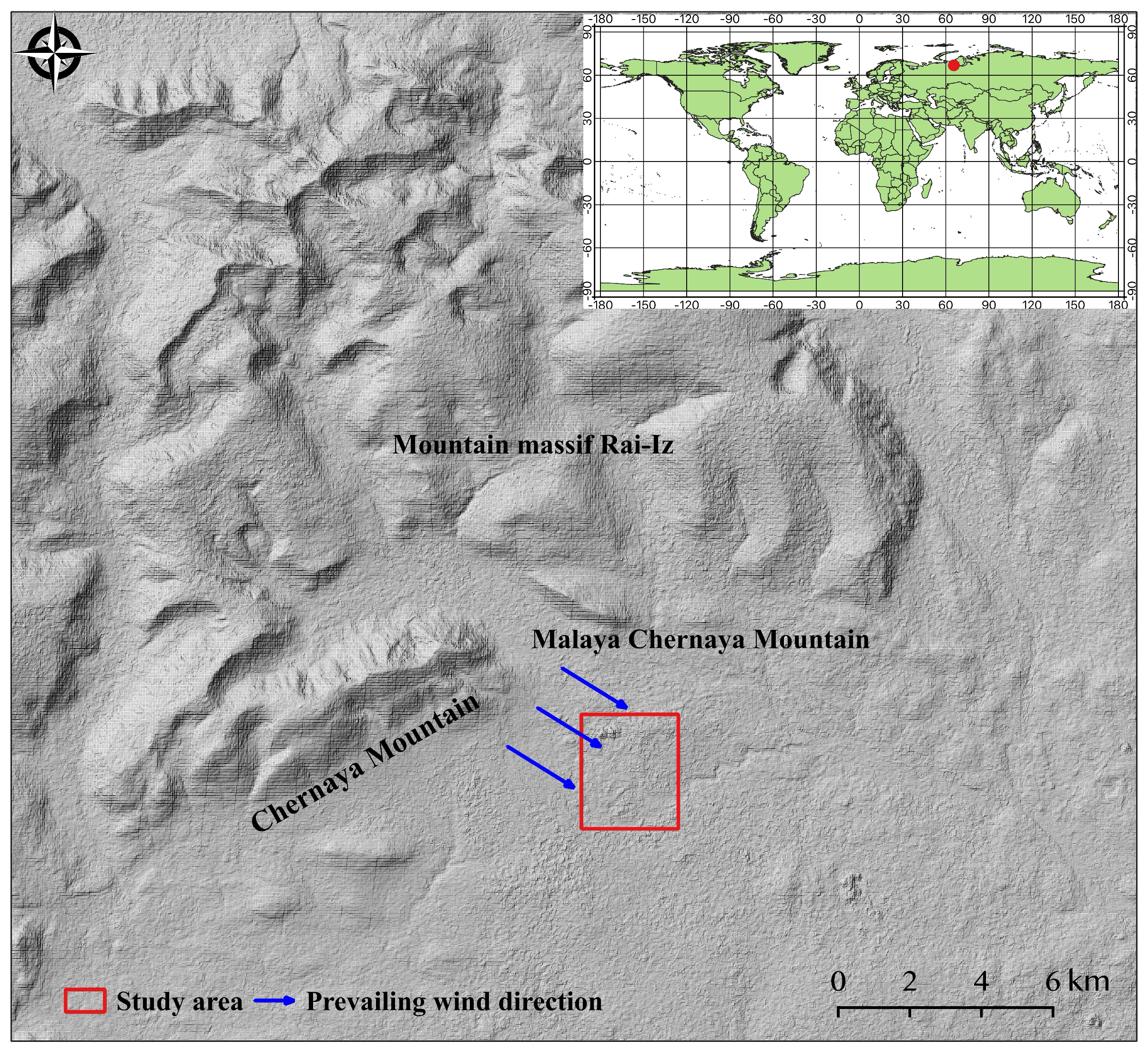
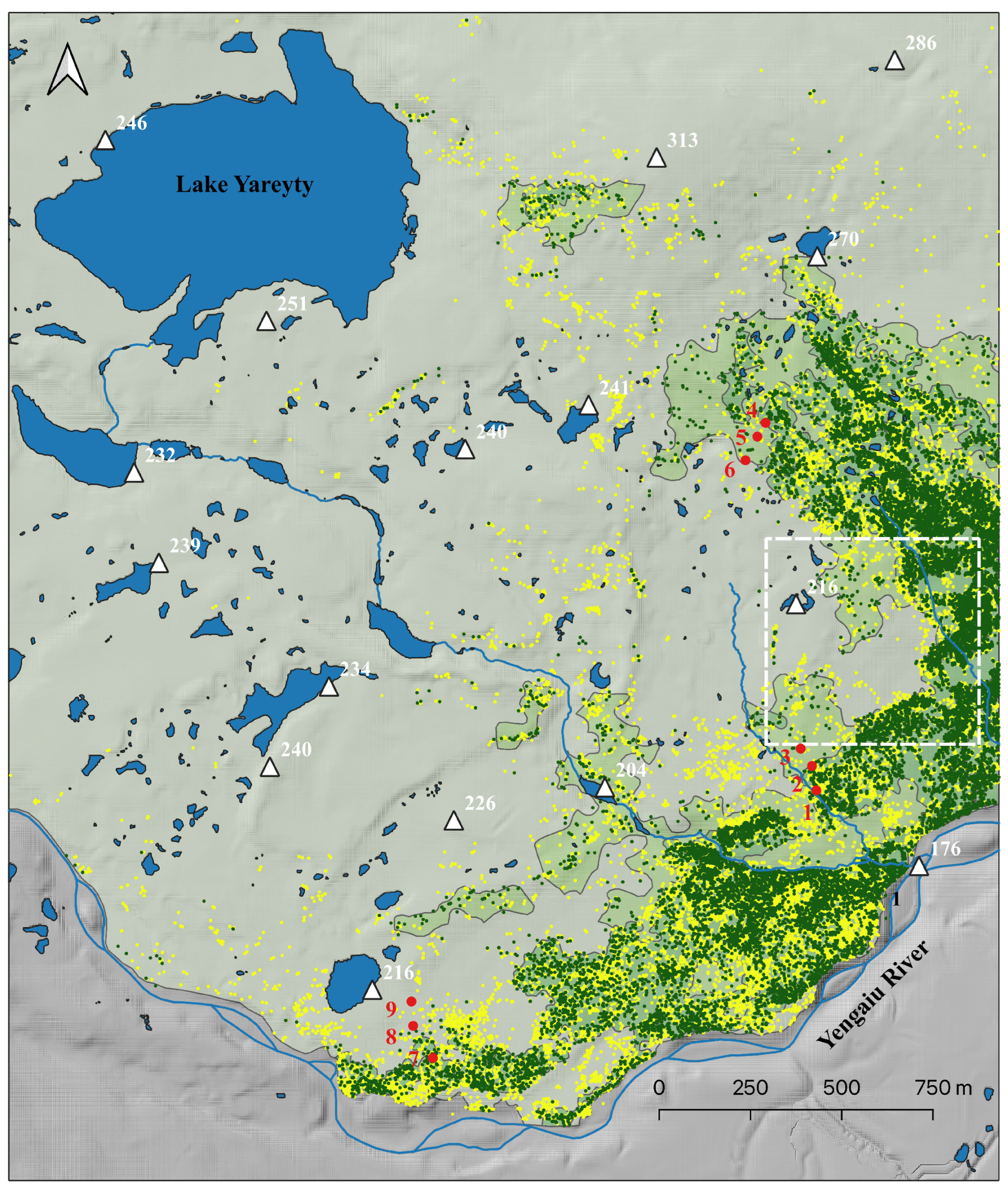
2.1. Study Area
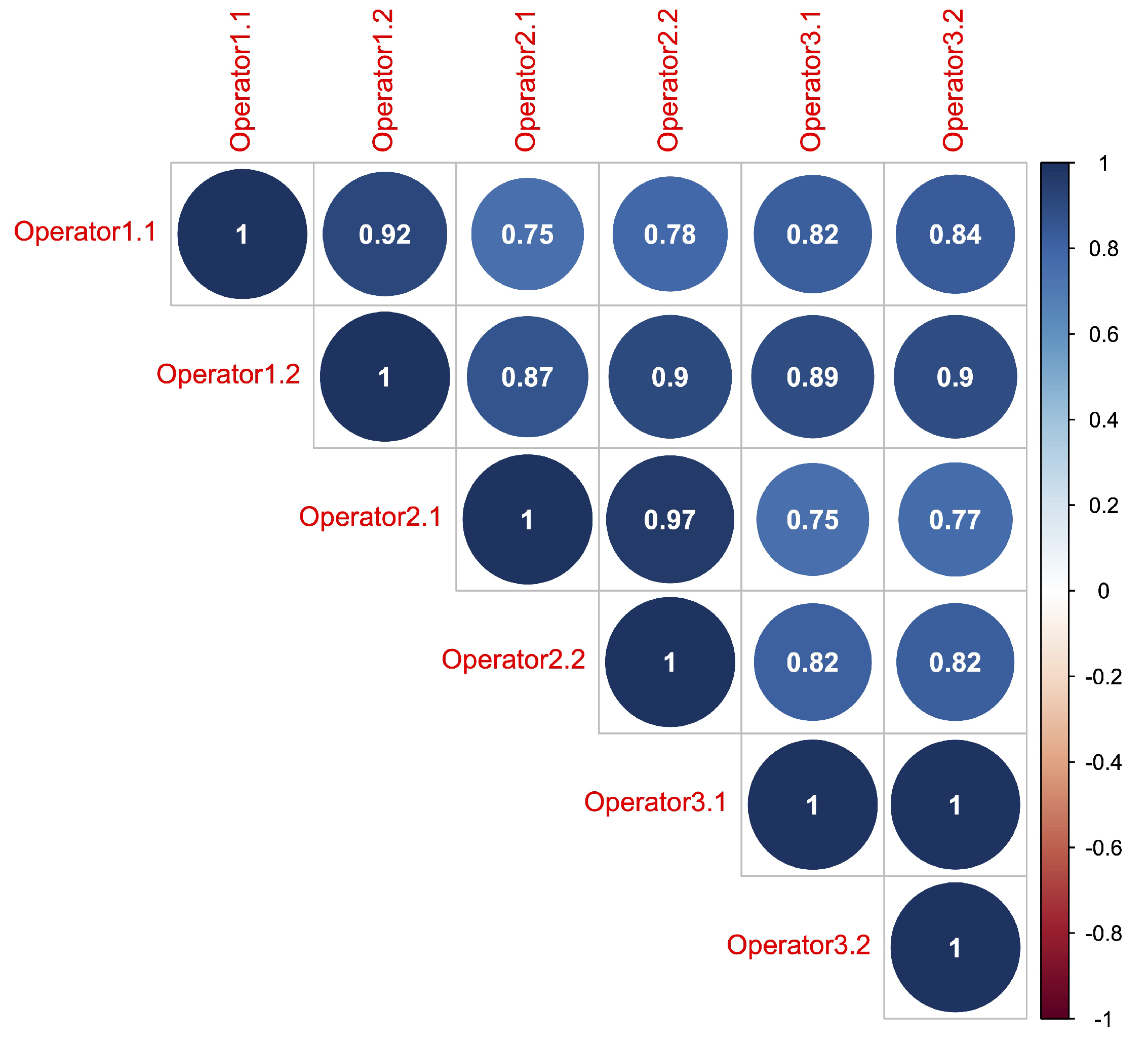
2.2. Ground Measurements
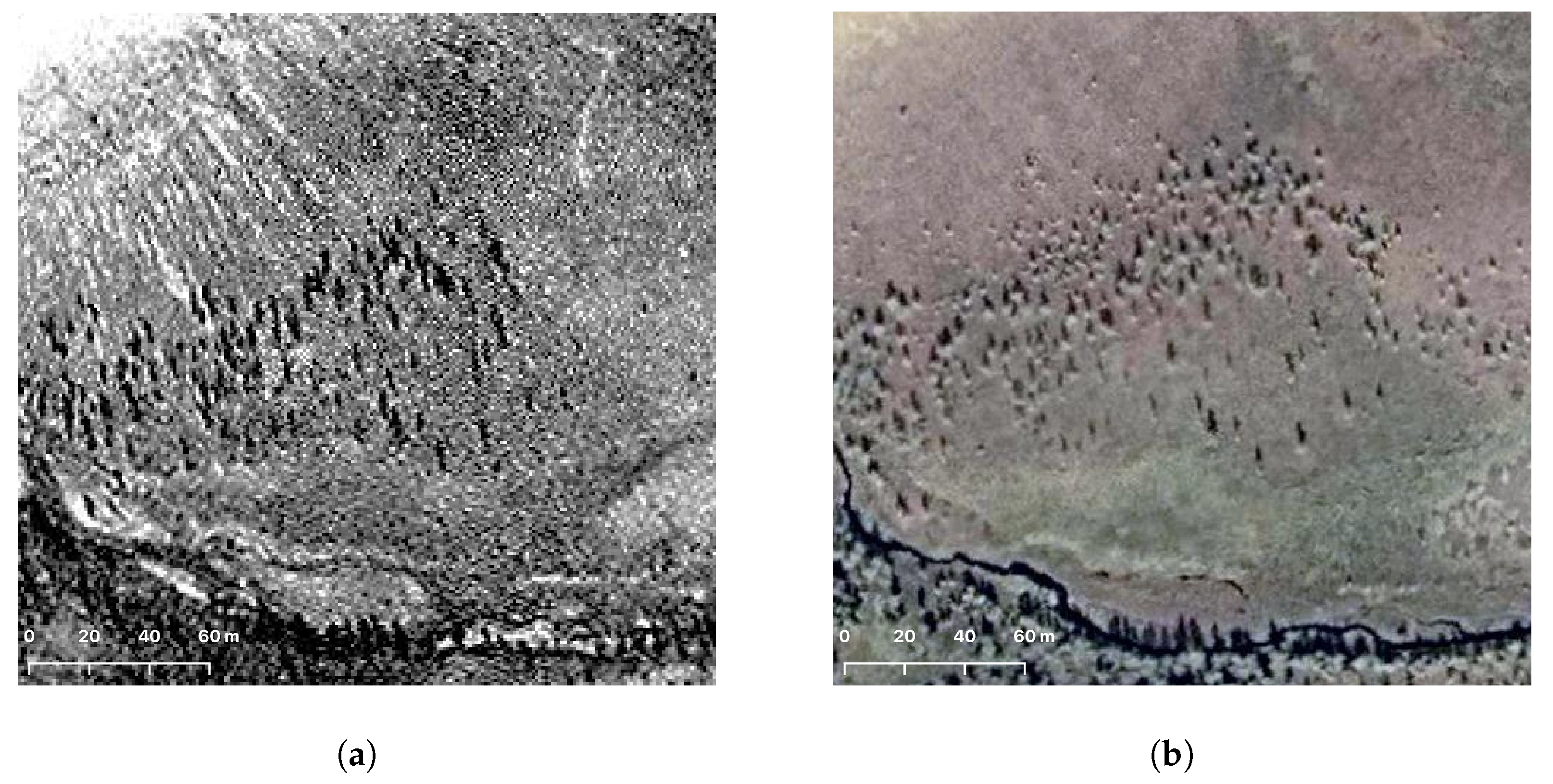
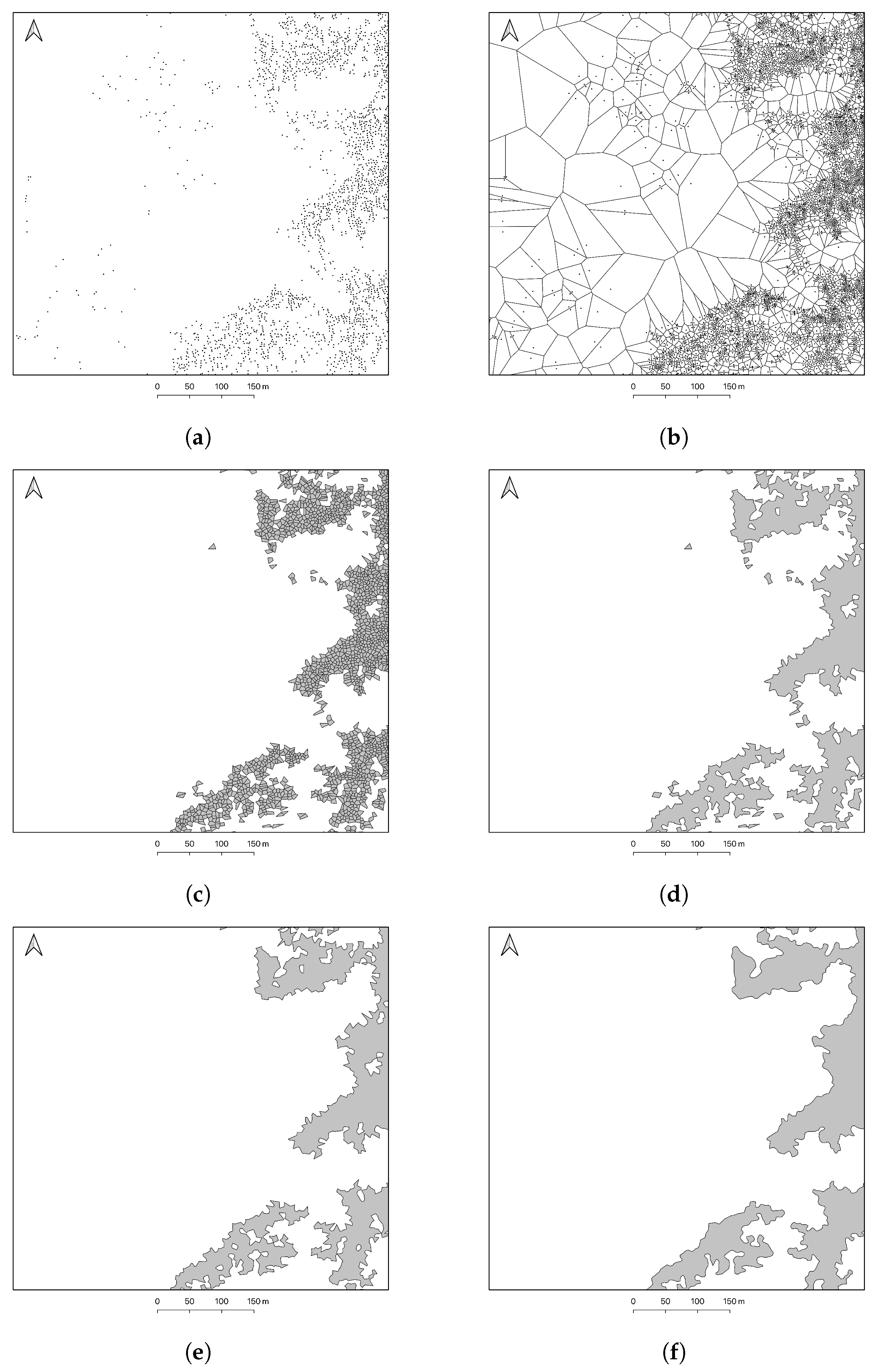
2.3. Tree Recognition
2.4. Forest–Tundra Patch Delineation

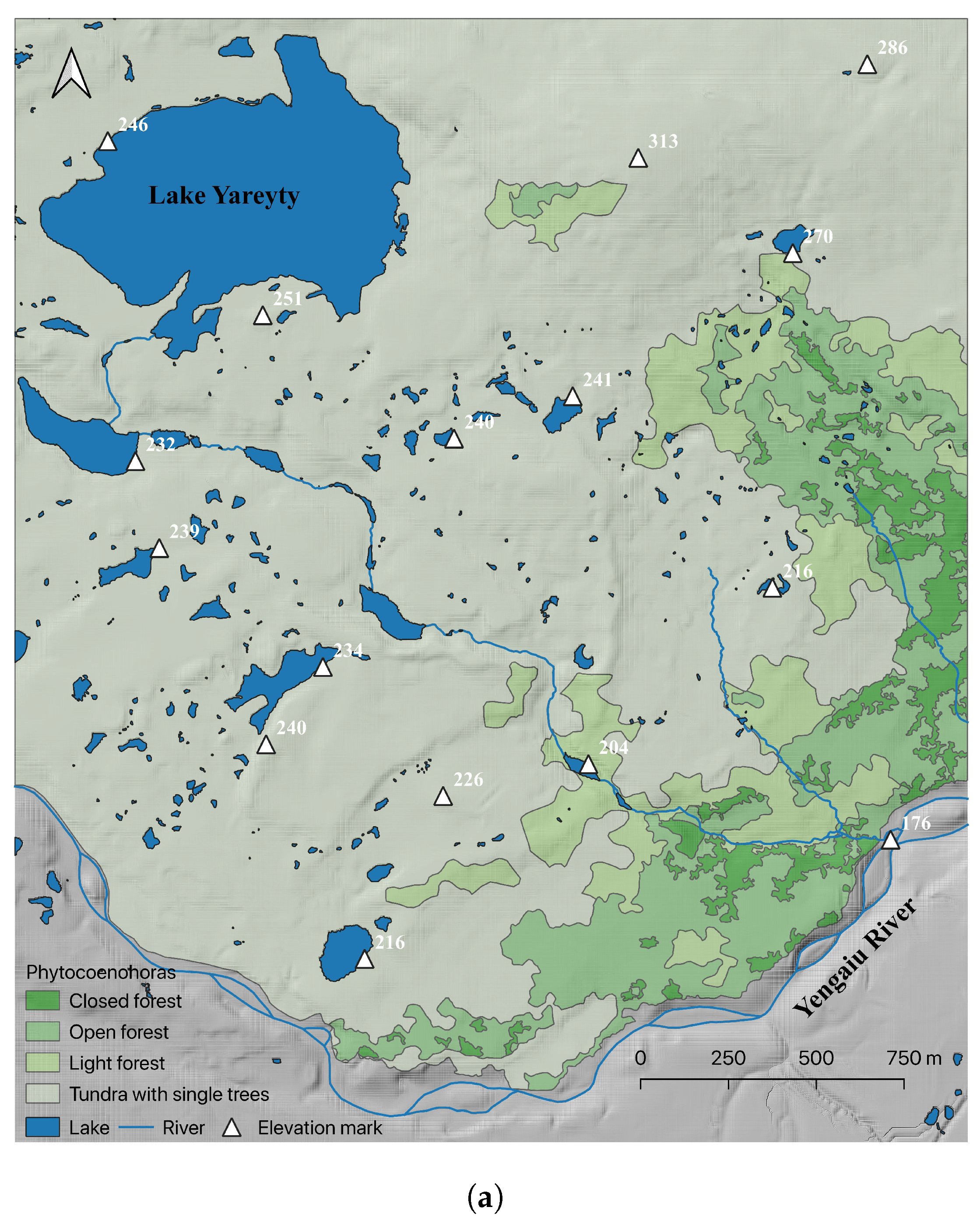
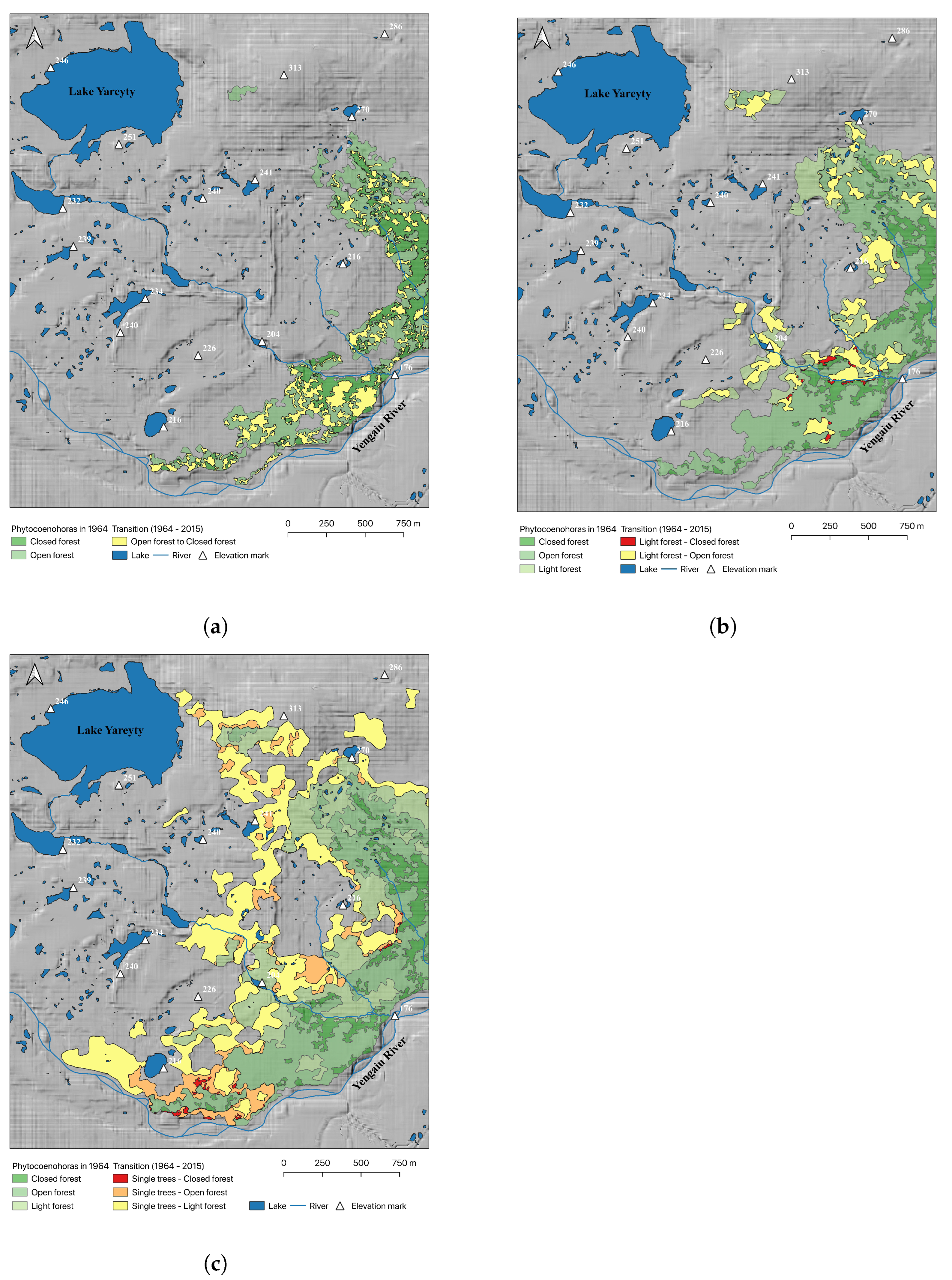
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shiyatov, S.G. Reconstruction of climate and the upper timberline dynamics since AD 745 by tree-ring data in the Polar Ural Mountains. In Proceedings of the International Conference on Past, Present and Future Climate, Helsinki, Finland, 22–25 August 1995; pp. 144–147. [Google Scholar]
- Kullman, L. Tree line population monitoring of Pinus sylvestris in the Swedish Scandes, 1973–2005: Implications for tree line theory and climate change ecology. J. Ecol. 2007, 95, 41–42. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartish, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’ev, M.; Bugman, H.; et al. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, L.; Agafonov, L.; Ljungqvist, F.C.; Churakova (Sidorova), O.; Düthorn, E.; Esper, J.; Hülsmann, L.; Kirdyanov, A.V.; Moiseev, P.; Myglan, V.S.; et al. Diverse growth trends and climate responses across Eurasia’s boreal forest. Environ. Res. Lett. 2016, 11, 074021. [Google Scholar] [CrossRef]
- Pellizzari, E.; Camarero, J.J.; Gazol, A.; Granda, E.; Shetti, R.; Wilmking, M.; Moiseev, P.; Pividori, M.; Carrer, M. Diverging shrub and tree growth from the Polar to the Mediterranean biomes across the European continent. Glob. Chang. Biol. 2017, 23, 3169–3180. [Google Scholar] [CrossRef] [PubMed]
- Mamet, S.D.; Brown, C.D.; Trant, A.J.; Laroque, C.P. Shifting global Larix distributions: Northern expansion and southern retraction as species respond to changing climate. J. Biogeogr. 2019, 46, 30–44. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Oldfather, M.F. Climate change, ecosystem processes and biological diversity responses in high elevation communities. Climate 2021, 9, 87. [Google Scholar] [CrossRef]
- Shiyatov, S.G. Rates of Change in the Upper Treeline Ecotone in the Polar Ural Mountains. Pages News 2003, 11, 8–10. [Google Scholar] [CrossRef]
- Shiyatov, S.G.; Terent’ev, M.M.; Fomin, V.V.; Zimmermann, N.E. Altitudinal and horizontal shifts of the upper boundaries of open and closed forests in the Polar Urals in the 20th century. Russ. J. Ecol. 2007, 38, 223–227. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Im, S.T.; Vdovin, A.S. Spatial distribution and temporal dynamics of high-elevation forest stands in southern Siberia. Glob. Ecol. Biogeogr. 2010, 19, 822–830. [Google Scholar] [CrossRef]
- Petrov, I.A.; Kharuk, V.I.; Dvinskaya, M.L.; Im, S.T. Reaction of coniferous trees in the Kuznetsk Alatau alpine forest-tundra ecotone to climate change. Contemp. Probl. Ecol. 2015, 8, 423–430. [Google Scholar] [CrossRef]
- Hansson, A.; Dargusch, P.; Shulmeister, J. A review of modern treeline migration, the factors controlling it and the implications for carbon storage. J. Mt. Sci. 2021, 18, 291–306. [Google Scholar] [CrossRef]
- Kark, S.; van Rensburg, B.J. Ecotones: Marginal or central areas of transition? Isr. J. Ecol. Evol. 2006, 52, 29–53. [Google Scholar] [CrossRef]
- Chiu, C.-A.; Lee, M.-F.; Tzeng, H.-Y.; Liao, M.-C. A concise scheme of vegetation boundary terms in subtropical high mountains. Afr. J. Agric. Res. J. Ecol. Evol. 2014, 9, 1560–1570. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. Global controls of forest line elevation in the northern and southern hemispheres. Glob. Ecol. Biogeogr. 2000, 9, 253–268. [Google Scholar] [CrossRef]
- Holtmeier, F.-K.; Broll, G. Treelines—Approaches at Different Scales. Sustainability 2017, 9, 808. [Google Scholar] [CrossRef]
- Smith, W.K.; Germino, M.J.; Hancock, T.E.; Johnson, D.M. Another perspective on altitudinal limits of alpine timberlines. Tree Physiol. 2003, 23, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef]
- Jain, A.K.; Murty, M.N.; Flynn, P.J. Data clustering: A review. ACM Comput. Surv. 1999, 31, 264–323. [Google Scholar] [CrossRef]
- Fortin, M.-J. Edge Detection Algorithms for Two-Dimensional Ecological Data. Ecology 1993, 75, 956–965. [Google Scholar] [CrossRef]
- Fortin, M.J.; Drapeau, P. Delineation of Ecological Boundaries: Comparison of Approaches and Significance Tests. Oikos 1995, 72, 323–332. [Google Scholar] [CrossRef]
- Gonzalez, R.C.; Woods, R.E. Digital Image Processing, 4th ed.; Pearson: New York, NY, USA, 2018; pp. 812–902. [Google Scholar]
- Clarke, C.L.; Edwards, M.E.; Gielly, L.; Ehrich, D.; Hughes, P.D.M.; Morozova, L.M.; Haflidason, H.; Mangerud, J.; Svendsen, J.I.; Alsos, I.G. Persistence of arctic-alpine flora during 24,000 years of environmental change in the Polar Urals. Sci. Rep. 2020, 9, 19613. [Google Scholar] [CrossRef] [PubMed]
- Shalaumova, Y.V.; Fomin, V.V.; Kapralov, D.S. Spatiotemporal dynamics of the Urals’ climate in the second half of the 20th century. Russ. Meteorol. Hydrol. 2010, 35, 107–114. [Google Scholar] [CrossRef]
- Gosz, J.R. Ecotone Hierarchies. Ecol. Appl. 1993, 3, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Fortin, M.-J.; Olson, R.J.; Ferson, S.; Iverson, L.; Hunsaker, C.; Edwards, G.; Klemas, V. Issues related to the detection of boundaries. Landsc. Ecol. 2000, 15, 453–466. [Google Scholar] [CrossRef]
- Hufkens, K.; Scheunders, P.; Ceulemans, R. Ecotones in vegetation ecology: Methodologies and definitions revisited. Ecol. Res. 2009, 24, 977–986. [Google Scholar] [CrossRef]
- Bateson, G. Mind and Nature: A Necessary Unity, 3rd ed.; E. P. Dutton: New York, NY, USA, 1979; pp. 16–229. [Google Scholar]
- Singh, C.P.; Panigrahy, S.; Thaplya, A.; Kimothi, M.M.; Soni, P.; Parihar, J.S. Monitoring the alpine treeline shift in parts of the Indian Himalayas using remote sensing. Curr. Sci. 2012, 102, 559–562. [Google Scholar]
- Kirdyanov, A.V.; Hagedorn, F.; Knorre, A.A.; Fedotova, E.V.; Vaganov, E.A.; Naurzbaev, M.M.; Moiseev, P.A.; Rigling, A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas 2012, 41, 56–67. [Google Scholar] [CrossRef]
- Aulló-Maestro, I.; Gómez, C.; Hernández, L.; Camarero, J.J.; Sánchez-González, M.; Cañellas, I.; Vázquez de la Cueva, A.; Montes, F. Monitoring montane-subalpine forest ecotone in the Pyrenees through sequential forest inventories and Landsat imagery. Ann. For. Sci. 2023, 80, 32. [Google Scholar] [CrossRef]
- Devi, N.; Hagedorn, F.; Moiseev, P.; Bugmann, H.; Shiyatov, S.; Mazepa, V.; Rigling, A. Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th century. Glob. Chang. Biol. 2008, 14, 1581–1591. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Treeline research-from the roots of the past to present time. A review. Forests 2020, 11, 38. [Google Scholar] [CrossRef]
- Cudlín, P.; Klopčič, M.; Tognetti, R.; Máliš, F.; Alados, C.L.; Bebi, P.; Grunewald, K.; Zhiyanski, M.; Andonowski, V.; Porta, N.L.; et al. Drivers of treeline shift in different European mountains. Clim. Res. 2017, 73, 135–150. [Google Scholar] [CrossRef]
- Mathisen, I.E.; Mikheeva, A.; Tutubalina, O.V.; Aune, A.; Hofgaard, A. Fifty years of tree line change in the Khibiny Mountains, Russia: Advantages of combined remote sensing and dendroecological approachesy. Appl. Veg. Sci. 2014, 17, 6–16. [Google Scholar] [CrossRef]
- Lackner, G.; Domine, F.; Nadeau, D.; Lafaysse, M.; Lackner, G.; Domine, F.; Nadeau, D.; Lafaysse, M.; Dumont, M.; Lackner, G.; et al. Snow properties at the forest—Tundra ecotone: Predominance of water vapor fluxes even in deep, moderately cold snowpacks. Cryospherei 2022, 16, 3357–3373. [Google Scholar] [CrossRef]
- Kharuk, I.V.; Im, S.T.; Dvinskaya, M.L.; Ranson, K.J. Climate-induced mountain tree-line evolution in southern Siberia. Scand. J. For. Res. 2010, 25, 446–454. [Google Scholar] [CrossRef]
- Fomin, V.V.; Shiyatov, S.G. Factors determining the phenomena in the upper tree line ecotone in the Polar Urals mountains. For. Russ. Econ. Them 2021, 16, 42–51. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Relocation of snow and its effects in the treeline ecotone—With special regard to the Rocky mountains, the Alps and Northern Europe. Cryospherei 2005, 136, 343–373. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area, ha/ % | |||
|---|---|---|---|
| Phytocoenohora | 1964 | 2015 | Δ (2015–1964) |
| Closed forest | 21.5/2.9 | 49.6/6.8 | 28.1/3.8 |
| Open forest | 70.5/9.6 | 96.2/13.1 | 25.7/3.5 |
| Light forest | 55.3/7.5 | 114.4/15.6 | 59.1/8.1 |
| Tundra with single trees | 585.7/79.9 | 472.8/64.5 | −112.9/−15.4 |
| Phytocoenohora Types | |||
|---|---|---|---|
| Num | 1964 | 2015 | Area, ha |
| 1 | Closed forest | Closed forest | 21.5 |
| 2 | Open forest | Closed forest | 25.8 |
| 3 | Open forest | Open forest | 44.7 |
| 4 | Light forest | Closed forest | 1.1 |
| 5 | Light forest | Open forest | 28.1 |
| 6 | Light forest | Light forest | 26.1 |
| 7 | Tundra with single trees | Closed forest | 1.2 |
| 8 | Tundra with single trees | Open forest | 23.4 |
| 9 | Tundra with single trees | Light forest | 88.3 |
| 10 | Tundra with single trees | Tundra with single trees | 472.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikhailovich, A.; Fomin, V. Quantitative Assessment of Forest–Tundra Patch Dynamics in Polar Urals Due to Modern Climate Change. Forests 2023, 14, 2340. https://doi.org/10.3390/f14122340
Mikhailovich A, Fomin V. Quantitative Assessment of Forest–Tundra Patch Dynamics in Polar Urals Due to Modern Climate Change. Forests. 2023; 14(12):2340. https://doi.org/10.3390/f14122340
Chicago/Turabian StyleMikhailovich, Anna, and Valery Fomin. 2023. "Quantitative Assessment of Forest–Tundra Patch Dynamics in Polar Urals Due to Modern Climate Change" Forests 14, no. 12: 2340. https://doi.org/10.3390/f14122340
APA StyleMikhailovich, A., & Fomin, V. (2023). Quantitative Assessment of Forest–Tundra Patch Dynamics in Polar Urals Due to Modern Climate Change. Forests, 14(12), 2340. https://doi.org/10.3390/f14122340