
Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review
 ,
,  and
and
Abstract
:1. Introduction
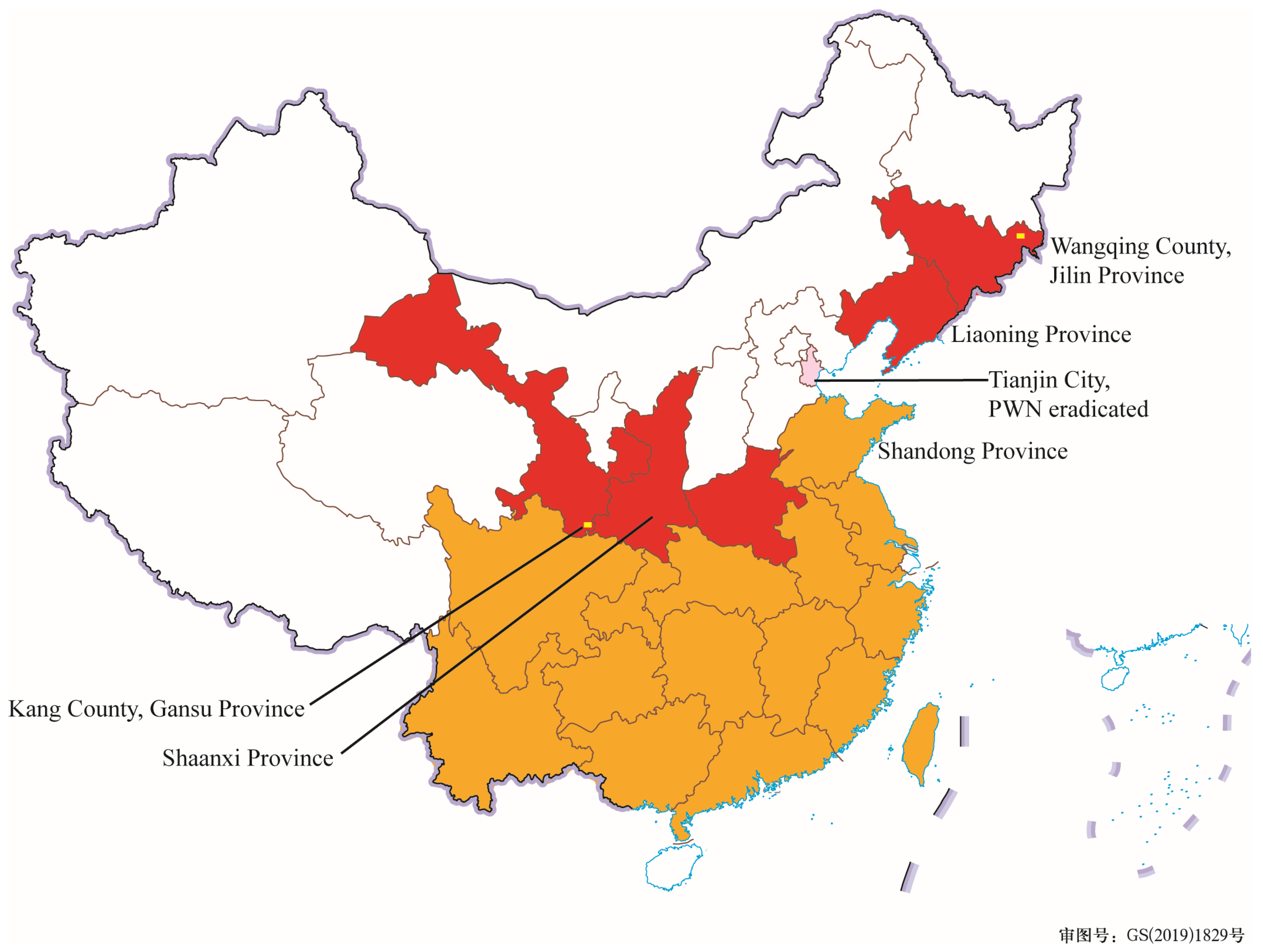
2. Trends of the Spread of PWD in Northeast and Northwest China
3. The Complexity of PWD
3.1. Complex Plant Disease System
3.2. Pathogenic Differentiation in Different Regions
3.3. Unclear Pathogenesis
4. PWN and Influencing Environmental Conditions
4.1. Temperature
4.2. Altitude
4.3. Other Environmental Conditions
5. PWN and Its Host Trees
5.1. PWN Host Trees and Non-Host Trees in China
5.2. Pinaceae in Northeast and Northwest China
6. PWN and Its Vector Insects
6.1. Vector Insects of PWNs Worldwide
6.2. Monochamus Species in Northeast and Northwest China
7. Comprehensive Risk of PWD in Northeast and Northwest China
7.1. Comprehensive Risk Analysis
7.2. Scientific Bottlenecks to Be Proven
7.2.1. The Pathogenicity of PWNs to Picea and Abies
7.2.2. The Distribution and PWN Transmissibility of Monochamus
7.3. Future PWD’s Management in Northeast and Northwest China
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steiner, G.; Buhrer, E.M. Aphelenchoides xylophilus n. sp., a nematode associated with blue-stain and other fungi in timber. J. Agric. Res. 1934, 48, 949–951. [Google Scholar]
- Bowers, W.; Hudak, J.; Raske, A.; Magasi, L.; Myren, D.; Lachance, D.; Cerezke, H.; Van Sickle, G. Host and Vector Surveys for the Pinewood Nematode, Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle (Nematoda: Aphelenchoididae) in Canada; Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 1992. [Google Scholar]
- Dwinell, L. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar] [CrossRef]
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. (In Korean) [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, causal agent of pine wilt disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Ruan, X.S. Occurrence and control of pine wood nematode in Taiwan. Plant Quar. 1996, 18, 86–87. (In Chinese) [Google Scholar]
- Winney, R. Preliminary observation on pine wood nematode in Hong Kong. Guangdong For. Sci. Technol. 1987, 16, 46–50. (In Chinese) [Google Scholar]
- Sun, Y.C. Pine wood nematode found in Sun Yat-sen Mausoleum, Nanjing. J. Jiangsu For. Sci. Technol. 1982, 9, 47. (In Chinese) [Google Scholar]
- Ye, J.R. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. (In Chinese) [Google Scholar]
- Zhang, X.; Zhao, J.J.; Yan, J.; Fang, G.F.; Huang, J.X. Economic loss assessment of pine wilt disease in Mainland China in 2017. J. Beijing For. Univ. 2020, 42, 96–106. (In Chinese) [Google Scholar]
- Zhao, J.J.; Huang, J.X.; Yan, J.; Fang, G.F. Economic loss of pine wood nematode disease in Mainland China from 1998 to 2017. Forests 2020, 11, 1042. [Google Scholar] [CrossRef]
- Song, Y.S.; Zang, X.Q. Adaptability analysis of pine wood nematode in China and preliminary study on quarantine countermeasures. For. Pest Dis. 1989, 8, 38–41. (In Chinese) [Google Scholar]
- Pan, L.; Cui, R.; Li, Y.; Feng, Y.; Zhang, X. Investigation of pinewood nematodes in Pinus tabuliformis Carr. under low-temperature conditions in Fushun, China. Forests 2020, 11, 993. [Google Scholar] [CrossRef]
- Yu, H.Y.; Wu, H. New host plant and new vector insect of pine wood nematode found in Liaoning Province. For. Pest Dis. 2018, 37, 61. (In Chinese) [Google Scholar]
- Yu, H.Y.; Wu, H.; Zhang, X.D.; Wang, L.M.; Zhang, X.F.; Song, Y.S. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. (In Chinese) [Google Scholar]
- Li, M.; Li, H.; Sheng, R.C.; Sun, H.; Sun, S.H.; Chen, F.M. The First record of Monochamus saltuarius (Coleoptera; Cerambycidae) as vector of Bursaphelenchus xylophilus and its new potential hosts in China. Insects 2020, 11, 636. [Google Scholar] [CrossRef]
- Lv, Q.; Wang, W.D.; Liang, J.; Yan, D.H.; Jia, X.Z.; Zhang, X.Y. Potential suitability assessment of Bursaphelenchus xylophilus in China. For. Res. 2005, 18, 460–464. (In Chinese) [Google Scholar]
- Pan, H.W. Study on the potential geographic distribution of Bursaphelenchus xylophilus in. China. Dissertation, China Academy of Forestry, Beijing, China, 2009. (In Chinese). [Google Scholar]
- Zhang, X.Y.; Lv, Q.; Feng, Y.M.; Shi, L.; Liang, J.; Yan, D.H.; Liu, Z.Y. Risk Assessment and Countermeasures of Pine wood Nematode Disease in China; Science Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- He, S.Y.; Wen, J.B.; Luo, Y.Q.; Zong, S.X.; Zhao, Y.X.; Han, J. The predicted geographical distribution of Bursaphelenchus xylophilus in China under climate warming. Chin. J. Appl. Entomol. 2012, 49, 236–243. (In Chinese) [Google Scholar]
- Zhao, J.; Han, X.; Shi, J. Potential distribution of Bursaphelenchus xylophilus in China due to adaptation cold conditions. J. Biosaf. 2017, 26, 191–198. (In Chinese) [Google Scholar]
- Yu, Z.J.; Li, S.; Zhou, Y.T.; Zhou, R.L.; Wang, Y.X. Spatial estimation and prediction of suitable distribution of Bursaphelenchus xylophilus with different warming modes in China. J. Northeast For. Univ. 2018, 46, 85–91. (In Chinese) [Google Scholar]
- Han, Y.Y.; Wang, Y.; Xiang, Y.; Ye, J.R. Prediction of potential distribution of Bursaphelenchus xylophilus in China based on Maxent ecological niche model. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2015, 39, 6–10. (In Chinese) [Google Scholar]
- Tang, X.; Yuan, Y.; Li, X.; Zhang, J. Maximum entropy modeling to predict the impact of climate change on pine wilt disease in China. Front Plant Sci 2021, 12, 652500. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sun, H.; Zhou, Y.T.; Li, X.D.; Yu, Z.J.; Dong, Z.H. Occurrence of major forest pests in 2021 and prediction of occurrence trend in 2022. For. Pest Dis. 2022, 41, 44–47. (In Chinese) [Google Scholar]
- Song, Y.S. Pine Wilt Disease in China: Thirty Years Review and Thinking; Northeast Forestry University Press: Harbin, China, 2013. (In Chinese) [Google Scholar]
- General Station of Forest and Grassland Pest Control of National Forestry and Grassland Administration; Nanjing Forestry University. Occurrence Regularity and Control Techniques of Pine Wilt Disease in China; China Forestry Publishing House: Beijing, China, 2019. (In Chinese) [Google Scholar]
- Ding, X.; Guo, Y.; Ye, J.; Wu, X.; Lin, S.; Chen, F.; Zhu, L.; Huang, L.; Song, X.; Zhang, Y.; et al. Population differentiation and epidemic tracking of Bursaphelenchus xylophilus in China based on chromosome-level assembly and whole-genome sequencing data. Pest Manag. Sci. 2022, 78, 1213–1226. [Google Scholar] [CrossRef]
- Kanzaki, N.; Giblin-Davis, R.M. Diversity and plant pathogenicity of Bursaphelenchus and related nematodes in relation to their vector bionomics. Curr. For. Rep. 2018, 4, 85–100. [Google Scholar] [CrossRef]
- Ryss, A.; Vieira, P.; Mota, M.; Kulinich, O. A synopsis of the genus Bursaphelenchus Fuchs, 1937 (Aphelenchida: Parasitaphelenchidae) with keys to species. Nematology 2005, 7, 393–458. [Google Scholar] [CrossRef]
- Wang, H.Y. Biodiversity and Biogeography of the Genus Bursaphelenchus Fuchs, 1937 (Nematoda: Parasitaphelenchidae). Doctoral Dissertation, Fujian Agriculture and Forestry University, Fuzhou, China, 2008. (In Chinese). [Google Scholar]
- Aikawa, T. Transmission biology of Bursaphelenchus xylophilus in relation to its insect vector. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 123–138. [Google Scholar]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, X.; Wei, Y.; Zhou, J.; Zhang, W.; Qin, P.; Chinta, S.; Kong, X.; Liu, Y.; Yu, H.; et al. Ascarosides coordinate the dispersal of a plant-parasitic nematode with the metamorphosis of its vector beetle. Nat. Commun. 2016, 7, 12341. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, F.X.; Hasegawa, K.; Mota, M.; Vicente, C.S. Bacterial role in pine wilt disease development–review and future perspectives. Environ. Microbiol. Rep. 2015, 7, 51–63. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inacio, M.L. Insights into the role of fungi in pine wilt disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.H.; Zhang, B.; Sun, J.H. Research progress on the interaction between associated microbes and pine wood nematode-vector beetle complex. For. Pest Dis. 2022, 41, 30–37. (In Chinese) [Google Scholar]
- Proenca, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiologyopen 2017, 6, e00415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.N.; Wang, L.; Shi, X.F.; Luo, L.; Ye, J.R. Negative effects of free-living nematodes on the populations of Bursaphelenchus xylophilus in dead pine trees. Biol. Control 2022, 168, 104858. [Google Scholar] [CrossRef]
- Akbulut, S.; Stamps, W.T. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Kwon, H.J.; Jung, J.K.; Jung, C.; Han, H.; Koh, S.H. Dispersal capacity of Monochamus saltuarius on flight mills. Entomol. Exp. Appl. 2018, 166, 420–427. [Google Scholar] [CrossRef]
- David, G.; Giffard, B.; Piou, D.; Jactel, H. Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. J. Appl. Entomol. 2014, 138, 566–576. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhang, R.Z.; Chen, J.Y. Species and their dispersal ability of Monochamus as vectors to transmit Bursaphelenchus xylophilus. J. Zhejiang A F Univ. 2007, 24, 350–356. (In Chinese) [Google Scholar]
- Li, H. Identification and Pathogenicity of Bursaphelenchus Species (Nematoda: Parasitaphelenchidae). Doctoral Dissertation, Ghent University, Ghent, Belgium, 2008. [Google Scholar]
- Futai, K.; Sutherland, J.R. Pathogenicity and attraction to host extracts of Canadian pinewood nematodes: Studies with Scots pine, western larch, and black spruce seedlings. Can. J. For. Res. 1989, 19, 1256–1261. [Google Scholar] [CrossRef]
- Kiyohara, T.; Bolla, R. Pathogenic variability among populations of the pinewood nematode, Bursaphelenchus xylophilus. For. Sci. 1990, 36, 1061–1076. [Google Scholar] [CrossRef]
- Liu, K.C.; Zeng, F.L.; Ben, A.L.; Han, Z.M. Pathogenicity and repulsion for toxin-producing bacteria of dominant bacteria on the surface of American pine wood nematodes. J. Phytopathol. 2017, 165, 580–588. [Google Scholar] [CrossRef]
- Jiao, G.Y.; Shen, P.Y.; Li, H.M.; Xu, P.F. Study on pathogenicity of Japanese pine wood nematode strain and Nanjing pine wood nematode strain to Cedrus deodara and Pinus massoniana. Plant Quar. 1996, 18, 193–195. (In Chinese) [Google Scholar]
- Ju, Y.W.; Zhao, B.G.; Pu, C.H. Different virulence of pine wood nematode from Japan and China on Cedrus deodara. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2007, 31, 139–140. (In Chinese) [Google Scholar]
- Zheng, J.R. Discussion on the Mechanism of Cedar’s Immunity to Pine Wood Nematode under Natural Conditions in China. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2007. (In Chinese). [Google Scholar]
- Zhen, W.; Chun, Y.W.; Zhe, M.F.; Dong, L.Z.; Lei, L.; Mi, R.L.; Zheng, L.; Jing, J.L.; Chang, K.S. Advances in research of pathogenic mechanism of pine wilt disease. Afr. J. Microbiol. Res. 2010, 4, 437–442. [Google Scholar] [CrossRef]
- Li, Y.X.; Wang, X.; Liu, Z.K.; Zhang, X.Y. Research advance of pathogenic mechanism of pine wood nematode. For. Pest Dis. 2022, 41, 11–20. (In Chinese) [Google Scholar]
- Shinya, R.; Morisaka, H.; Takeuchi, Y.; Futai, K.; Ueda, M. Making headway in understanding pine wilt disease: What do we perceive in the postgenomic era? J. Biosci. Bioeng. 2013, 116, 1–8. [Google Scholar] [CrossRef]
- Wen, T.Y.; Wu, X.Q.; Hu, L.J.; Qiu, Y.J.; Rui, L.; Zhang, Y.; Ding, X.L.; Ye, J.R. A novel pine wood nematode effector, BxSCD1, suppresses plant immunity and interacts with an ethylene-forming enzyme in pine. Mol. Plant Pathol. 2021, 22, 1399–1412. [Google Scholar] [CrossRef]
- Wang, R.; Li, Y.; Li, D.; Zhang, W.; Wang, X.; Wen, X.; Liu, Z.; Feng, Y.; Zhang, X. Identification of the Extracellular Nuclease Influencing Soaking RNA Interference Efficiency in Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2022, 23, 12278. [Google Scholar] [CrossRef]
- Rutherford, T.; Webster, J. Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Fan, H.J.; Guan, Z.X. Analysis on the changes of meteorological elements in Wangqing county in the last 52 years. Jilin Meteorol. 2010, 17, 15–18. (In Chinese) [Google Scholar]
- Wharton, D.A.; Goodall, G.; Marshall, C.J. Freezing survival and cryoprotective dehydration as cold tolerance mechanisms in the Antarctic nematode Panagrolaimus davidi. J. Exp. Biol. 2003, 206, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.R.; Wharton, D.A. The ability of the Antarctic nematode Panagrolaimus davidi to survive intracellular freezing is dependent upon nutritional status. J. Comp. Physiol. B 2013, 183, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Shatilovich, A.; Tchesunov, A.; Neretina, T.; Grabarnik, I.; Gubin, S.; Vishnivetskaya, T.; Onstott, T.C.; Rivkina, E. Viable nematodes from late Pleistocene permafrost of the Kolyma river lowland. Dokl. Biol. Sci. 2018, 480, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.R.; Fiedler, T.J.; Harris, T.W.; Carvalho, F.; Antoshechkin, I.; Han, M.; Sternberg, P.W.; Stein, L.D.; Chalfie, M. WormBook: The online review of Caenorhabditis elegans biology. Nucleic Acids Res. 2007, 35, D472–D475. [Google Scholar] [CrossRef]
- Szewczyk, N.J.; Mancinelli, R.L.; McLamb, W.; Reed, D.; Blumberg, B.S.; Conley, C.A. Caenorhabditis elegans survives atmospheric breakup of STS-107, space shuttle Columbia. Astrobiology 2005, 5, 690–705. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, N.; Futai, K. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus group. Nematology 2002, 4, 35–41. [Google Scholar] [CrossRef]
- Li, Z.X.; Tao, J.; Zong, S.X. Cold tolerance in pinewood nematode Bursaphelenchus xylophilus promoted multiple invasion events in mid-temperate zone of China. Forests 2022, 13, 1100. [Google Scholar] [CrossRef]
- Pan, L.; Cui, R.; Li, Y.X.; Zhang, W.; Bai, J.W.; Li, J.W.; Zhang, X.Y. Third-Stage dispersal juveniles of Bursaphelenchus xylophilus can resist low-temperature stress by entering cryptobiosis. Biology 2021, 10, 785. [Google Scholar] [CrossRef]
- Kong, Q.Q.; Ding, X.L.; Chen, Y.F.; Ye, J.R. Comparison of morphological indexes and the pathogenicity of Bursaphelenchus xylophilus in northern and southern China. Forests 2021, 12, 310. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhang, X.Y. Analysis on the trend of invasion and expansion of Bursaphelenchus xylophilus. For. Pest Dis. 2018, 37, 1–4. (In Chinese) [Google Scholar]
- Yik, C.P.; Birchfield, W. Observations on the morphology of the pine wood nematode, Bursaphelenchus xylophilus. J. Nematol. 1981, 13, 376–384. [Google Scholar] [PubMed]
- Dropkin, V.; Foudin, A.; Kondo, E.; Linit, M.; Smith, M.; Robbins, K. Pinewood nematode: A threat to US forests? [Bursaphelenchus xylophilus]. Plant Dis. 1981, 65, 1022–1027. [Google Scholar] [CrossRef]
- Wingfield, M.; Blanchette, R. The pine-wood nematode, Bursaphelenchus xylophilus, in Minnesota and Wisconsin: Insect associates and transmission studies. Can. J. For. Res. 1983, 13, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Blunt, T.D.; Jacobi, W.R.; Appel, J.A.; Tisserat, N.; Todd, T.C. First report of pine wilt in Colorado, USA. Plant Health Prog. 2014, 15, 103–104. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, M.; Akiba, M. Possible altitude and temperature limits on pine wilt disease: The reproduction of vector sawyer beetles (Monochamus alternatus), survival of causal nematode (Bursaphelenchus xylophilus), and occurrence of damage caused by the disease. Eur. J. For. Res. 2014, 133, 225–233. [Google Scholar] [CrossRef]
- Haran, J.; Roques, A.; Bernard, A.; Robinet, C.; Roux, G. Altitudinal barrier to the spread of an invasive species: Could the Pyrenean chain slow the natural spread of the pinewood nematode? PLoS ONE 2015, 10, e0134126. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y. The biogeography and economic geography of the pine wood nematode in China. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2011, 35, 144–145. (In Chinese) [Google Scholar]
- Fu, L.; Li, N.; Elias, T.; Mill, R. Pinaceae. In Flora of China 4; Flora of China Editorial Committee; Science Press: Beijing, China, 1999; pp. 9–10. [Google Scholar]
- Ye, J.R.; Wu, X.Q. Research progress of pine wilt disease. For. Pest Dis. 2022, 41, 1–10. (In Chinese) [Google Scholar]
- Ge, J.J.; Bai, X.Y.; Cheng, H.R. Resistance of Cedrus deodara and Pinus taiwanensis to pine wood nematode (Bursapherenchus xylophilus). Acta Phytopathol. Sin. 1992, 23, 335–338. (In Chinese) [Google Scholar]
- Editorial Committee of FRPS Chinese Academy of Sciences. Flora Reipublicae Popularis Sinicae Volume 7: Gymnosperm; Science Press: Beijing, China, 1978. (In Chinese) [Google Scholar]
- Sutherland, J.R.; Ring, F.M.; Seed, J.E. Canadian conifers as hosts of the pinewood nematode (Bursaphelenchus xylophilus): Results of seedling inoculations. Scand. J. For. Res. 1991, 6, 209–216. [Google Scholar] [CrossRef]
- Yang, B.J.; Hu, K.J.; Wang, Q.L.; Sun, Y.L.; Wu, Z.M.; Wang, Q.M. Studies on the resistance of pine trees to pine wood nematode, Bursaphelenchus xylophilus. For. Res. 1993, 6, 249–255. (In Chinese) [Google Scholar]
- Wingfield, M.; Blanchette, R.; Kondo, E. The pine wood nematode on balsam fir in Minnesota. Phytopatholgy 1982, 72, 965. [Google Scholar]
- Kondo, E.; Foudin, A.; Linit, M.; Smith, M.; Bolla, R.; Winter, R.; Dropkin, V. Pine wilt Disease-Nematological, Entomological, and Biochemical Investigations; University of Missouri-Columbia Agricultural Experiment Station: Columbia, SC, USA, 1982. [Google Scholar]
- Kasuga, H.; Togashi, K. Nine-month persistence of Bursaphelenchus xylophilus in living Abies sachalinensis and Picea jezoensis seedlings and nematode reproduction in stem sections. Nematology 2012, 14, 547–554. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Lima, M.; Vasconcelos, M.W. Susceptibility evaluation of Picea abies and Cupressus lusitanica to the pine wood nematode (Bursaphelenchus xylophilus). Plant Pathol. 2013, 62, 1398–1406. [Google Scholar] [CrossRef]
- Editorial Committee of China Forest. China Forest Volume 1 General; China Forestry Publishing House: Beijing, China, 1997. (In Chinese) [Google Scholar]
- Editorial Committee of China Forest. China Forest Volume 2 Coniferous Forest; China Forestry Publishing House: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Cesari, M.; Marescalchi, O.; Francardi, V.; Mantovani, B. Taxonomy and phylogeny of European Monochamus species: First molecular and karyological data. J. Zool. Syst. Evol. Res. 2005, 43, 1–7. [Google Scholar] [CrossRef]
- Wang, Q. Biogeography study of Monochamus Guér. Sci. Silvae Sin. 1988, 24, 297–302. (In Chinese) [Google Scholar]
- Zheng, S.Z.; Zhang, K.; Yang, X.J.; Xu, M.; An, Y.L. Sequence comparison and phylogenetic analysis of mt DNA CO I gene sequence fragments in Monochamus species (Coleoptera: Cerambycidae). Chin. J. Appl. Entomol. 2015, 52, 370–381. (In Chinese) [Google Scholar]
- General Station of Forest and Grassland Pest Control of National Forestry and Grassland Administration. Forest pest in China 2014–2017 National Forest Pest Survey Results; China Forestry Publishing House: Beijing, China, 2019. (In Chinese) [Google Scholar]
- General Station of Forest Pest Control of State Forestry Administration. Overview of Forestry Pests in China: Compilation of National Forestry Pest Survey Results from 2003 to 2007; China Forestry Publishing House: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Department of Forestry Administration and Protection of the Ministry of Forestry of the People’s Republic of China. List of Forest Pest and Disease Survey in China; Department of Forestry Administration and Protection of the Ministry of Forestry: Beijing, China, 1988. (In Chinese)
- Shanghai Landscape Gardening Research Institute. Survey of Plant Diseases, Pests and Natural Enemies of Garden Plants in China and Study on Quarantine Pests (II); Shanghai Landscape Gardening Research Institute: Shanghai, China, 1987. (In Chinese) [Google Scholar]
- Hua, L.Z. List of Cerambycidae Insects in China; Sun Yatsen University Press: Guangzhou, China, 1982. (In Chinese) [Google Scholar]
- Chen, S.X.; Xie, Y.Z.; Deng, G.F. Economic Insects of China Vol. 1 Coleoptera Cerambycidae; Science Press: Beijing, China, 1959. (In Chinese) [Google Scholar]
- Pu, F.J. Economic Insects of China Vol. 19 Coleoptera Cerambycidae 2; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Jiang, S.N.; Pu, F.J.; Hua, L.Z. Economic Insects of China Vol. 35 Coleoptera Cerambycidae 3; Science Press: Beijing China, 1985. (In Chinese) [Google Scholar]
- Wang, Z.M.; Pi, Z.Q.; Hou, B. Monochamus alternatus found in Jilin Province. For. Pest Dis. 2006, 25, 35. (In Chinese) [Google Scholar]
- Pajares, J.A.; Alvarez, G.; Hall, D.R.; Ibarra, N.; Hoch, G.; Halbig, P.; Cocos, D.; Johansson, H.; Schroeder, M. Attractants for management of the pine sawyer beetle Monochamus sutor, a potential vector of Bursaphelenchus xylophilus. J. Appl. Entomol. 2017, 141, 97–111. [Google Scholar] [CrossRef]
- Schroeder, M. Trapping strategy for Monochamus sutor and Monochamus galloprovincialis: Potential vectors of the pine wood nematode in Scandinavia. Agric. For. Entomol. 2019, 21, 372–378. [Google Scholar] [CrossRef]
- Wang, K.; Li, L.J.; Yu, F.; Wang, L.G.; Yang, H.J. Analysis and recommendations about epidemic situation of intercepted longhorn beetles at China’s ports. For. Pest Dis. 2018, 37, 22–26. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
| Country | Isolation | Seedling Inoculation |
|---|---|---|
| USA (Origin) | Picea glauca, Picea pungens, Abies balsamea [33,84,85] | |
| Canada (Origin) | Picea glauca, Picea mariana, Picea rubens, Abies balsamea [2] | Picea engelmannii, Picea glauca, Picea mariana, Picea rubens, Picea sitchensis, Abies amabilis, Abies balsamea, Abies grandis [82,83] |
| Japan (Invasive) | Picea abies, Abies firma [4,5] | Picea abies, Picea jezoensis, Picea mariana, Abies homolepis, Abies sachalinensis [5,86] |
| Species | Distribution in Northeast (NE) and Northwest (NW) China | Distribution | Lowest Distribution Altitude (Meters) |
|---|---|---|---|
| Pinus densiflora var. ussuriensis | NE | Heilongjiang Province | 50 |
| Pinus koraiensis | NE | Lesser Khingan (Heilongjiang Province), Wanda Mountains (Heilongjiang Province), Zhangguangcai Range (Heilongjiang and Jilin provinces), Changbai Mountain (Jilin Province) | 50 |
| Pinus sylvestris var. sylvestriformis | NE | Changbai Mountain | 630 |
| Pinus thunbergii | NE | Liaoning Province | 400 |
| Larix gmelinii | NE | Greater Khingan (Heilongjiang Province), Lesser Khingan | 300 |
| Larix olgensis | NE | Changbai Mountain | 500 |
| Picea jezoensis | NE | Greater Khingan, Lesser Khingan, Zhangguangcai Range, Changbai Mountain | 700 |
| Picea koraiensis | NE | Greater Khingan, Lesser Khingan, Wanda Mountains, Zhangguangcai Range, Changbai Mountain | 300 |
| Abies nephrolepis | NE | Lesser Khingan, Zhangguangcai Range, Changbai Mountain | 500 |
| Species | Distribution in Northeast (NE) and Northwest (NW) China | Relationship with PWN (Pine Wood Nematode) |
|---|---|---|
| Monochamus alternatus | NE and NW | Confirmed vector in China [43] |
| Monochamus basifossulatus | No distribution | |
| Monochamus galloprovincialis | NE | Confirmed vector in Europe [43] |
| Monochamus guerryi | No distribution | |
| Monochamus saltuarius | NE and NW | Confirmed vector in China [19] |
| Monochamus sartor urussovii | NE and NW | |
| Monochamus sutor | NE and NW |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests 2023, 14, 174. https://doi.org/10.3390/f14020174
Xu Q, Zhang X, Li J, Ren J, Ren L, Luo Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests. 2023; 14(2):174. https://doi.org/10.3390/f14020174
Chicago/Turabian StyleXu, Qinwang, Xuejiao Zhang, Jiaxing Li, Jiaru Ren, Lili Ren, and Youqing Luo. 2023. "Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review" Forests 14, no. 2: 174. https://doi.org/10.3390/f14020174
APA StyleXu, Q., Zhang, X., Li, J., Ren, J., Ren, L., & Luo, Y. (2023). Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests, 14(2), 174. https://doi.org/10.3390/f14020174