
Analysis of the Cell Structural Characters of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) and Its Varieties

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Detection of Cell Structure Properties
2.2.1. Determination of Vascular Bundle Sizes
2.2.2. Determination of the Length and Width of Fiber Cells and Parenchyma Cells
2.2.3. Determination of Wall Thickness and Lumen Diameters of Fiber Cells and Parenchyma Cells
2.3. Data Processing and Analysis
3. Results
3.1. Genetic Variation of The Vascular Bundle-Related Indexes
Vascular Bundle Size
3.2. Genetic Variation of Fiber Cell Morphology
3.2.1. Fiber Cell Length and Width
3.2.2. Fiber Cell Wall Thickness and Lumen Diameter
3.3. Genetic Variation of Parenchyma Cell Morphology
3.3.1. Parenchyma Cell Length and Width
3.3.2. Parenchyma Cell Wall Thickness and Lumen Diameter
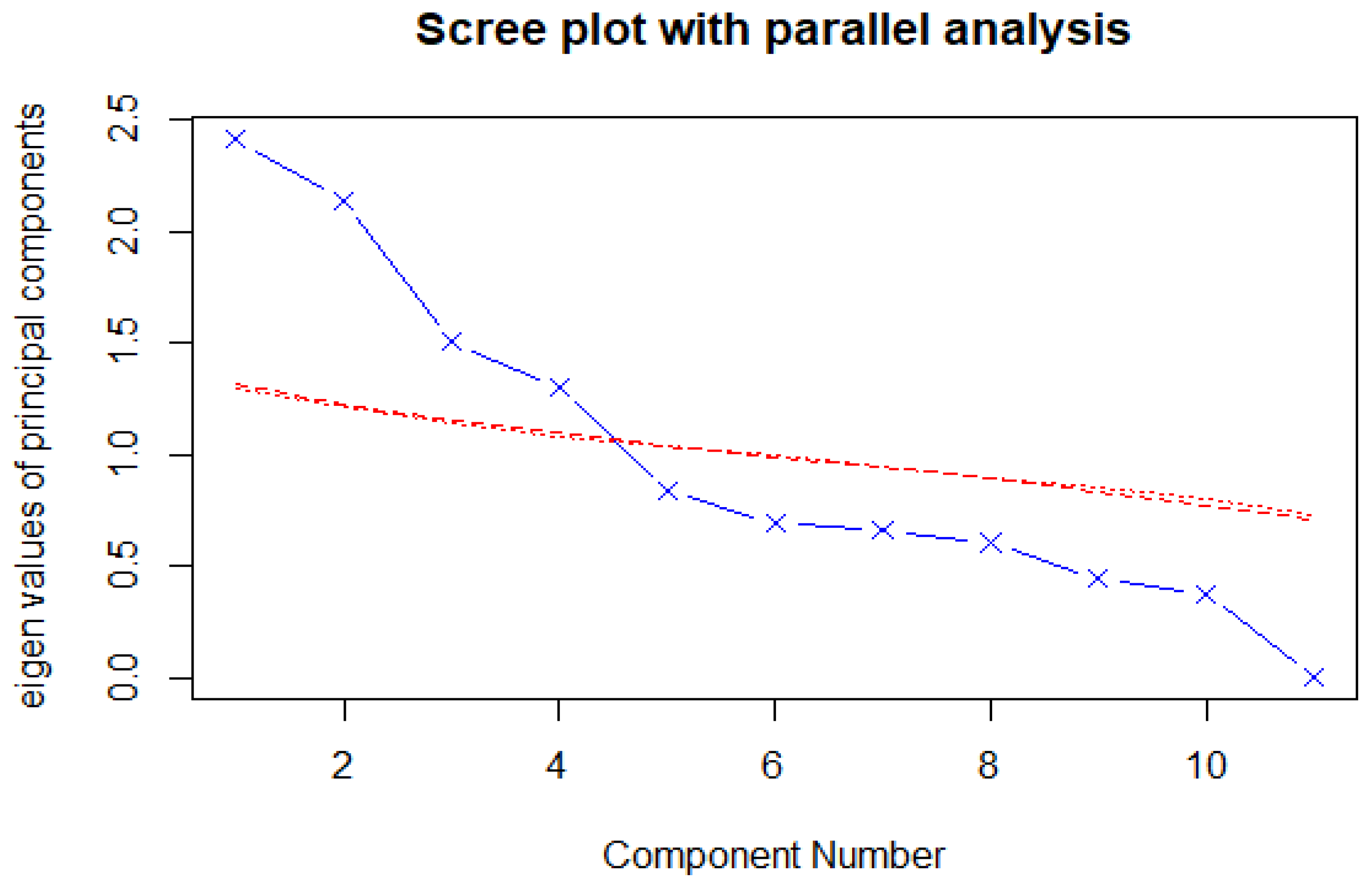
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fei, B.; Gao, Z.; Wang, J.; Liu, Z. Chapter 14—Biological, Anatomical, and Chemical Characteristics of Bamboo. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 283–306. [Google Scholar]
- Wu, C.J.; Zhang, J.C.; Yu, D.M.; Li, R.G. Dissolving pulp from bamboo-willow. Cellulose 2018, 25, 777–785. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, H.K.; Lu, F.; Tian, G.L.; Lin, J.G. Bamboo fibers for composite applications: A mechanical and morphological investigation. J. Mater. Sci. 2014, 49, 2559–2566. [Google Scholar] [CrossRef]
- Cao, X.; Li, F.; Zheng, T.; Li, G.; Wang, W.; Li, Y.; Chen, S.; Li, X.; Lu, Y. Cellulose-Based functional hydrogels derived from bamboo for product design. Front Plant Sci 2022, 13, 958066. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, J.E.G.; Elbersen, H.W.; Daza Montaño, C.M. 6—Bamboo Production for Industrial Utilization. In Perennial Grasses for Bioenergy and Bioproducts; Alexopoulou, E., Ed.; Academic Press: Boston, MA, USA, 2018; pp. 175–216. [Google Scholar]
- Sugesty, S.; Kardiansyah, T.; Hardiani, H. Bamboo as Raw Materials for Dissolving Pulp with Environmental Friendly Technology for Rayon Fiber. Procedia Chem. 2015, 17, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Gratani, L.; Crescente, M.F.; Varone, L.; Fabrini, G.; Digiulio, E. Growth pattern and photosynthetic activity of different bamboo species growing in the Botanical Garden of Rome. Flora—Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 77–84. [Google Scholar] [CrossRef]
- Huang, Y.; Fei, B.; Wei, P.; Zhao, C. Mechanical properties of bamboo fiber cell walls during the culm development by nanoindentation. Ind. Crops Prod. 2016, 92, 102–108. [Google Scholar] [CrossRef]
- Wahab, R.; Mohamed, A.; Mustafa, M.T.; Hassan, A. Physical Characteristics and Anatomical Properties of Cultivated Bamboo (Bambusa vulgaris Schrad.) Culms. J. Biol. Sci. 2009, 9, 753–759. [Google Scholar] [CrossRef]
- Parameswaran, N.; Liese, W. On the fine structure of bamboo fibres. Wood Sci. Technol. 1976, 10, 231–246. [Google Scholar] [CrossRef]
- Liese, W. Anatomy and Properties of Bamboo; Bamboos; Geselschaft fiir Technische Zusanimenarbeit: Deutsche, Germany, 1985. [Google Scholar]
- Akinlabi, E.T.; Anane-Fenin, K.; Akwada, D.R. Properties of Bamboo. In Bamboo: The Multipurpose Plant; Springer International Publishing: Cham, Switzerland, 2017; pp. 87–147. [Google Scholar]
- Wen, Y.; Xu, K.K.; Tang, J.; Li, Y.S. Research Status and Development Trend of Steel-Bamboo Composite Structure. Adv. Mater. Res. 2014, 893, 716–719. [Google Scholar]
- Xie, J.; Qi, J.; Hu, T.; Xiao, H.; Chen, Y.; De Hoop, C.F.; Huang, X. Anatomical characteristics and physical–mechanical properties of Neosinocalamus affinis from Southwest China. Eur. J. Wood Wood Prod. 2017, 75, 659–662. [Google Scholar] [CrossRef]
- Kanzawa, E.; Aoyagi, S.; Nakano, T. Vascular bundle shape in cross-section and relaxation properties of Moso bamboo (Phyllostachys pubescens). Mater. Sci. Eng. C 2011, 31, 1050–1054. [Google Scholar] [CrossRef]
- Mohmod, A.L.; Tarmeze, W.; Ahmad, F. Anatomical features and mechanical properties of three Malaysian bamboos. J. Trop. For. Sci. 1990, 2, 227–234. [Google Scholar]
- Chen, Y.; Das, R.; Battley, M. Effects of cell size and cell wall thickness variations on the strength of closed-cell foams. Int. J. Eng. Sci. 2017, 120, 220–240. [Google Scholar] [CrossRef]
- Zhang, D.; He, M.; Qin, S.; Yu, J. Effect of fiber length and dispersion on properties of long glass fiber reinforced thermoplastic composites based on poly(butylene terephthalate). RSC Adv. 2017, 7, 15439–15454. [Google Scholar] [CrossRef] [Green Version]
- Wangaard, F.G.; Woodson, G.E. Fiber length strength interrelationship for slash pine and its effect on pulp-sheet properties. Wood Sci. 1973, 5, 235–240. [Google Scholar]
- Donaldson, L.A.; Nanayakkara, B.; Radotić, K.; Djikanovic-Golubović, D.; Mitrović, A.; Bogdanović Pristov, J.; Simonović Radosavljević, J.; Kalauzi, A. Xylem parenchyma cell walls lack a gravitropic response in conifer compression wood. Planta 2015, 242, 1413–1424. [Google Scholar] [CrossRef]
- Zheng, H.; Cai, M.; Bai, Y.; Xu, J.; Xie, Y.; Song, H.; Li, J.; Gao, J. The Effect of Guttation on the Growth of Bamboo Shoots. Forests 2021, 13, 31. [Google Scholar] [CrossRef]
- Vorontsova, M.S.; Clark, L.G.; Dransfield, J.; Govaerts, R.; Baker, W. World Checklist of Bamboos and Rattans; International Network of Bamboo and Rattan; Kew: London, UK, 2016. [Google Scholar]
- Scurlock, J.M.O.; Dayton, D.C.; Hames, B. Bamboo: An overlooked biomass resource? Biomass Bioenergy 2000, 19, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Lu, Y.; Li, L.; Zhao, Q.; Feng, Q.; Gao, Z.; Lu, H.; Hu, T.; Yao, N.; Liu, K.; et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla). Nat. Genet. 2013, 45, 456–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Zhang, C.; Zhang, Y.; Hu, T.; Mu, S.; Li, X.; Gao, J. Transcriptome sequencing and analysis of the fast growing shoots of moso bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e78944. [Google Scholar] [CrossRef] [PubMed]
- Yeasmin, L.; Ali, M.N.; Gantait, S.; Chakraborty, S. Bamboo: An overview on its genetic diversity and characterization. 3 Biotech 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.H.; Jo, H.G.; Yang, J.H.; Ki, S.H.; Shin, H.J. Antioxidative and Anti-Melanogenic Activities of Bamboo Stems (Phyllostachys nigra variety henosis) via PKA/CREB-Mediated MITF Downregulation in B16F10 Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 409. [Google Scholar] [CrossRef] [Green Version]
- Nirmala, C.; Bisht, M.S.; Bajwa, H.K.; Santosh, O. Bamboo: A rich source of natural antioxidants and its applications in the food and pharmaceutical industry. Trends Food Sci. Technol. 2018, 77, 91–99. [Google Scholar] [CrossRef]
- Yang, J.H.; Choi, M.H.; Na, C.S.; Cho, S.S.; Kim, J.H.; Ku, S.K.; Cho, I.J.; Shin, H.J.; Ki, S.H. Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury. Nutrients 2019, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Hartono, R.; Iswanto, A.H.; Priadi, T.; Herawati, E.; Farizky, F.; Sutiawan, J.; Sumardi, I. Physical, Chemical, and Mechanical Properties of Six Bamboo from Sumatera Island Indonesia and Its Potential Applications for Composite Materials. Polymers 2022, 14, 4868. [Google Scholar] [CrossRef]
- Hemingway, H.W.; Burrows, A.M.; Omstead, K.M.; Zohdy, S.; Pastor, J.F.; Muchlinski, M.N. Vertical Clinging and Leaping Ahead: How Bamboo Has Shaped the Anatomy and Physiology of Hapalemur. Anat. Rec. 2020, 303, 295–307. [Google Scholar] [CrossRef]
- Luo, J.; Lian, C.; Liu, R.; Zhang, S.; Yang, F.; Fei, B. Comparison of metaxylem vessels and pits in four sympodial bamboo species. Sci. Rep. 2019, 9, 10876. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.U.; Konnerth, J.; Ramage, M.H.; Gusenbauer, C. Mapping thermal conductivity across bamboo cell walls with scanning thermal microscopy. Sci. Rep. 2019, 9, 16667. [Google Scholar] [CrossRef]
- Shao, Z.; Wang, F. Mechanical Characteristics of Bamboo Structure and Its Components. In The Fracture Mechanics of Plant Materials: Wood and Bamboo; Springer: Singapore, 2018; pp. 125–146. [Google Scholar]
- Wang, S.; Zhan, H.; Li, P.; Chu, C.; Li, J.; Wang, C. Physiological Mechanism of Internode Bending Growth After the Excision of Shoot Sheath in Fargesia yunnanensis and Its Implications for Understanding the Rapid Growth of Bamboos. Front. Plant Sci. 2020, 11, 418. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qiao, G.; Xu, J.; Jin, K.; Fan, M.; Ding, Y.; Wei, Q.; Zhuo, R. Anatomical Characteristics and Variation Mechanisms on the Thick-Walled and Dwarfed Culm of Shidu Bamboo (Phyllostachys nidularia f. farcta). Front. Plant Sci. 2022, 13, 876658. [Google Scholar] [CrossRef]
- Zhang, W.; Tian, G.; Polle, A.; Janz, D.; Euring, D.; Yue, X.; Zhao, H.; Fei, B.; Jiang, Z. Comparative characterization of ethanol organosolv lignin polymer from bamboo green, timber and yellow. Wood Sci. Technol. 2018, 52, 1331–1341. [Google Scholar] [CrossRef]
- Zhang, W.; Fei, B.; Polle, A.; Euring, D.; Tian, G.; Yue, X.; Chang, Y.; Jiang, Z.; Hu, T. Crystal and thermal response of cellulose isolation from bamboo by Two different chemical treatments. Bioresour. Technol. 2019, 14, 3471–3480. [Google Scholar]
- Zhang, W.B.; Fei, B.H.; Hu, T.; Ma, Y.J.; Chang, Y.T.; Xia, M.S.; Fan, K.K.; Jiang, Z.H. Comparative Study on Cell Structure Traits of Moso Bamboo (Phyllostachys edulis) in Different Areas. J. Mater. Sci. Eng. 2020, 38, 431–438. [Google Scholar]
- Xie, J.; Qi, J.; Hu, T.; De Hoop, C.F.; Hse, C.Y.; Shupe, T.F. Effect of fabricated density and bamboo species on physical–mechanical properties of bamboo fiber bundle reinforced composites. J. Mater. Sci. 2016, 51, 7480–7490. [Google Scholar] [CrossRef]
- Zou, L.; Jin, H.; Lu, W.-Y.; Li, X. Nanoscale structural and mechanical characterization of the cell wall of bamboo fibers. Mater. Sci. Eng. C 2009, 29, 1375–1379. [Google Scholar] [CrossRef]
- Costa, M.M.E.; Melo, S.L.S.; Santos, J.V.M.; Araújo, E.A.; Cunha, G.P.; Deus, E.P.; Schmitt, N. Influence of physical and chemical treatments on the mechanical properties of bamboo fibers. Procedia Eng. 2017, 200, 457–464. [Google Scholar] [CrossRef]
- Song, J.F.; Xu, S.C.; Wang, H.X.; Wu, X.Q.; Zou, M. Bionic design and multi-objective optimization for variable wall thickness tube inspired bamboo structures. Thin-Walled Struct. 2018, 125, 76–88. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, T.; Chang, Y.; Fei, B.; Ma, Y.; Deng, Y.; Xia, M.; Fan, K.; Zhang, X.; Jiang, Z. Correlation between Genetic Characteristics, Cell Structure and Material Properties of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) in Different Areas of China. Forests 2022, 13, 107. [Google Scholar] [CrossRef]
- Lever, J.; Krzywinski, M.; Altman, N. Principal component analysis. Nat. Methods 2017, 14, 641–642. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Shancen, Z.; Fei, B.; Liu, H.; Yang, H.; Dai, H.; Wang, D.; Jin, W.; Tang, F.; Gao, Q.; et al. Announcing the Genome Atlas of Bamboo and Rattan (GABR) project: Promoting research in evolution and in economically and ecologically beneficial plants. GigaScience 2017, 6, gix046. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Liu, L.; Wang, X.; Liang, L.; Yue, J.; Li, L. Comparative Analyses of Anatomical Structure, Phytohormone Levels, and Gene Expression Profiles Reveal Potential Dwarfing Mechanisms in Shengyin Bamboo (Phyllostachys edulis f. tubaeformis). Int. J. Mol. Sci. 2018, 19, 1697. [Google Scholar] [CrossRef] [Green Version]
- Hou, D. Moso Bamboo Transposon. In The Moso Bamboo Genome; Gao, J., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 57–68. [Google Scholar]
- Isagi, Y.; Oda, T.; Fukushima, K.; Lian, C.; Yokogawa, M.; Kaneko, S. Predominance of a single clone of the most widely distributed bamboo species Phyllostachys edulis in East Asia. J. Plant Res. 2016, 129, 21–27. [Google Scholar] [CrossRef]
- Gottlieb, L.D. Genetics and Morphological Evolution in Plants. Am. Nat. 1984, 123, 681–709. [Google Scholar] [CrossRef]
- Henderson, I.R.; Salt, D.E. Natural genetic variation and hybridization in plants. J. Exp. Bot. 2017, 68, 5415–5417. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, S.-V.; Katche, E.; Ihien, E.; Chawla, H.S.; Mason, A.S. The role of genomic structural variation in the genetic improvement of polyploid crops. Crop J. 2019, 7, 127–140. [Google Scholar] [CrossRef]
- Robertson, A. (Ed.) Natural Selection and Continuous Variation1980; Springer: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Lu, L.; Shao, D.; Qiu, X.; Sun, L.; Yan, W.; Zhou, X.; Yang, L.; He, Y.; Yu, S.; Xing, Y. Natural variation and artificial selection in four genes determine grain shape in rice. New Phytol. 2013, 200, 1269–1280. [Google Scholar] [CrossRef]

{kind=link}
| Abbreviation | Species and Variety |
|---|---|
| M | Phyllostachys edulis (Carr.) H.de Lehaie |
| HC | Phyllostachys edulis (Carr.) H.de Lehaie f. luteosulcata (Wen) Chao et Renv. |
| LC | Phyllostachys edulis (Carr.) H.de Lehaie f. bicolor (Nakai) G.H.Lai |
| JS | Phyllostachys edulis (Carr.) H.de Lehaie f. gracilis (Hsiung) Chao et Renv. |
| HPH | Phyllostachys edulis (Carr.) H.de Lehaie f. huamozhu (Wen) Chao et Renv. |
| LPH | Phyllostachys edulis (Carr.) H.de Lehaie f. nabeshimana (Muroi) Chao et Renv. |
| Q | Phyllostachys edulis (Carr.) H.de Lehaie f. obliquinoda (Z.P.Wang et N.X.Ma) Ohrnberger |
| HB | Phyllostachys edulis (Carr.) H.de Lehaie f. pachyloen (G.Y.Yang et al.) Y.L.Ding ex G.H.Lai |
| SY | Phyllostachys edulis (Carr.) H.de Lehaie f. tubaeformis (S.Y.Wang) Ohrnberger |
| GJ | Phyllostachys edulis (Carr.) H.de Lehaie ’Kikko-chiku’,G.H.Lai |
| Traits | Site | Mean | Std DeV. | CV /% | Max | Min | Range |
|---|---|---|---|---|---|---|---|
| Tangential length of vascular bundle/μm | M | 377.50a | 44.16 | 11.70 | 476.95 | 279.23 | 197.72 |
| HC | 458.45d | 33.91 | 7.40 | 540.35 | 408.36 | 131.99 | |
| LC | 393.18a | 30.14 | 7.67 | 448.66 | 331.53 | 117.13 | |
| JS | 526.04f | 47.13 | 8.96 | 627.11 | 426.97 | 200.14 | |
| HPH | 432.00bc | 37.69 | 8.73 | 500.35 | 356.79 | 143.56 | |
| LPH | 495.87e | 25.91 | 5.22 | 533.27 | 442.86 | 90.41 | |
| Q | 463.77d | 42.82 | 9.23 | 581.61 | 400.28 | 181.33 | |
| HB | 419.72b | 41.78 | 9.95 | 535.91 | 340.63 | 195.28 | |
| SY | 444.78cd | 29.23 | 6.57 | 489.90 | 383.12 | 106.78 | |
| GJ | 379.98a | 38.12 | 10.03 | 495.63 | 324.07 | 171.56 | |
| Radial length of vascular bundle/μm | M | 421.6ab | 54.25 | 12.87 | 519.98 | 317.58 | 202.40 |
| HC | 532.99c | 31.22 | 5.86 | 619.27 | 480.33 | 138.94 | |
| LC | 406.54a | 46.92 | 11.54 | 601.77 | 360.97 | 240.80 | |
| JS | 527.05c | 44.88 | 8.51 | 621.33 | 448.53 | 172.80 | |
| HPH | 526.59c | 26.95 | 5.12 | 578.63 | 478.93 | 99.70 | |
| LPH | 576.67e | 39.54 | 6.86 | 659.67 | 476.89 | 182.78 | |
| Q | 567.38de | 42.86 | 7.55 | 653.55 | 482.44 | 171.11 | |
| HB | 550.22cd | 57.93 | 10.53 | 650.07 | 422.25 | 227.82 | |
| SY | 442.46b | 24.63 | 5.57 | 500.50 | 406.92 | 93.58 | |
| GJ | 399.31a | 28.76 | 7.20 | 473.21 | 345.62 | 127.59 | |
| Radial/tangential ratio of vascular bundle/% | M | 1.13bc | 0.17 | 15.34 | 1.56 | 0.88 | 0.68 |
| HC | 1.17cd | 0.12 | 10.22 | 1.43 | 1.00 | 0.42 | |
| LC | 1.04a | 0.12 | 11.92 | 1.44 | 0.87 | 0.57 | |
| JS | 1.01a | 0.14 | 13.38 | 1.20 | 0.79 | 0.41 | |
| HPH | 1.23d | 0.11 | 9.22 | 1.47 | 0.99 | 0.48 | |
| LPH | 1.17cd | 0.09 | 7.82 | 1.35 | 1.04 | 0.32 | |
| Q | 1.24d | 0.16 | 13.12 | 1.53 | 0.93 | 0.60 | |
| HB | 1.33e | 0.24 | 17.74 | 1.87 | 0.86 | 1.01 | |
| SY | 1.00a | 0.09 | 9.11 | 1.18 | 0.86 | 0.32 | |
| GJ | 1.06ab | 0.09 | 8.68 | 1.23 | 0.86 | 0.36 |
| Table | Source | df | MS | F Value | p-Value |
|---|---|---|---|---|---|
| Tangential length of vascular bundle/μm | Between Groups | 9 | 73,776.550 | 51.622 | 1.66 × 10−55 **** |
| Within Groups | 295 | 1429.164 | |||
| Total | 304 | ||||
| Radial length of vascular bundle/μm | 9 | 147,911.805 | 85.635 | 6.04 × 10−77 **** | |
| 295 | 1727.239 | ||||
| 304 | |||||
| Radial/tangential ratio of vascular bundle/% | Between Groups | 9 | 0.365 | 18.223 | 4.82 × 10−24 **** |
| Within Groups | 295 | 0.020 | |||
| Total | 304 |
| Traits | Species | Mean | Std DeV. | CV /% | Max | Min | Range |
|---|---|---|---|---|---|---|---|
| Fiber length/μm | M | 2411.27e | 143.98 | 5.97 | 2662.02 | 2166.81 | 495.21 |
| HC | 2241.66de | 346.71 | 15.47 | 2750.63 | 1722.90 | 1027.73 | |
| LC | 1962.37bc | 341.48 | 17.40 | 2463.80 | 1360.91 | 1102.89 | |
| JS | 1940.25bc | 493.87 | 25.45 | 3079.47 | 1279.60 | 1799.87 | |
| HPH | 2305.85de | 561.78 | 24.36 | 3482.49 | 1147.26 | 2335.23 | |
| LPH | 2133.10cd | 277.72 | 13.02 | 2991.63 | 1668.24 | 1323.39 | |
| Q | 2089.40cd | 530.89 | 25.41 | 3325.26 | 1156.91 | 2168.35 | |
| HB | 1923.98bc | 412.73 | 21.45 | 2520.58 | 1168.42 | 1352.16 | |
| SY | 1282.30a | 143.46 | 11.19 | 1531.01 | 1008.68 | 522.33 | |
| GJ | 1766.72b | 275.79 | 15.61 | 2332.18 | 1221.45 | 1110.73 | |
| Fiber width/μm | M | 23.10d | 1.99 | 8.63 | 27.03 | 19.48 | 7.55 |
| HC | 20.16c | 3.17 | 15.74 | 27.75 | 13.71 | 14.04 | |
| LC | 14.57a | 2.93 | 20.13 | 24.53 | 8.29 | 16.24 | |
| JS | 14.74a | 2.72 | 18.45 | 22.29 | 10.11 | 12.18 | |
| HPH | 17.81b | 3.56 | 19.97 | 25.66 | 11.55 | 14.11 | |
| LPH | 18.20b | 4.54 | 24.94 | 27.40 | 10.13 | 17.27 | |
| Q | 15.06a | 3.04 | 20.17 | 20.38 | 8.46 | 11.92 | |
| HB | 14.23a | 2.42 | 16.99 | 21.32 | 8.70 | 12.62 | |
| SY | 13.21a | 1.94 | 14.67 | 16.71 | 7.64 | 9.07 | |
| GJ | 14.12a | 3.06 | 21.67 | 18.94 | 7.15 | 11.79 | |
| Fiber cell wall thickness /μm | M | 7.26b | 0.70 | 9.64 | 8.66 | 5.84 | 2.82 |
| HC | 8.67bcd | 2.03 | 23.47 | 12.49 | 3.26 | 9.24 | |
| LC | 8.36bc | 1.85 | 22.15 | 11.77 | 5.21 | 6.56 | |
| JS | 7.80bc | 2.06 | 26.40 | 12.64 | 4.16 | 8.48 | |
| HPH | 8.96cd | 2.24 | 25.03 | 13.72 | 5.06 | 8.66 | |
| LPH | 8.32bc | 1.75 | 21.07 | 11.77 | 5.47 | 6.30 | |
| Q | 2.96a | 1.86 | 62.61 | 12.54 | 1.67 | 10.87 | |
| HB | 8.26bc | 2.70 | 32.73 | 14.24 | 3.14 | 11.10 | |
| SY | 9.74d | 2.26 | 23.19 | 14.45 | 5.93 | 8.52 | |
| GJ | 8.01bc | 1.57 | 19.54 | 12.98 | 5.26 | 7.72 | |
| Fiber lumen diameter /μm | M | 3.36b | 0.34 | 10.15 | 4.30 | 2.57 | 1.73 |
| HC | 2.44a | 0.42 | 17.14 | 3.51 | 1.67 | 1.84 | |
| LC | 2.35a | 0.44 | 18.84 | 3.28 | 1.12 | 2.16 | |
| JS | 2.30a | 0.57 | 24.72 | 3.07 | 1.16 | 1.91 | |
| HPH | 2.17a | 0.57 | 26.21 | 3.50 | 1.12 | 2.39 | |
| LPH | 2.51a | 0.49 | 19.65 | 3.35 | 1.07 | 2.28 | |
| Q | 4.79c | 1.60 | 33.45 | 6.96 | 1.23 | 5.73 | |
| HB | 2.64a | 0.63 | 23.68 | 3.72 | 1.53 | 2.19 | |
| SY | 2.40a | 0.64 | 26.65 | 3.42 | 1.10 | 2.32 | |
| GJ | 2.32a | 0.68 | 29.18 | 5.01 | 1.14 | 3.87 |
| Traits | Source | df | MS | F Value | p-Value |
|---|---|---|---|---|---|
| Fiber length/μm | Between Groups | 9 | 3,501,009.571 | 24.262 | 8.70 × 10−32 **** |
| Within Groups | 330 | 144,300.619 | |||
| Total | 339 | ||||
| Fiber width /μm | Between Groups | 9 | 348.208 | 38.058 | 4.82 × 10−46 **** |
| Within Groups | 330 | 9.149 | |||
| Total | 339 | ||||
| Fiber cell wall thickness/μm | Between Groups | 9 | 101.432 | 26.520 | 2.46 × 10−33 **** |
| Within Groups | 294 | 3.825 | |||
| Total | 303 | ||||
| Fiber lumen diameter/μm | Between Groups | 9 | 19.164 | 37.210 | 1.12 × 10−43 **** |
| Within Groups | 294 | 0.515 | |||
| Total | 303 |
| Traits | Site | Mean | Std DeV. | CV /% | Max | Min | Range |
|---|---|---|---|---|---|---|---|
| Parenchyma length/μm | M | 135.45e | 10.43 | 7.70 | 154.05 | 113.79 | 40.26 |
| HC | 91.06c | 18.16 | 19.94 | 146.58 | 61.60 | 84.98 | |
| LC | 85.00bc | 23.41 | 27.54 | 126.96 | 44.05 | 82.91 | |
| JS | 76.56ab | 21.75 | 28.41 | 129.72 | 42.52 | 87.20 | |
| HPH | 104.67d | 26.49 | 25.31 | 170.87 | 64.34 | 106.53 | |
| LPH | 69.56a | 20.58 | 29.58 | 128.86 | 40.10 | 88.76 | |
| Q | 67.97a | 16.18 | 23.80 | 103.06 | 42.19 | 60.87 | |
| HB | 72.27a | 18.47 | 25.55 | 116.72 | 44.78 | 71.94 | |
| SY | 69.53a | 23.41 | 33.67 | 116.86 | 35.02 | 81.84 | |
| GJ | 65.48a | 20.21 | 30.87 | 106.71 | 33.45 | 73.26 | |
| Parenchyma width /μm | M | 51.63cd | 4.37 | 8.47 | 58.99 | 43.77 | 15.22 |
| HC | 48.21c | 8.42 | 17.46 | 70.26 | 32.57 | 37.69 | |
| LC | 42.34b | 7.45 | 17.59 | 59.38 | 24.44 | 34.94 | |
| JS | 38.73b | 10.99 | 28.38 | 69.55 | 22.11 | 47.44 | |
| HPH | 54.05d | 11.92 | 22.05 | 75.05 | 31.67 | 43.38 | |
| LPH | 39.16b | 8.06 | 20.59 | 61.52 | 23.87 | 37.65 | |
| Q | 39.58b | 7.40 | 18.69 | 53.60 | 21.03 | 32.57 | |
| HB | 32.86a | 7.37 | 22.42 | 43.97 | 20.73 | 23.24 | |
| SY | 33.20a | 7.53 | 22.69 | 53.19 | 19.13 | 34.06 | |
| GJ | 30.70a | 5.95 | 19.37 | 44.65 | 21.31 | 23.34 | |
| Parenchyma wall thickness /μm | M | 5.64de | 0.95 | 16.88 | 8.22 | 3.42 | 4.80 |
| HC | 5.14cde | 1.36 | 26.57 | 8.05 | 2.63 | 5.42 | |
| LC | 4.57bc | 0.89 | 19.50 | 6.81 | 3.16 | 3.66 | |
| JS | 5.73e | 1.03 | 17.88 | 7.80 | 3.50 | 4.29 | |
| HPH | 4.39bc | 0.80 | 18.16 | 5.76 | 2.94 | 2.82 | |
| LPH | 5.01cd | 0.93 | 18.58 | 8.33 | 3.16 | 5.18 | |
| Q | 2.69a | 1.80 | 67.05 | 9.15 | 1.00 | 8.15 | |
| HB | 4.18b | 0.75 | 17.98 | 6.32 | 2.77 | 3.55 | |
| SY | 4.48bc | 0.95 | 21.25 | 6.58 | 2.80 | 3.78 | |
| GJ | 4.08b | 1.04 | 25.58 | 6.84 | 2.36 | 4.48 | |
| Parenchyma lumen diameter/μm | M | 25.39b | 3.22 | 12.69 | 34.04 | 20.08 | 13.96 |
| HC | 33.98c | 9.57 | 28.16 | 53.52 | 16.83 | 36.69 | |
| LC | 26.35bc | 11.53 | 43.74 | 46.07 | 0.00 | 46.07 | |
| JS | 25.77b | 10.93 | 42.41 | 45.60 | 10.23 | 35.37 | |
| HPH | 28.67bc | 9.31 | 32.47 | 48.37 | 11.05 | 37.32 | |
| LPH | 33.69c | 10.33 | 30.64 | 49.70 | 14.59 | 35.11 | |
| Q | 10.73a | 10.12 | 94.30 | 41.19 | 3.07 | 38.12 | |
| HB | 27.23bc | 9.60 | 35.27 | 49.99 | 10.08 | 39.90 | |
| SY | 33.17c | 12.54 | 37.79 | 57.64 | 13.68 | 43.95 | |
| GJ | 23.33b | 9.16 | 39.25 | 46.15 | 10.55 | 35.60 |
| Traits | Source | df | MS | F Value | p-Value |
|---|---|---|---|---|---|
| Parenchyma length/μm | Between Groups | 9 | 16,378.346 | 39.523 | 2.09 × 10−47 **** |
| Within Groups | 330 | 414.403 | |||
| Total | 339 | ||||
| Parenchyma width/μm | Between Groups | 9 | 2081.207 | 27.665 | 1.45 × 10 35 **** |
| Within Groups | 330 | 75.228 | |||
| Total | 339 | ||||
| Parenchyma wall thickness/μm | Between Groups | 9 | 23.756 | 20.003 | 3.91 × 10−26 **** |
| Within Groups | 294 | 1.188 | |||
| Total | 303 | ||||
| Parenchyma lumen diameter/μm | Between Groups | 9 | 1390.235 | 14.634 | 1.45 × 10−19 **** |
| Within Groups | 293 | 95.001 | |||
| Total | 302 |
| Principal Component | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 | PC10 | PC11 | |
| Proportion of Variance | 22.18 | 19.45 | 13.68 | 11.85 | 7.65 | 6.13 | 6.00 | 5.46 | 4.081 | 3.46 | 0.06 |
| Cumulative Proportion | 22.18 | 41.62 | 55.30 | 67.16 | 74.80 | 80.94 | 86.94 | 92.40 | 96.48 | 99.94 | 100 |
| Standard Deviations | 1.5618 | 1.4626 | 1.2268 | 1.1418 | 0.9172 | 0.8215 | 0.8125 | 0.7751 | 0.6700 | 0.6167 | 0.0835 |
| Trait | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Tangential length of vascular bundle/μm | −0.1133 | 0.0744 | −0.6133 | −0.5205 |
| Radial length of vascular bundle/μm | 0.0862 | 0.4308 | −0.5861 | 0.1771 |
| Radial/tangential ratio of vascular bundle/% | 0.1874 | 0.3864 | −0.0786 | 0.6528 |
| Fiber length/μm | 0.4545 | 0.0634 | −0.0905 | −0.1345 |
| Fiber width/μm | 0.5062 | −0.1030 | −0.0360 | −0.0815 |
| Fiber cell wall thickness/μm | 0.1025 | 0.4203 | 0.1862 | −0.1649 |
| Fiber lumen diameter/μm | −0.0739 | −0.4157 | −0.1681 | 0.2871 |
| Parenchyma length/μm | 0.4685 | −0.1713 | 0.1728 | −0.0172 |
| Parenchyma width/μm | 0.4751 | −0.0595 | −0.0617 | −0.0653 |
| Parenchyma wall thickness/μm | −0.0271 | −0.3520 | −0.3177 | 0.3605 |
| Parenchyma lumen diameter/μm | 0.1413 | −0.3724 | −0.2591 | −0.0511 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Jiang, Z.; Chang, Y.; Fei, B.; Ma, Y.; Deng, Y.; Zhang, X.; Hu, T. Analysis of the Cell Structural Characters of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) and Its Varieties. Forests 2023, 14, 235. https://doi.org/10.3390/f14020235
Zhang W, Jiang Z, Chang Y, Fei B, Ma Y, Deng Y, Zhang X, Hu T. Analysis of the Cell Structural Characters of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) and Its Varieties. Forests. 2023; 14(2):235. https://doi.org/10.3390/f14020235
Chicago/Turabian StyleZhang, Wenbo, Zehui Jiang, Yanting Chang, Benhua Fei, Yanjun Ma, Yayun Deng, Xue Zhang, and Tao Hu. 2023. "Analysis of the Cell Structural Characters of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) and Its Varieties" Forests 14, no. 2: 235. https://doi.org/10.3390/f14020235