
The Effects of Stand Density Control on Carbon Cycle in Chamaecyparis obtusa (Siebold and Zucc.) Endl. Forests

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Study Methods
2.2.1. Light Intensity
2.2.2. Aboveground Biomass of Understory Vegetation
2.2.3. Tree Carbon Storage and Annual Net Primary Production
2.2.4. Litterfall
2.2.5. Forest Floor and Soil C Concentration and Storage
2.2.6. Litter Decomposition
2.2.7. Soil Respiration
2.2.8. Net Ecosystem Production
2.2.9. Statistical Analysis
3. Results and Discussion
3.1. Changes in C Storage by C Pool Resulting from Different Thinning Intensities in Cypress Forests
3.1.1. Aboveground C Storage in Trees
3.1.2. Forest Floor and Soil C Storage
3.2. Changes in C Shift by C Pool Resulting from Different Thinning Intensities in Cypress Forests
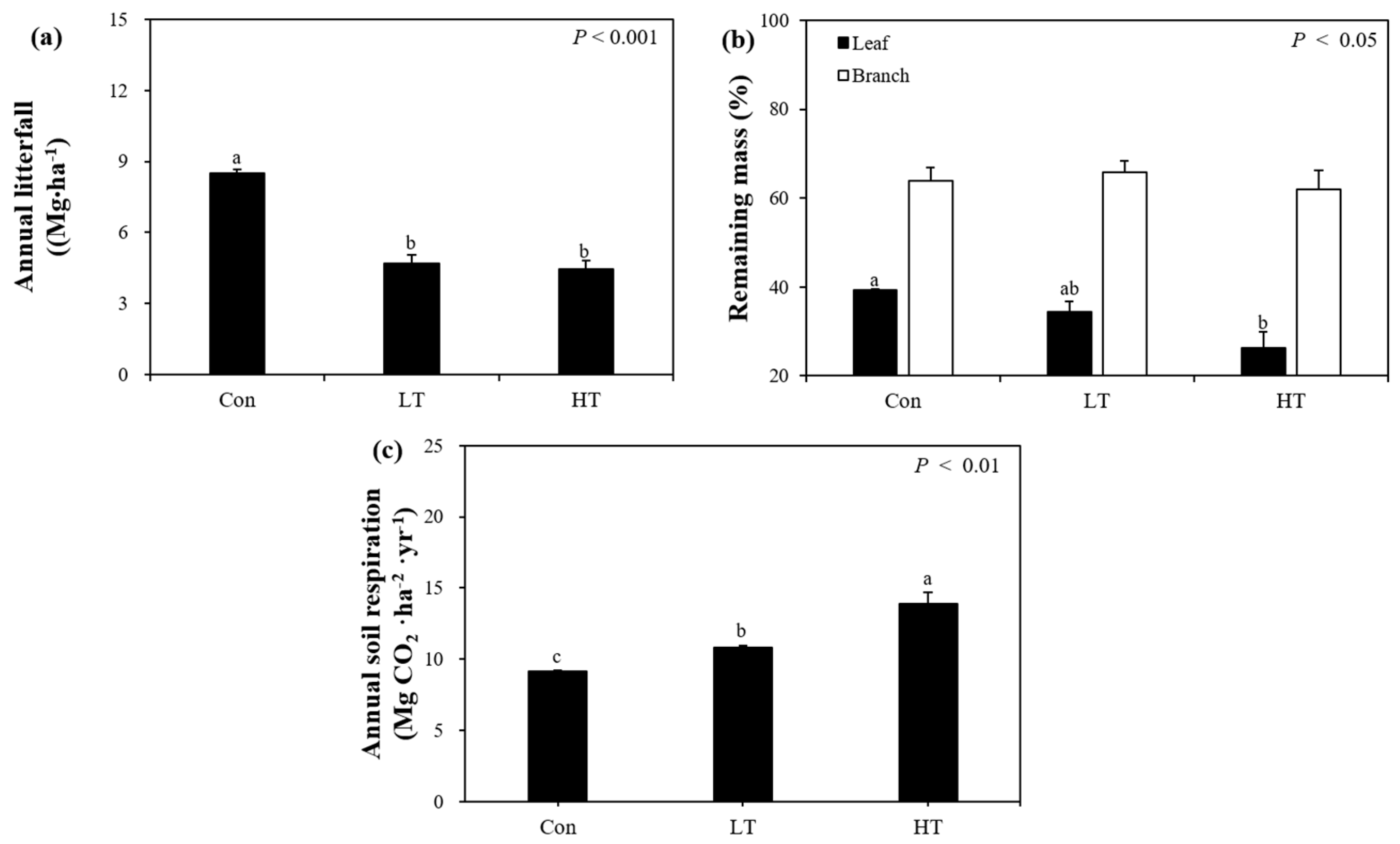
3.2.1. Litterfall
3.2.2. Litter Decomposition
3.2.3. Soil Respiration
3.3. C cycle by Thinning Intensity of Cypress Forest
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tans, P.P.; Fung, I.Y.; Takahashi, T. Observational contrains on the global atmospheric CO2 budget. Science 1990, 247, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Candell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, B.W.; Kolb, T.E.; Hart, S.C.; Kaye, J.P.; Dore, S.; Montes-Helu, M. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA ponderosa pine forests. For. Ecol. Manag. 2008, 255, 4047–4055. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Good Practice Guidance for Land Use, Land-Use Change and Forestry; IPCC: Hayama, Japan, 2003; ISBN 4887880030. [Google Scholar]
- Poeplau, C.; Don, A.; Vesterdal, L.; Leifeld, J.; Van Wesemael, B.A.S.; Schumacher, J.; Gensior, A. Temporal dynamics of soil organic carbon after land-use change in the temperate zone–carbon response functions as a model approach. Glob. Chang. Biol. 2011, 17, 2415–2427. [Google Scholar] [CrossRef]
- North, M.; Hurteau, M.; Innes, J. Fire suppression and fuels treatment effects on mixed-conifer carbon stocks and emissions. Ecol. Appl. 2009, 19, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Burton, L.D. Introduction to Forestry Science; Cengage Learning: Boston, MA, USA, 2012; ISBN 9781133711629. [Google Scholar]
- Nishizono, T. Effects of thinning level and site productivity on age-related changes in stand volume growth can be explained by a single rescaled growth curve. For. Ecol. Manag. 2010, 259, 2276–2291. [Google Scholar] [CrossRef]
- Lee, J.; Han, S.H.; Kim, S.; Lee, S.; Son, Y.M.; Son, Y. A meta-analysis on the effect of forest thinning on diameter growth and carbon stocks in Korea. J. Korean Soc. For. Sci. 2015, 104, 527–535. [Google Scholar] [CrossRef]
- Nilsen, P.; Strand, L.T. Thinning intensity effects on carbon and nitrogen stores and fluxes in a Norway spruce (Picea abies (L.) Karst.) stand after 33 years. For. Ecol. Manag. 2008, 256, 201–208. [Google Scholar] [CrossRef]
- Jonard, M.; Misson, L.; Ponette, Q. Long-term thinning effects on the forest floor and the foliar nutrient status of Norway spruce stands in the Belgian Ardennes. Can. J. For. Res. 2006, 36, 2684–2695. [Google Scholar] [CrossRef]
- Prescott, C.E. Effects of clearcutting and alternative silvicultural systems on rates of decomposition and nitrogen mineralization in a coastal montane coniferous forest. For. Ecol. Manag. 1097, 95, 253–260. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Guan, D.; Li, W.; Sun, D.; Jin, C.; Yuan, F.; Wang, A.; Wu, J. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. For. Ecol. Manag. 2018, 429, 36–43. [Google Scholar] [CrossRef]
- Kim, S.; Kim, C.; Han, S.H.; Lee, S.T.; Son, Y. A multi-site approach toward assessing the effect of thinning on soil carbon contents across temperate pine, oak, and larch forests. For. Ecol. Manag. 2018, 424, 62–70. [Google Scholar] [CrossRef]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korean, 2020; pp. 236–237. (In Korean)
- Korea Forest Service. Detailed Implementation Plan for Major Tasks in 2022; Korea Forest Service: Daejeon, Korean, 2022; p. 177. (In Korean)
- Kim, C.; Son, Y.; Lee, W.K.; Jeong, J.; Noh, N.J. Influences of forest tending works on carbon distribution and cycling in a Pinus densiflora S. et Z. stand in Korea. For. Ecol. Manag. 2009, 257, 1420–1426. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S.O.; Do, K.B.; Ji, W.D.; Kim, S.G.; Back, Y.D.; Kim, K.J. Analysis of the component and immunological efficacy of Chamaecyparis obtusa leaf extract. Korean J. Clin. Lab. Sci. 2018, 50, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Park, S.G.; Kang, H.M. Characteristics of vegetation structure in Chamaecyparis obtusa stands. Korean J. Environ. Ecol. 2015, 29, 907–916. [Google Scholar] [CrossRef]
- Kim, C.; Baek, G.; Choi, B.; Ha, J.; Bae, E.J.; Lee, K.S.; Son, Y.M. Carbon stocks of tree, forest floor, and mineral soil in Cryptomeria japonica and Chamaecyparis obtusa stands. J. Korean Soc. For. Sci. 2020, 109, 169–178. [Google Scholar]
- Park, S.G.; You, H.C.; Oh, C.J.; Choi, W.K. Analysis of the correlation between site environmental factors and tree ring growth in Chamaecyparis obtusa stands in Jeonnam Province. Korean J. Environ. Ecol. 2015, 29, 777–784. [Google Scholar] [CrossRef]
- Park, J.H.; Son, Y.M.; Lee, K.S.; Park, E.J.; Jung, S.Y. The Adequacy Assessment and Growth Effects by Methods of 1st Thinning in Chamaecyparis obtusa Forest. J. Korean Soc. For. Sci. 2019, 108, 349–356. [Google Scholar]
- Ko, S.; Yoon, T.K.; Kim, S.; Kim, C.; Lee, S.T.; Seo, K.W.; Son, Y. Thinning intensity effects on carbon storage of soil, forest floor and coarse woody debris in Pinus densiflora stands. J. Korean Soc. For. Sci. 2014, 103, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Song, S.-J.; Jang, K.-S.; Hwang, I.-C.; An, K.-W.; Lee, K.-H. Effects of Tree Density Control on Carbon Dynamics in Young Pinus densiflora stands. J. Korean For. Soc. 2016, 105, 275–283. [Google Scholar]
- Korea Soil Information System. Soil Classification Map. Available online: www.soil.rda.go.kr (accessed on 17 October 2022). (In Korean)
- Korea Meteorological Association (KMA). Climatological Normals of Korea. Available online: www.kma.go.kr (accessed on 29 September 2022). (In Korean)
- Korea Forest Research Institute (KFRI). Carbon Emission Factors and Biomass Allometric Equations by Species in Korea; KFRI: Seoul, Korea, 2014; pp. 58–59. (In Korean) [Google Scholar]
- Korea Forest Research Institute. Main Tree Species Carbon Emission Factors for Forest Greenhouse Inventory; KFRI: Seoul, Korea, 2010; p. 89. (In Korean) [Google Scholar]
- Wider, R.K.; Lang, G.E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Kim, H.J.; Han, S.H.; Kim, S.; Chang, H.; Son, Y. Effects of Artificial CO2 Release in Soil on Chlorophyll Content and Growth of Pinus densiflora and Quercus variabilis Seedlings. J. Korean Soc. For. Sci. 2018, 107, 351–360. [Google Scholar]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Koo, J.W.; Son, Y.; Kim, R.H.; Kim, J. A study on methods of separating soil respiration by source. Korean J. Agric. For. Meteorol. 2005, 7, 28–34. [Google Scholar]
- Lee, M.S.; Nakane, K.; Nakatsubo, T.; Koizumi, H. Seasonal Changes in the Contribution of Root Respiration to Total Soil Respiration in a Cool-Temperate Deciduous Forest. In Roots: The Dynamic Interface between Plants and the Earth; Abe, J., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 311–318. ISBN 9789401729239. [Google Scholar]
- Lee, S.T.; Son, Y.M.; Lee, K.J.; Hwang, J.; Choi, J.C.; Shin, H.C.; Park, N.C. Aboveground Carbon Storage of Quercus acuta Stands by Thinning Intensity. Korean J. Agric. For. Meteorol. 2005, 7, 282–288. [Google Scholar]
- Hwang, J.H.; Bae, S.W.; Lee, K.J.; Lee, K.S.; Kim, H.S. Short-term Effect of Thinning on Aboveground Carbon Storage in Korean Pine (Pinus koraiensis) Plantation. J. Korean Soc. For. Sci. 2008, 97, 605–610. [Google Scholar]
- Hou, L.; Li, Z.; Luo, C.; Bai, L.; Dong, N. Optimization forest thinning measures for carbon budget in a mixed pine-oak stand of the Qingling Mountains, China: A Case study. Forests 2016, 7, 272. [Google Scholar] [CrossRef]
- Choi, J.K.; Lee, B.K.; Lee, D.S.; Choi, I.H. Growth Monitoring of Korean White Pine (Pinus koraiensis) Plantation by Thinning Intensity. J. Korean For. Soc. 2014, 103, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Bae, E.J.; Son, Y.M.; Jeon, H.G.; Lee, K.S.; Jung, S.Y. Increment Change Monitoring of DBH and Stand Volume Based on Thinning Treatments in Quercus glauca Stands. J. Agric. Life Sci. 2021, 55, 71–75. [Google Scholar] [CrossRef]
- Lee, D.; Jung, S.; Choi, J. Analysis of Tree Growth Characteristics by First and Second Thinning in Korean White Pine Plantations. J. Korean Soc. For. Sci. 2022, 111, 150–164. [Google Scholar] [CrossRef]
- Zhou, D.; Zhao, S.Q.; Liu, S.; Oeding, J. A meta-analysis on the impacts of partial cutting on forest structure and carbon storage. Biogeosciences 2013, 10, 3691–3703. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Son, Y.; Lee, W.K.; Yang, A.R.; Noh, N.J.; Byun, J.G. Influence of thinning on carbon storage in a Japanese larch (Larix kaempferi) plantation in Yangpyeong, central Korea. For. Sci. Technol. 2010, 6, 35–40. [Google Scholar] [CrossRef]
- Inagaki, Y.; Kuramoto, S.; Torii, A.; Shinomiya, Y.; Fukata, H. Effects of thinning on leaf-fall and leaf-litter nitrogen concentration in hinoki cypress (Chamaecyparis obtusa Endlicher) plantation stands in Japan. For. Ecol. Manag. 2008, 255, 1859–1867. [Google Scholar] [CrossRef]
- Achat, D.L.; Fortin, M.; Landmann, G.; Ringeval, B.; Augusto, L. Forest soil carbon is threatened by intensive biomass harvesting. Sci. Rep. 2015, 5, 15991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slodicak, M.; Novak, J.; Skovsgaard, J.P. Wood production, litter fall and humus accumulation in a Czech thinning experiment in Norway spruce (Picea abies (L.) Karst.). For. Ecol. Manag. 2005, 209, 157–166. [Google Scholar] [CrossRef]
- Novák, J.; Slodičák, M.; Dušek, D.; Kacálek, D. Norway spruce litterfall and forest floor in the IUFRO thinning experiment CZ 13-Vítkov. J. For. Sci. 2013, 59, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Korea Forest Research Institute. The 5th National Forest Inventory Report; Korea Forest Research Institute: Seoul, Korea, 2011; p. 87. (In Korean) [Google Scholar]
- Hennessey, T.C.; Dougherty, P.M.; Cregg, B.M.; Wittwer, R.F. Annual variation in needle fall of a loblolly pine stand in relation to climate and stand density. For. Ecol. Manag. 1992, 51, 329–338. [Google Scholar] [CrossRef]
- Berg, B.; Meentemeyer, V. Litter fall in some European coniferous forests as dependent on climate: A synthesis. Can. J. For. Res. 2001, 31, 292–301. [Google Scholar] [CrossRef]
- Trettin, C.C.; Jurgensen, M.F.; Gale, M.R.; McLaughlin, J.W. Soil Carbon in Northern Forested Wetlands: Impacts of Silvicultural Practices. In Carbon Forms and Functions in Forest Soils; McFee, W.W., Kelly, J.M., Eds.; 1995; pp. 437–461. ISBN 9780891188186. [Google Scholar]
- Kunhamu, T.K.; Kumar, B.M.; Viswanath, S. Does thinning affect litterfall, litter decomposition, and associated nutrient release in Acacia mangium stands of Kerala in peninsular India? Can. J. For. Res. 2009, 39, 792–801. [Google Scholar] [CrossRef]
- Lim, H.; Choi, W.J.; Ahn, K.; Lee, K.H. Ecosystem respiration and tree growth influenced by thinning in a red pine forest in southern Korea. For. Sci. Technol. 2012, 8, 192–204. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter production in forests of the world. In Advances in Ecological Research; Cragg, J.B., Ed.; Academic Press: Cambridge, MA, USA, 1964; Volume 2, pp. 101–157. ISBN 9780120139026. [Google Scholar]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Tang, J.; Qi, Y.; Xu, M.; Misson, L.; Goldstein, A.H. Forest thinning and soil respiration in a ponderosa pine plantation in the Sierra Nevada. Tree Physiol. 2005, 25, 57–66. [Google Scholar] [CrossRef]
- Steele, S.J.; Gower, S.T.; Vogel, J.G.; Norman, J.M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiol. 1997, 17, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Han, H.; Kang, F.; Liu, K.; Song, Y.; Zhou, B.; Li, Y. Short-term effects of thinning on soil respiration in a pine (Pinus tabulaeformis) plantation. Biol. Fertil. Soils 2014, 50, 357–367. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Chen, D.; Huang, G.; Zhou, L.; Fu, S. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biol. Biochem. 2011, 43, 2355–2357. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; Montero, G.; Del Río, M. Carbon stocks in a Scots pine afforestation under different thinning intensities management. Mitig. Adapt. Strateg. Glob. Chang. 2016, 21, 1059–1072. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Year (Age) | Con | LT | HT |
|---|---|---|---|---|
| Thinning intensity (%) | 0% | 30% | 50% | |
| Light intensity (μmol·m2·s−1) | 205 ± 80 c | 355 ± 34 b | 649 ± 43 a | |
| Trees per ha (N ha−1) | 2000 (27) | 2900 ± 43 | 1858 ± 134 | 1458 ± 42 |
| 2018 (45) | 2338 ± 22 | 1792 ± 33 | 1308 ± 22 | |
| Minimum Diameter at breast height (DBH) (cm) | 2000 (27) | 9.10 ± 0.3 a | 9.93 ± 0.96 a | 10.60 ± 0.31 a |
| 2018 (45) | 11.85 ± 0.85 a | 12.67 ± 0.37 ab | 13.97 ± 0.29 b | |
| Maximum DBH (cm) | 2000 (27) | 20.05 ± 0.65 a | 21.00 ± 0.15 a | 20.33 ± 0.44 a |
| 2018 (45) | 28.15 ± 0.75 a | 30.17 ± 0.28 ab | 32.47 ± 0.95 b | |
| Mean DBH (cm) | 2000 (27) | 13.75 ± 0.16 b | 14.99 ± 0.27 a | 15.60 ± 0.49 a |
| 2018 (45) | 18.65 ± 0.28 c | 20.42 ± 0.14 b | 23.11 ± 0.45 a | |
| Aboveground biomass of understory vegetation (Mg·ha−1) | 0.02 ± 0.002 c | 0.06 ± 0.01 b | 0.43 ± 0.02 a |
| Con | LT | HT | ||
|---|---|---|---|---|
| Mean annual increment (MAI) (cm·year−1) | 0.27 ± 0.04 b | 0.30 ± 0.02 b | 0.42 ± 0.01 a | |
| Carbon storage (Mg·C·ha−1) | 2000 | 120.10 ± 1.22 a | 90.25 ± 3.98 b | 77.02 ± 5.33 b |
| 2018 | 179.43 ± 3.88 a | 163.86 ± 3.79 ab | 151.78 ± 8.78 b | |
| Net primary production (NPP) (Mg·C·ha−1·year−1) | 3.30 ± 0.28 b | 4.09 ± 0.20 a | 4.15 ± 0.26 a | |
| Carbon Storage (Mg·C·ha−1) | Con | LT | HT |
|---|---|---|---|
| Forest floor | 7.86 ± 0.66 a | 6.26 ± 0.01 b | 3.83 ± 0.32 c |
| Soil | 93.9 ± 7.84 | 90.3 ± 2.11 | 86.5 ± 4.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-G.; Lee, D.-H.; Jung, J.-Y.; Lee, S.-G.; Han, S.H.; Kim, S.; Kim, H.-J. The Effects of Stand Density Control on Carbon Cycle in Chamaecyparis obtusa (Siebold and Zucc.) Endl. Forests. Forests 2023, 14, 217. https://doi.org/10.3390/f14020217
Lee J-G, Lee D-H, Jung J-Y, Lee S-G, Han SH, Kim S, Kim H-J. The Effects of Stand Density Control on Carbon Cycle in Chamaecyparis obtusa (Siebold and Zucc.) Endl. Forests. Forests. 2023; 14(2):217. https://doi.org/10.3390/f14020217
Chicago/Turabian StyleLee, Jeong-Gwan, Du-Hee Lee, Jun-Young Jung, Sle-Gee Lee, Seung Hyun Han, Seongjun Kim, and Hyun-Jun Kim. 2023. "The Effects of Stand Density Control on Carbon Cycle in Chamaecyparis obtusa (Siebold and Zucc.) Endl. Forests" Forests 14, no. 2: 217. https://doi.org/10.3390/f14020217
APA StyleLee, J.-G., Lee, D.-H., Jung, J.-Y., Lee, S.-G., Han, S. H., Kim, S., & Kim, H.-J. (2023). The Effects of Stand Density Control on Carbon Cycle in Chamaecyparis obtusa (Siebold and Zucc.) Endl. Forests. Forests, 14(2), 217. https://doi.org/10.3390/f14020217