
Function of Tryptophan 2,3-Dioxygenase in Monochamus alternatus Hope Revealed by RNA Interference
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Identification of the TDO Gene in M. alternatus
2.3. Isolation of M. alternatus RNA and Synthesis of dsRNA Molecules
2.4. RNAi in M. alternatus in the Pupal Stage
2.5. Real-Time Quantitative PCR (RT-qPCR)
2.6. Statistical Analysis
3. Results
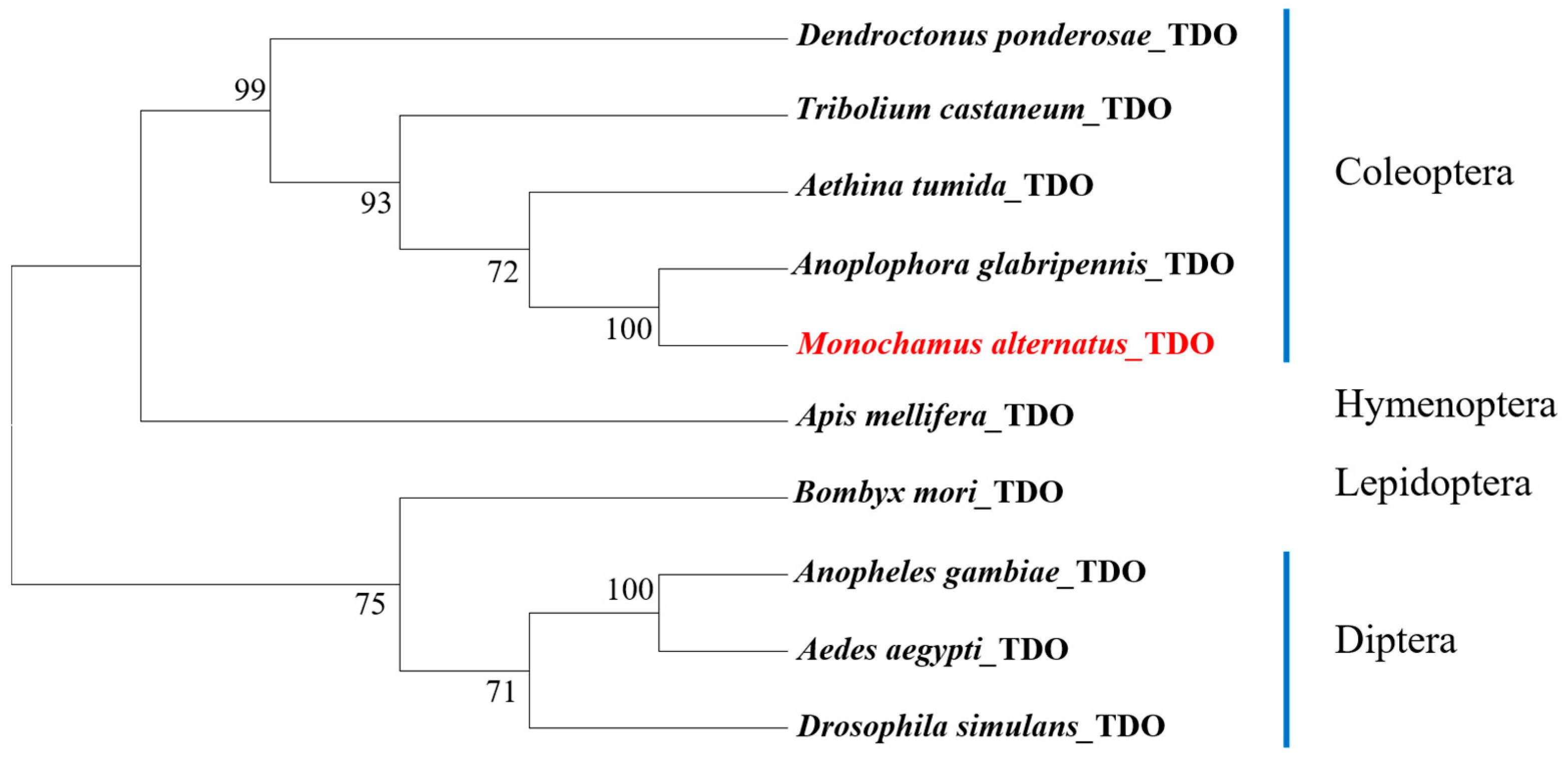
3.1. Identification of the TDO Gene and Homology Analysis
3.2. TDO Silencing and Its Phenotypic Impact in M. alternatus
3.3. Gene Expression of TDO during Different Developmental Stages of M. alternatus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Li, H.; Sheng, R.C.; Sun, H.; Sun, S.H.; Chen, F.M. The first record of Monochamus saltuarius (Coleoptera; Cerambycidae) as vector of Bursaphelenchus xylophilus and its new potential hosts in China. Insects 2020, 11, 636. [Google Scholar] [CrossRef]
- Shi, J.; Luo, Y.; Wu, H.; Kari, H.; Liang, L. Impact of the invasion by Bursaphelenchus xylophilus on forest growth and related growth models of Pinus massoniana population. Acta Ecol. Sin. 2008, 28, 3193–3204. [Google Scholar]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Kim, H.M.; Choi, I.S.; Lee, S.; Hwang, I.M.; Chun, H.H.; Wi, S.G.; Kim, J.C.; Shin, T.Y.; Kim, J.C.; Kim, J.S.; et al. Advanced strategy to produce insecticidal destruxins from lignocellulosic biomass Miscanthus. Biotechnol. Biofuels 2019, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Merchant, A.; Zhao, Z.; Zhang, Y.; Zhang, J.; Zhang, Q.; Wang, Q.; Zhou, X.; Li, X. Characterization of MaltOBP1, a minus-C odorant-binding protein, from the Japanese pine sawyer beetle, Monochamus alternatus Hope (Coleoptera: Cerambycidae). Front. Physiol. 2020, 11, 212. [Google Scholar] [CrossRef] [Green Version]
- Tittiger, C. Functional genomics and insect chemical ecology. J. Chem. Ecol. 2004, 30, 2335–2358. [Google Scholar] [CrossRef]
- Phelps, T.J.; Palumbo, A.V.; Beliaev, A.S. Metabolomics and microarrays for improved understanding of phenotypic characteristics controlled by both genomics and environmental constraints. Curr. Opin. Biotechnol. 2002, 13, 20–24. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Hou, Y.; Zhou, X.; Chen, Q.; Guo, C.; Xia, Q.; Zhang, Y.; Zhao, P. Integrative proteomics and metabolomics analysis of insect larva brain: Novel insights into the molecular mechanism of insect wandering behavior. J. Proteome Res. 2016, 15, 193–204. [Google Scholar] [CrossRef]
- Bachmann, A.; Knust, E. The Use of P-Element Transposons to Generate Transgenic Flies. Methods Mol. Biol. 2008, 420, 61–77. [Google Scholar]
- Labbe, G.M.; Nimmo, D.D.; Alphey, L. piggybac- and PhiC31-mediated genetic transformation of the Asian tiger mosquito, Aedes albopictus (Skuse). PLoS Negl. Trop. Dis. 2010, 4, e788. [Google Scholar] [CrossRef] [Green Version]
- Pondeville, E.; Puchot, N.; Meredith, J.M.; Lynd, A.; Vernick, K.D.; Lycett, G.J.; Eggleston, P.; Bourgouin, C. Efficient ΦC31 integrase–mediated site-specific germline transformation of Anopheles gambiae. Nat. Protoc. 2014, 9, 1698–1712. [Google Scholar] [CrossRef]
- Chu, F.; Klobasa, W.; Wu, P.; Pinzi, S.; Grubbs, N.; Gorski, S.; Cardoza, Y.; Lorenzen, M.D. Germline transformation of the western corn rootworm, Diabrotica virgifera virgifera. Insect Mol. Biol. 2017, 26, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Quan, G.X.; Kanda, T.; Tamura, T. Induction of the white egg 3 mutant phenotype by injection of the double-stranded RNA of the silkworm white gene. Insect Mol. Biol. 2002, 11, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Adrianos, S.; Lorenzen, M.; Oppert, B. Metabolic pathway interruption: CRISPR/Cas9-mediated knockout of tryptophan 2,3-dioxygenase in Tribolium castaneum. J. Insect Physiol. 2018, 107, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Kistler, K.E.; Vosshall, L.B.; Matthews, B.J. Genome engineering with CRISPR-Cas9 in the mosquito Aedes aegypti. Cell Rep. 2015, 11, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Ma, D.; Huang, Y.; He, W.; Li, Y.; Vasseur, L.; You, M. Genome-wide investigation of transcription factors provides insights into transcriptional regulation in Plutella xylostella. Mol. Genet. Genomics 2018, 293, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Gupta, G.P.; Rajam, M.V. Silencing of acetylcholinesterase gene of Helicoverpa armigera by siRNA affects larval growth and its life cycle. J. Insect Physiol. 2009, 55, 273–278. [Google Scholar] [CrossRef]
- Fishilevich, E.; Vélez, A.M.; Storer, N.P.; Li, H.; Bowling, A.J.; Rangasamy, M.; Worden, S.E.; Narva, K.E.; Siegfried, B.D. RNAi as a management tool for the western corn rootworm, Diabrotica virgifera virgifera. Pest Manag. Sci. 2016, 72, 1652–1663. [Google Scholar] [CrossRef] [Green Version]
- Spit, J.; Philips, A.; Wynant, N.; Santos, D.; Plaetinck, G.; Broeck, J.V. Knockdown of nuclease activity in the gut enhances RNAi efficiency in the Colorado potato beetle, Leptinotarsa decemlineata, but not in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2017, 81, 103–116. [Google Scholar] [CrossRef]
- Camargo, R.A.; Barbosa, G.O.; Possignolo, I.P.; Peres, L.E.; Lam, E.; Lima, J.E.; Figueira, A.; Marques-Souza, H. RNA interference as a gene silencing tool to control Tuta absoluta in tomato (Solanum lycopersicum). PeerJ 2016, 4, e2673. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Reichelt, M.; Heckel, D.G. Functional analysis of the ABCs of eye color in Helicoverpa armigera with CRISPR/Cas9-induced mutations. Sci. Rep. 2017, 7, 40025. [Google Scholar] [CrossRef] [Green Version]
- Paglino, A.; Lombardo, F.; Arca, B.; Rizzi, M.; Rossi, F. Purification and biochemical characterization of a recombinant Anopheles gambiae tryptophan 2,3-dioxygenase expressed in Escherichia coli. Insect Biochem. Mol. Biol. 2008, 38, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Quan, G.X.; Kim, I.; Komoto, N.; Sezutsu, H.; Ote, M.; Shimada, T.; Kanda, T.; Mita, K.; Kobayashi, M.; Tamura, T. Characterization of the kynurenine 3-monooxygenase gene corresponding to the white egg 1 mutant in the silkworm Bombyx mori. Mol. Genet. Genom. 2002, 267, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Linzen, B. The Tryptophan → Ommochrome Pathway in Insects. In Advances in Insect Physiology; Treherne, J.E., Berridge, M.J., Wigglesworth, V.B., Eds.; Academic Press: Cambridge, MA, USA, 1974; pp. 117–246. [Google Scholar]
- Dustmann, J.H. Pigment studies on several eye-colour mutants of the honey bee, Apis mellifera. Nature 1968, 219, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Fridell, Y.-W.C.; Searles, L.L. Vermilion as a small selectable marker gene for Drosophila transformation. Nucleic Acids Res. 1991, 19, 5082. [Google Scholar] [CrossRef] [Green Version]
- Bottino-Rojas, V.; Ferreira-Almeida, I.; Nunes, R.D.; Feng, X.; Pham, T.B.; Kelsey, A.; Carballar-Lejarazu, R.; Gantz, V.; Oliveira, P.L.; James, A.A. Beyond the eye: Kynurenine pathway impairment causes midgut homeostasis dysfunction and survival and reproductive costs in blood-feeding mosquitoes. Insect Biochem. Mol. Biol. 2022, 142, 103720. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Howells, A.J.; Ryall, R.L. The ommochrome biosynthetic pathway in Drosophila melanogaster: The head particulate phenoxazinone synthase and the developmental onset of xanthommatin synthesis. Biochem. Genet. 1976, 14, 1077. [Google Scholar] [CrossRef]
- Klemenz, R.; Weber, U.; Gehring, W.J. The white gene as a marker in a new P-element vector for gene transfer in Drosophila. Nucleic Acids Res. 1987, 15, 3947–3959. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.B.; Nicholas, H.B.J.; Deerfield, D.W.I. GeneDoc: Analysis and visualization of genetic variation. Embnew News 1997, 4, 1–6. [Google Scholar]
- Corthell, J.T. DNA and RNA extraction protocols, In Basic Molecular Protocols in Neuroscience: Tips, Tricks, and Pitfalls; Academic Press: Cambridge, MA, USA, 2014; pp. 11–19. [Google Scholar]
- Stratakis, E.; Schartau, W. Tryptophan 2,3-dioxygenase in the development of the stick insect, Carausius morosus Br. J. Comp. Physiol. B 1979, 132, 351–355. [Google Scholar] [CrossRef]
- Handler, A.M. A current perspective on insect gene transformation. Insect Biochem. Mol. Biol. 2001, 31, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Schmid, B.G.M.; Pogoda, F.S.; Wimmer, E.A. Fluorescent transformation markers for insect transgenesis. Insect Biochem. Mol. Biol. 2002, 32, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Besansky, N.J.; Fahey, G.T. Utility of white gene in estimating phylogenetic relationships among mosquitoes (Diptera: Culicidae). Moleciular Biol. Evol. 1997, 14, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, M.D.; Brown, S.J.; Denell, R.E.; Beeman, R.W. Cloning and characterization of the Tribolium castaneum eye-color genes encoding tryptophan oxygenase and kynurenine 3-monooxygenase. Genetics 2002, 160, 225–234. [Google Scholar] [CrossRef]
- Sullivan, D.T.; Bell, L.A.; Paton, D.R.; Sullivan, M.C. Purine transport by malpighian tubules of pteridine-deficient eye color mutants of Drosophila melanogaster. Biochem. Genet. 1979, 17, 565–573. [Google Scholar] [CrossRef]
- Souza, D.; Christensen, S.A.; Wu, K.; Buss, L.; Kleckner, K.; Darrisaw, C.; Shirk, P.D.; Siegfried, B.D. RNAi-induced knockdown of white gene in the southern green stink bug (Nezara viridula L.). Sci. Rep. 2022, 12, 10396. [Google Scholar] [CrossRef]
- Xu, P.; Ze, L.J.; Kang, W.N.; Wu, J.J.; Jin, L.; Anjum, A.A.; Li, G.Q. Functional divergence of white genes in Henosepilachna vigintioctopunctata revealed by RNA interference. Insect Mol. Biol. 2020, 29, 466–476. [Google Scholar]
- Osanai-Futahashi, M.; Tatematsu, K.I.; Yamamoto, K.; Narukawa, J.; Uchino, K.; Kayukawa, T.; Shinoda, T.; Banno, Y.; Tamura, T.; Sezutsu, H. Identification of the Bombyx red egg gene reveals involvement of a novel transporter family gene in late steps of the insect ommochrome biosynthesis pathway. J. Biol. Chem. 2012, 287, 17706–17714. [Google Scholar] [CrossRef] [Green Version]
- Beard, C.B.; Benedict, M.Q.; Primus, J.P.; Finnerty, V.; Collins, F.H. Eye pigments in wild-type and eye-color mutant strains of the African malaria vector Anopheles gambiae. J. Hered. 1995, 86, 375–380. [Google Scholar] [CrossRef]
- Grubbs, N.; Haas, S.; Beeman, R.W.; Lorenzen, M.D. The ABCs of eye color in Tribolium castaneum: Orthologs of the Drosophila white, scarlet, and brown Genes. Genetics 2015, 199, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.E.; Bradish, H.M.; Gatehouse, J.A.; Fitches, E.C. Systemic RNAi in the small hive beetle Aethina tumida Murray (Coleoptera: Nitidulidae), a serious pest of the European honey bee Apis mellifera. Pest Manage. Sci. 2017, 73, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira-Pinto, C.E.; Coelho, R.R.; Leite, A.G.B.; Silveira, D.A.; de Souza, D.A.; Lopes, R.B.; Macedo, L.L.P.; Silva, M.C.M.; Ribeiro, T.P.; Morgante, C.V.; et al. Increasing Anthonomus grandis susceptibility to Metarhizium anisopliae through RNAi-induced AgraRelish knockdown: A perspective to combine biocontrol and biotechnology. Pest Manag. Sci. 2021, 77, 4054–4063. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Name | Sequence (5′-3′) |
|---|---|---|
| Primers for dsRNA synthesis | TDO-F | TAATACGACTCACTATAGGGAGATGGGGGAAATACCAACGAGC |
| TDO-R | TAATACGACTCACTATAGGGAGACTTCTCATGGCGGAGGTGAG | |
| GFP-F | TAATACGACTCACTATAGGGATGAGTAAAGGAGAAGAACT | |
| GFP-R | TAATACGACTCACTATAGGGTTTGTATAGTTCATCCATGC | |
| Primers for RT-qPCR | Actin-F | AGCCGGTTTCGCCGGTGATGAC |
| Actin-R | CACTTCATGATGGAGTTGTAGAC | |
| TDO-qPCR-F | TCAGCGATGGTTGGAGAGGA | |
| TDO-qPCR-R | TCTCCGTCTTTTCGTCGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, L.-J.; Weng, X.-Q.; Weng, M.-Q.; Guo, Y.-J.; Carballar-Lejarazú, R.; Zhang, F.-P.; Wu, S.-Q. Function of Tryptophan 2,3-Dioxygenase in Monochamus alternatus Hope Revealed by RNA Interference. Forests 2023, 14, 215. https://doi.org/10.3390/f14020215
Sheng L-J, Weng X-Q, Weng M-Q, Guo Y-J, Carballar-Lejarazú R, Zhang F-P, Wu S-Q. Function of Tryptophan 2,3-Dioxygenase in Monochamus alternatus Hope Revealed by RNA Interference. Forests. 2023; 14(2):215. https://doi.org/10.3390/f14020215
Chicago/Turabian StyleSheng, Liang-Jing, Xiao-Qian Weng, Ming-Qing Weng, Ya-Jie Guo, Rebeca Carballar-Lejarazú, Fei-Ping Zhang, and Song-Qing Wu. 2023. "Function of Tryptophan 2,3-Dioxygenase in Monochamus alternatus Hope Revealed by RNA Interference" Forests 14, no. 2: 215. https://doi.org/10.3390/f14020215