
Towards the Conservation of Monumental Taxus baccata L. Trees of Thasos Island: Genetic Insights


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Material
2.3. Molecular Analysis
2.4. Statistical Analysis
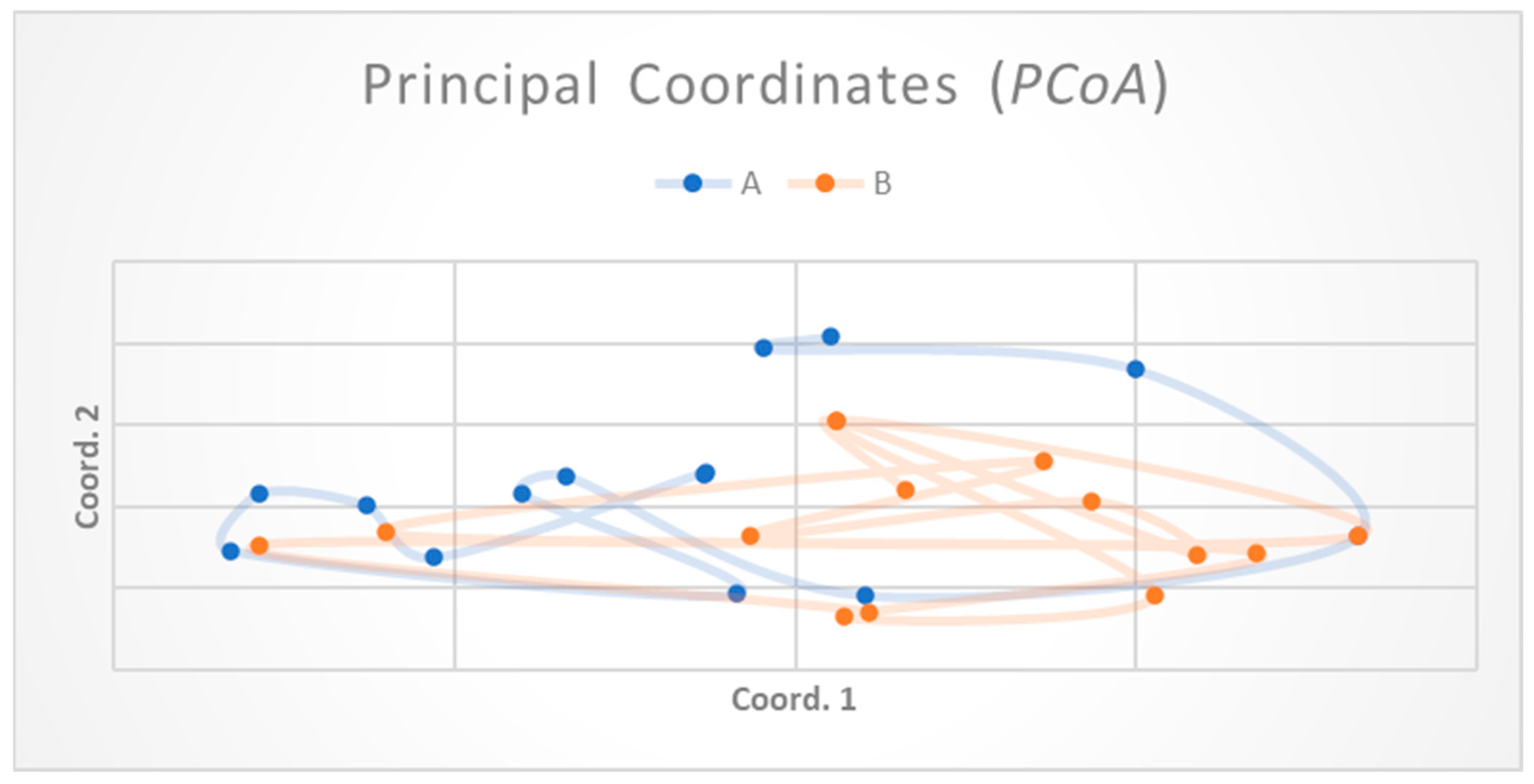
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. In Proceedings of the International Symposium on Population Genetics of Forest Trees, Corvallis, OR, USA, 31 July–2 August 1990; pp. 95–124. [Google Scholar]
- Robledo-Arnuncio, J.J. Wind pollination over mesoscale distances: An investigation with Scots pine. New Phytol. 2011, 190, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- García-Mozo, H.; López-Orozco, R.; Oteros, J.; Galán, C. Factors Driving Autumn Quercus Flowering in a Thermo-Mediterranean Area. Agronomy 2022, 12, 2596. [Google Scholar] [CrossRef]
- Thompson, J.N. The geographic mosaic of coevolution. In The Geographic Mosaic of Coevolution; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Sarvia, F.; De Petris, S.; Borgogno-Mondino, E. Exploring climate change effects on vegetation phenology by MOD13Q1 data: The piemonte region case study in the period 2001–2019. Agronomy 2021, 11, 555. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Peñuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef]
- Iwaizumi, M.G.; Mukasyaf, A.A.; Tamaki, I.; Nasu, J.Y.; Miyamoto, N.; Tamura, M.; Watanabe, A. Genetic diversity and structure of seed pools in an old planted Pinus thunbergii population and seed collection strategy for gene preservation. Tree Genet. Genomes 2023, 19, 9. [Google Scholar] [CrossRef]
- Young, A.; Boshier, D.; Boyle, T. Forest Conservation Genetics: Principles and Practice; CSIRO Publishing: Clayton, Australia, 2000. [Google Scholar]
- Oddou-Muratorio, S.; Petit, R.; Le Guerroué, B.; Guesnet, D.; Demesure, B. Pollen-versus seed-mediated gene flow in a scattered forest tree species. Evolution 2001, 55, 1123–1135. [Google Scholar]
- Thomas, P.; Polwart, A. Taxus baccata L. J. Ecol. 2003, 91, 489–524. [Google Scholar] [CrossRef]
- Dalmaris, E.; Avramidou, E.V.; Xanthopoulou, A.; Aravanopoulos, F.A. Dataset of targeted metabolite analysis for five taxanes of hellenic Taxus baccata l. populations. Data 2020, 5, 22. [Google Scholar] [CrossRef]
- Bnouham, M.; Ziyyat, A.; Mekhfi, H.; Tahri, A.; Legssyer, A. Medicinal plants with potential antidiabetic activity—A review of ten years of herbal medicine research (1990–2000). Dubai Diabetes Endocrinol. J. 2006, 14, 1–25. [Google Scholar] [CrossRef]
- Benkhnigue, O.; Ben Akka, F.; Salhi, S.; Fadli, M.; Douira, A.; Zidane, L. Catalogue des plantes médicinales utilisées dans le traitement du diabète dans la région d’Al Haouz-Rhamna (Maroc). J. Anim. Plant Sci. 2014, 23, 3539–3568. [Google Scholar]
- Bellakhdar, J. Contribution à L’étude de la Pharmacopée Traditionnelle au Maroc: La Situation Actuelle, les Produits, les Sources du Savoir (Enquête Ethnopharmacologique de Terrain Réalisée de 1969 à 1992). Ph.D. Thesis, Université Paul Verlaine-Metz, Metz, France, 1997. [Google Scholar]
- García, J.C. Biogeografía del tejo (Taxus baccata L.) en el Norte de África. El tejo en el Mediterráneo Occidental: Jornadas Internacionales sobre el tejo y las Tejeras en el Mediterráneo Occidental. Geography 2007, 177–183. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=3049781 (accessed on 3 December 2023).
- Iszkulo, G.; Pers-Kamczyc, E.; Nalepka, D.; Rabska, M.; Walas, L.; Dering, M. Postglacial migration dynamics helps to explain current scattered distribution of Taxus baccata. Dendrobiology 2016, 76, 81–89. [Google Scholar] [CrossRef]
- IUCN Species Survival Commission. IUCN Red List Categories and Criteria: Version 3.1; IUCN Species Survival Commissio: Gland, Switzerland, 2001. [Google Scholar]
- Myking, T.; Vakkari, P.; Skrøppa, T. Genetic variation in northern marginal Taxus baccata L. populations. Implications for conservation. Forestry 2009, 82, 529–539. [Google Scholar] [CrossRef]
- Schirone, B.; Ferreira, R.C.; Vessella, F.; Schirone, A.; Piredda, R.; Simeone, M.C. Taxus baccata in the Azores: A relict form at risk of imminent extinction. Biodivers. Conserv. 2010, 19, 1547–1565. [Google Scholar] [CrossRef]
- Devaney, J.L.; Jansen, M.A.; Whelan, P.M. Spatial patterns of natural regeneration in stands of English yew (Taxus baccata L.); Negative neighbourhood effects. For. Ecol. Manag. 2014, 321, 52–60. [Google Scholar] [CrossRef]
- Iszkuło, G.; Jasińska, A.K.; Giertych, M.J.; Boratyński, A. Do secondary sexual dimorphism and female intolerance to drought influence the sex ratio and extinction risk of Taxus baccata? Plant Ecol. 2009, 200, 229–240. [Google Scholar] [CrossRef]
- Dubreuil, M.; Riba, M.; González-Martínez, S.C.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Genetic effects of chronic habitat fragmentation revisited: Strong genetic structure in a temperate tree, Taxus baccata (Taxaceae), with great dispersal capability. Am. J. Bot. 2010, 97, 303–310. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Dubreuil, M.; Riba, M.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Spatial genetic structure of Taxus baccata L. in the western Mediterranean Basin: Past and present limits to gene movement over a broad geographic scale. Mol. Phylogenet. Evol. 2010, 55, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Mayol, M.; Riba, M.; González-Martínez, S.C.; Bagnoli, F.; de Beaulieu, J.L.; Berganzo, E.; Burgarella, C.; Dubreuil, M.; Krajmerová, D.; Paule, L. Adapting through glacial cycles: Insights from a long-lived tree (Taxus baccata). New Phytol. 2015, 208, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Burgarella, C.; Navascués, M.; Zabal-Aguirre, M.; Berganzo, E.; Riba, M.; Mayol, M.; Vendramin, G.; Gonzalez-Martinez, S.C. Recent population decline and selection shape diversity of taxol-related genes. Mol. Ecol. 2012, 21, 3006–3021. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Herbivory, plant regeneration, and species coexistence. J. Ecol. 1996, 84, 609–615. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Magård, E. Population ecology and conservation status of the last natural population of English yew Taxus baccata in Denmark. Biol. Conserv. 1999, 88, 173–182. [Google Scholar] [CrossRef]
- Linares, J.C. Shifting limiting factors for population dynamics and conservation status of the endangered English yew (Taxus baccata L., Taxaceae). For. Ecol. Manag. 2013, 291, 119–127. [Google Scholar] [CrossRef]
- Heilbuth, J.C. Lower species richness in dioecious clades. Am. Nat. 2000, 156, 221–241. [Google Scholar] [CrossRef]
- Vamosi, J.C.; Vamosi, S.M. Present day risk of extinction may exacerbate the lower species richness of dioecious clades. Divers. Distrib. 2005, 11, 25–32. [Google Scholar] [CrossRef]
- Zhang, D.; Yu, H.; Gao, L.; Wang, J.; Dong, H.; Guo, Y.; Hu, S. Genetic Diversity in Oilseed and Vegetable Mustard (Brassica juncea L.) Accessions Revealed by Nuclear and Mitochondrial Molecular Markers. Agronomy 2023, 13, 919. [Google Scholar] [CrossRef]
- Fields, J.; Saxton, A.M.; Beyl, C.A.; Kopsell, D.A.; Cregan, P.B.; Hyten, D.L.; Cuvaca, I.; Pantalone, V.R. Seed Protein and Oil QTL in a Prominent Glycine max Genetic Pedigree: Enhancing Stability for Marker Assisted Selection. Agronomy 2023, 13, 567. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Kumar, A.; Sharma, S.; Singh, B.; Sood, S.; Dipta, B.; Singh, R.; Siddappa, S.; Thakur, A.K.; Dalamu, D. Analysis of Genetic Diversity, Population Structure and Association Mapping for Late Blight Resistance in Potato (Solanum tuberosum L.) Accessions Using SSR Markers. Agronomy 2023, 13, 294. [Google Scholar] [CrossRef]
- Zafeiriou, I.; Sakellariou, M.; Mylona, P.V. Seed Phenotyping and Genetic Diversity Assessment of Cowpea (V. unguiculata) Germplasm Collection. Agronomy 2023, 13, 274. [Google Scholar] [CrossRef]
- Hu, G.; Jiang, Q.; Wang, Z.; Li, Z.; Liao, W.; Shen, D.; Zhong, C. Genetic Diversity Analysis and Core Collection Construction of the Actinidia chinensis Complex (Kiwifruit) Based on SSR Markers. Agronomy 2022, 12, 3078. [Google Scholar] [CrossRef]
- Ganopoulos, I.V.; Avramidou, E.; Fasoula, D.A.; Diamantidis, G.; Aravanopoulos, F.A. Assessing inter-and intra-cultivar variation in Greek Prunus avium by SSR markers. Plant Genet. Resour. 2010, 8, 242–248. [Google Scholar] [CrossRef]
- De Dato, G.D.; Teani, A.; Mattioni, C.; Aravanopoulos, F.; Avramidou, E.V.; Stojnic, S.; Ganopoulos, I.; Belletti, P.; Ducci, F. Genetic analysis by nuSSR markers of Silver Birch (Betula pendula Roth) populations in their Southern European distribution range. Front. Plant Sci. 2020, 11, 310. [Google Scholar] [CrossRef]
- Avramidou, E.; Ganopoulos, I.V.; Aravanopoulos, F.A. DNA fingerprinting of elite Greek wild cherry (Prunus avium L.) genotypes using microsatellite markers. Forestry 2010, 83, 527–533. [Google Scholar] [CrossRef]
- Wahlsteen, E.; Avramidou, E.V.; Bozic, G.; Mediouni, R.M.; Schuldt, B.; Sobolewska, H. Continental-wide population genetics and post-Pleistocene range expansion in field maple (Acer campestre L.), a subdominant temperate broadleaved tree species. Tree Genet. Genomes 2023, 19, 15. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant endemism centres and biodiversity hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef]
- Elhag, M.; Yimaz, N.; Bahrawi, J.; Boteva, S. Evaluation of optical remote sensing data in burned areas mapping of Thasos Island, Greece. Earth Syst. Environ. 2020, 4, 813–826. [Google Scholar] [CrossRef]
- Spanos, I.A.; Daskalakou, E.N.; Thanos, C.A. Postfire, natural regeneration of Pinus brutia forests in Thasos island, Greece. Acta Oecol. 2000, 21, 13–20. [Google Scholar] [CrossRef]
- Chebykina, E.; Abakumov, E. Essential Role of Forest Fires in Humic Acids Structure and Composition Alteration. Agronomy 2022, 12, 2910. [Google Scholar] [CrossRef]
- Kelepertsis, A.; Bibou, A. Heavy metal contamination of soils at old mining sites on Thasos Island, Greece. Environ. Geochem. Health 1991, 13, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, M.; Sebastiani, F.; Mayol, M.; González-Martínez, S.C.; Riba, M.; Vendramin, G.G. Isolation and characterization of polymorphic nuclear microsatellite loci in Taxus baccata L. Conserv. Genet. 2008, 9, 1665–1668. [Google Scholar] [CrossRef]
- Mahmoodi, P.; Khayam Nekouei, S.M.; Mardi, M.; Pirseyedi, S.M.; Ghaffari, M.R.; Ramroodi, M.; Siahsar, B.A. Isolation and characterization of new microsatellite marker in Taxus baccata L. Conserv. Genet. Resour. 2010, 2, 195–199. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. An exact test for population differentiation. Evolution 1995, 49, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T. hp-rare 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Notes 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L. Maximum likelihood estimation of the frequency of null alleles at microsatellite loci. Conserv. Genet. 2006, 7, 991. [Google Scholar] [CrossRef]
- Marshall, T.; Slate, J.; Kruuk, L.; Pemberton, J. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chybicki, I.J.; Oleksa, A.; Kowalkowska, K. Variable rates of random genetic drift in protected populations of English yew: Implications for gene pool conservation. Conserv. Genet. 2012, 13, 899–911. [Google Scholar] [CrossRef]
- Lewandowski, A.; Burczyk, J.; Mejnartowicz, L. Genetic structure of English yew (Taxus baccata L.) in the Wierzchlas Reserve: Implications for genetic conservation. For. Ecol. Manag. 1995, 73, 221–227. [Google Scholar] [CrossRef]
- Litkowiec, M.; Lewandowski, A.; Wachowiak, W. Genetic variation in Taxus baccata L.: A case study supporting Poland’s protection and restoration program. For. Ecol. Manag. 2018, 409, 148–160. [Google Scholar] [CrossRef]
- Tröber, U.; Ballian, D. Genetic characterization of English yew (Taxus baccata L.) populations in Bosnia and Herzegovina. Eur. J. For. Res. 2011, 130, 479–489. [Google Scholar] [CrossRef]
- Hilfiker, K.; Holderegger, R.; Rotach, P.; Gugerli, F. Dynamics of genetic variation in Taxus baccata: Local versus regional perspectives. Can. J. Bot. 2004, 82, 219–227. [Google Scholar] [CrossRef]
- Hilfiker, K.; Gugerli, F.; Schütz, J.-P.; Rotach, P.; Holderegger, R. Low RAPD variation and female-biased sex ratio indicate genetic drift in small populations of the dioecious conifer Taxus baccata in Switzerland. Conserv. Genet. 2004, 5, 357–365. [Google Scholar] [CrossRef]
- Zarek, M. RAPD analysis of genetic structure in four natural populations of Taxus baccata from Southern Poland. Acta Biol. Cracoviensia Ser. Bot. 2009, 51, 67–75. [Google Scholar]
- Stefanović, M.; Nikolić, B.; Matić, R.; Popović, Z.; Vidaković, V.; Bojović, S. Exploration of sexual dimorphism of Taxus baccata L. needles in natural populations. Trees 2017, 31, 1697–1710. [Google Scholar] [CrossRef]
- Tumpa, K.; Liber, Z.; Šatović, Z.; Medak, J.; Idžojtić, M.; Vidaković, A.; Vukelić, J.; Šapić, I.; Nikl, P.; Poljak, I. High level of phenotypic differentiation of common yew (Taxus baccata L.) populations in the north-western part of the Balkan Peninsula. Forests 2022, 13, 78. [Google Scholar] [CrossRef]
- Paule, L.; Gömöry, D.; Longauer, R. Present distribution and ecological conditions of the English yew (Taxus baccata L.) in Europe. In Proceedings of the International Yew Resources Conference: Yew (Taxus) Conservation Biology and Interactions, Berkeley, CA, USA, 12–13 March 1993; pp. 189–196. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Citation | Motif | Size Range (bp) | Annealing Temperature °C |
|---|---|---|---|---|
| Tax23 | [51] | (GT)21 | 153–183 | 60 |
| Tax26 | [51] | (GT)30 | 208–284 | 60 |
| Tax36 | [51] | (GT)25 | 126–262 | 60 |
| Tax86 | [51] | (GT)34 | 152–304 | 60 |
| Tax92 | [51] | (GT)24 | 160–280 | 60 |
| ABRII-TB50 | [52] | (TC)8-(CT)13 | 270–330 | 60 |
| ABRII-TB56 | [52] | (TG)6-(CTAGAT)2 | 204–270 | 60 |
| ABRII-TB58 | [52] | (CTATCC)2-(TCTGTA)2 | 206–230 | 60 |
| Locus | Na | Ne | I | Ho | He | Fst | Fis | fn | PIC |
|---|---|---|---|---|---|---|---|---|---|
| Tax23 | 7 | 2.457 | 1.031 | 0.250 | 0.591 | 0.016 | 0.577 | 0.006 | 0.540 |
| Tax26 | 8 | 3.297 | 1.271 | 0.714 | 0.691 | 0.038 | −0.033 | 0.41 | 0.667 |
| Tax36 | 16 | 5.494 | 1.863 | 0.607 | 0.811 | 0.035 | 0.252 | 0.096 | 0.823 |
| Tax86 | 11 | 3.741 | 1.429 | 0.286 | 0.693 | 0.092 | 0.587 | 0.000 | 0.733 |
| Tax92 | 15 | 4.467 | 1.680 | 0.679 | 0.772 | 0.081 | 0.121 | 0.036 | 0.541 |
| ABRII-TB50 | 7 | 2.338 | 0.969 | 0.250 | 0.564 | 0.117 | 0.557 | 0.043 | 0.819 |
| ABRII-TB56 | 4 | 1.960 | 0.683 | 0.857 | 0.490 | 0.020 | -0.750 | 0.987 | 0.568 |
| ABRII-TB58 | 6 | 2.405 | 0.966 | 0.250 | 0.584 | 0.050 | 0.572 | 0.038 | 0.375 |
| Population | Ne | He | I | AR | PAR | Fst |
|---|---|---|---|---|---|---|
| A | 3.299 | 0.663 | 1.270 | 3.48 | 1.13 | 0.199 |
| B | 3.241 | 0.636 | 1.203 | 3.31 | 0.95 | 0.251 |
| Mean | 3.270 | 0.649 | 1.236 | 3.395 | 1.04 | 0.225 |
| Source | df | SS | MS | Est. Var. | % | ΦST |
|---|---|---|---|---|---|---|
| Among Pops | 1 | 17.893 | 17.893 | 0.778 | 10 | 0.067 |
| Within Pops | 26 | 181.929 | 6.997 | 6.997 | 90 | |
| Total | 27 | 199.821 | 7.776 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malliarou, E.; Avramidou, E.V.; Ranis, G.D.; Bountis, D.I. Towards the Conservation of Monumental Taxus baccata L. Trees of Thasos Island: Genetic Insights. Forests 2023, 14, 2384. https://doi.org/10.3390/f14122384
Malliarou E, Avramidou EV, Ranis GD, Bountis DI. Towards the Conservation of Monumental Taxus baccata L. Trees of Thasos Island: Genetic Insights. Forests. 2023; 14(12):2384. https://doi.org/10.3390/f14122384
Chicago/Turabian StyleMalliarou, Ermioni, Evangelia V. Avramidou, Georgios D. Ranis, and Diamantis I. Bountis. 2023. "Towards the Conservation of Monumental Taxus baccata L. Trees of Thasos Island: Genetic Insights" Forests 14, no. 12: 2384. https://doi.org/10.3390/f14122384
APA StyleMalliarou, E., Avramidou, E. V., Ranis, G. D., & Bountis, D. I. (2023). Towards the Conservation of Monumental Taxus baccata L. Trees of Thasos Island: Genetic Insights. Forests, 14(12), 2384. https://doi.org/10.3390/f14122384