
Ecological and Soil Data Applied to Conservation Management of an Urban Forest

Abstract
:1. Introduction
2. Materials and Methods
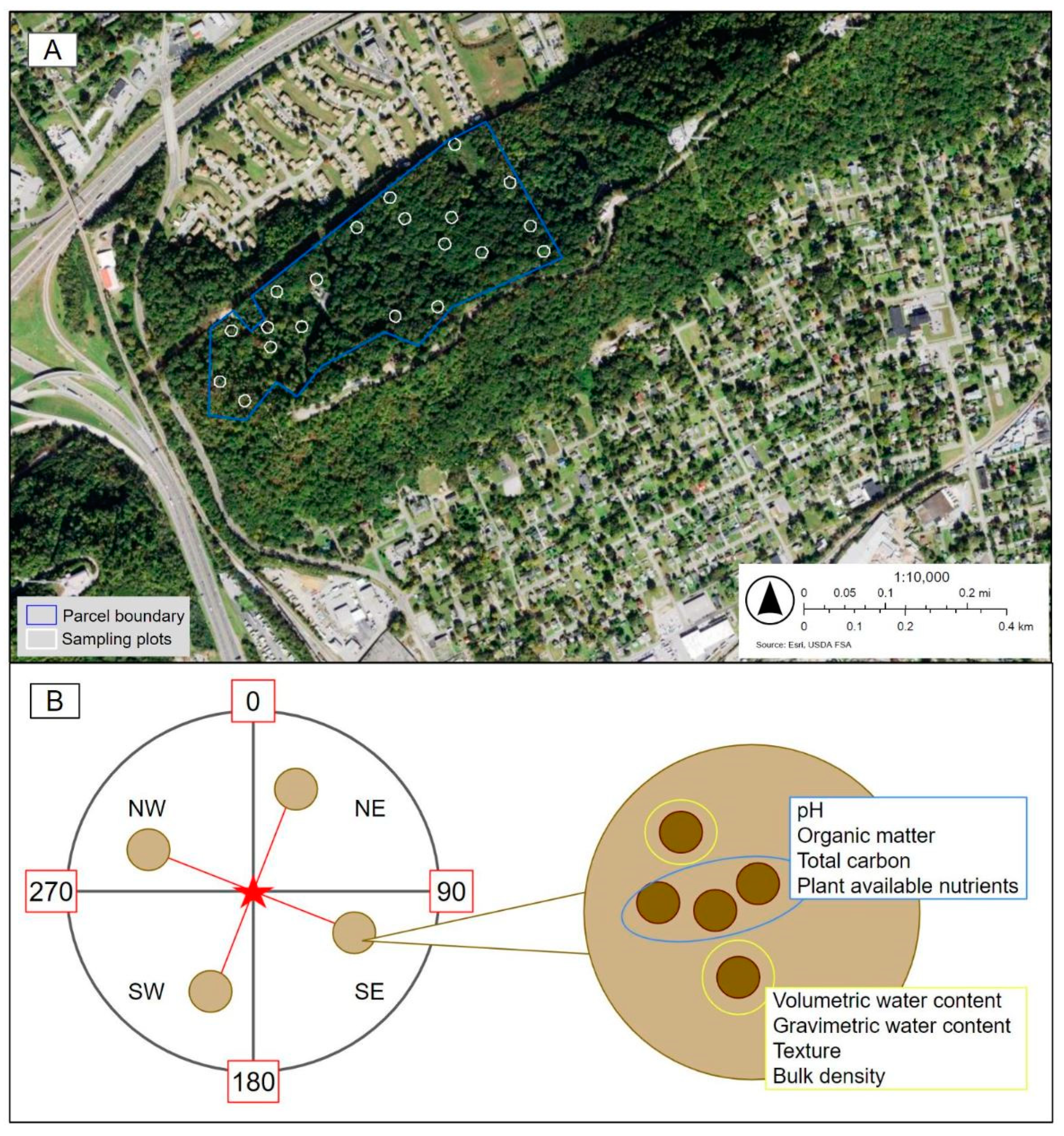
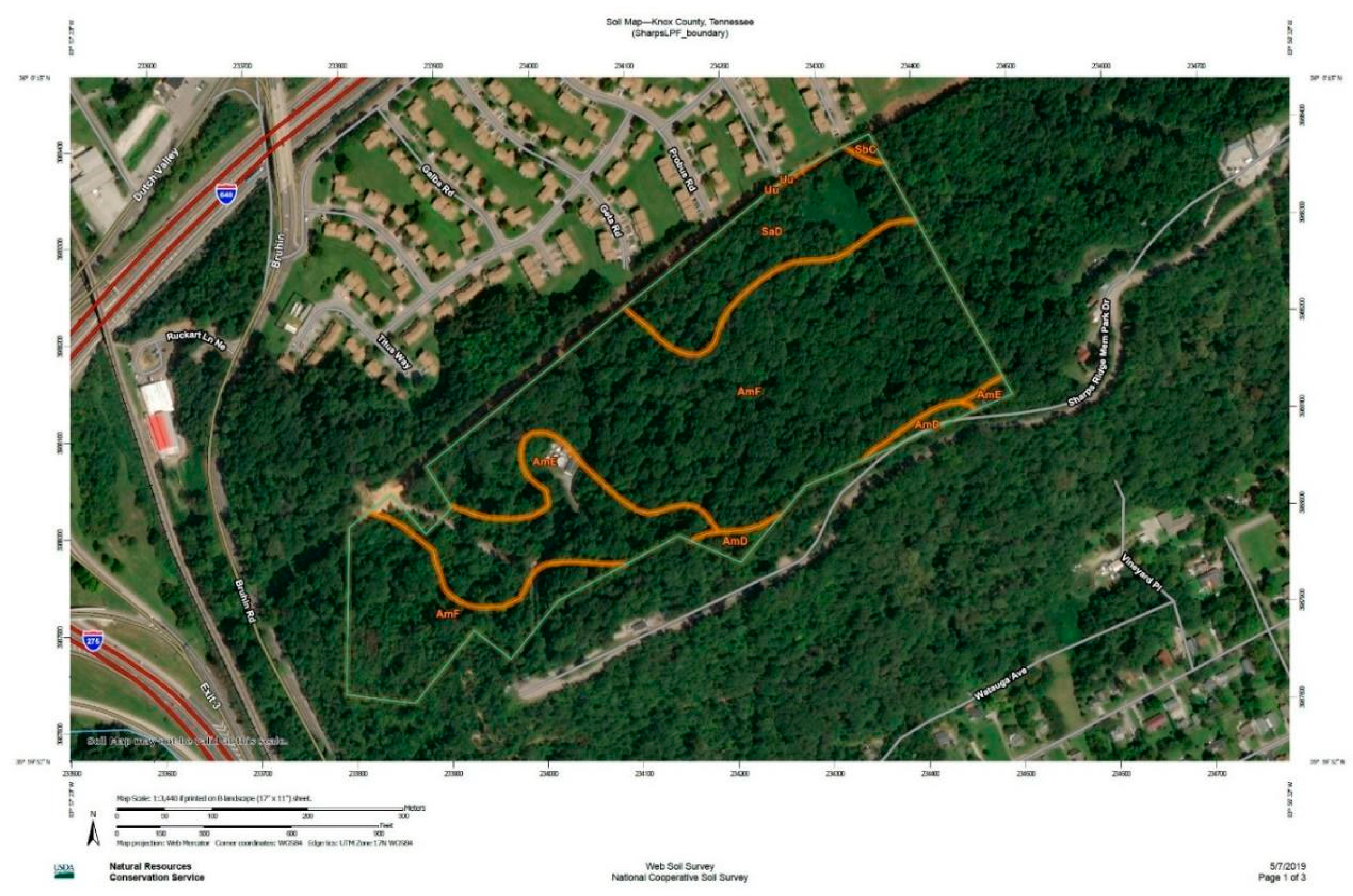
2.1. Study Site

2.2. Ecological Data Collection
2.3. Soil Collection and Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Urban Forest Composition and Condition
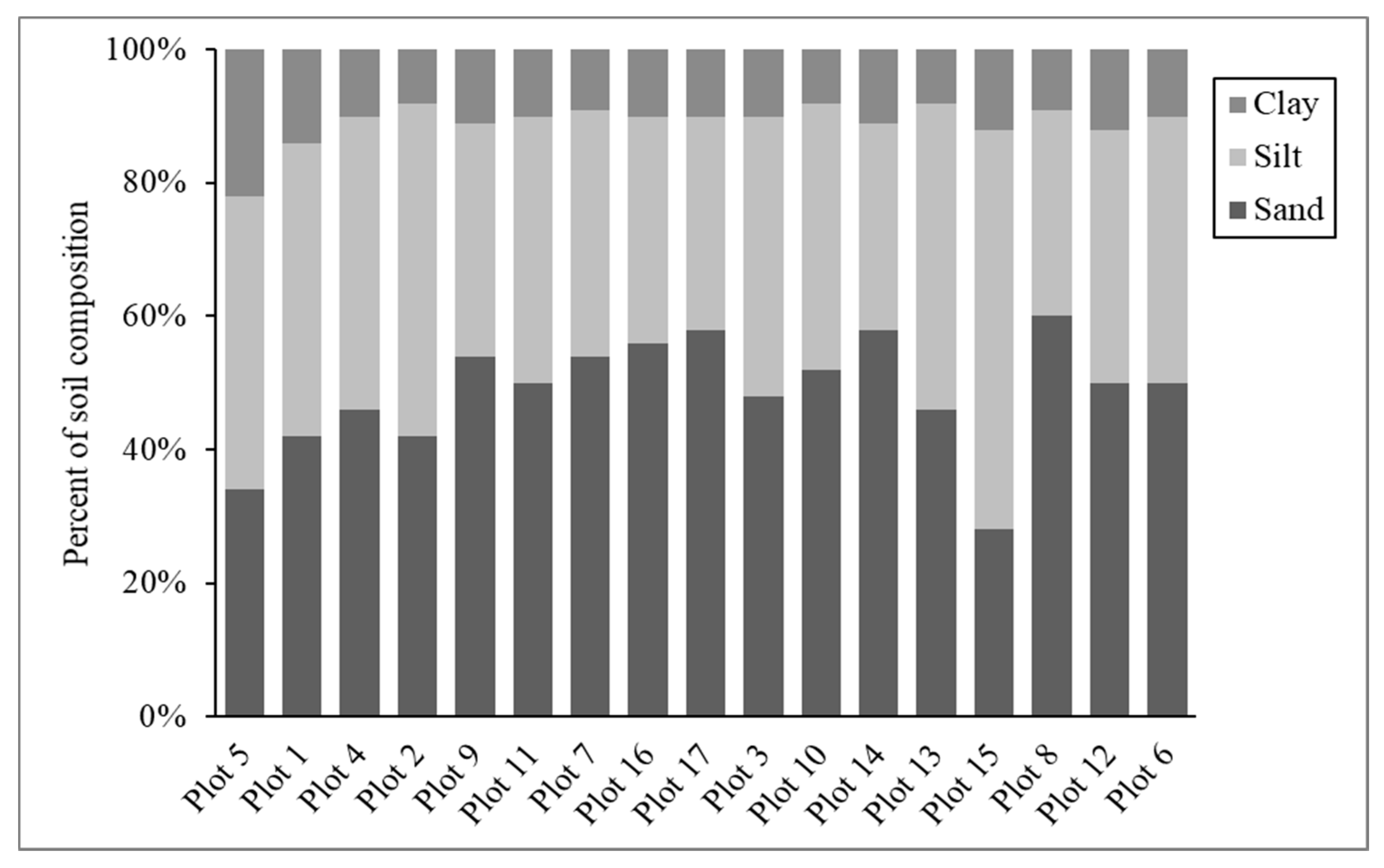
3.2. Soil Attributes
3.3. Relationships between Soil Attributes and Forest Bioindicators
Richness, Diversity, Equitability, Native Abundance across Strata
3.4. Within-Strata Diversity
3.5. Plot-Level Impacts
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
| Map Unit Symbol | Map Unit Name | Hectares in AOI | Percent of AOI |
|---|---|---|---|
| AmD | Apison-Montevallo complex, 12 to 25 percent slopes | 0.32 | 1.8% |
| AmE | Apison-Montevallo complex, 25–35 percent slopes, rocky | 2.43 | 14.1% |
| AmF | Apison-Montevallo complex, 35 to 75 percent slopes, rocky | 12.06 | 69.8% |
| SaD | Salacoa gravelly loam, 12 to 25 percent slopes | 2.39 | 13.9% |
| SbC | Salcoa-Apison complex, 5 to 12 percent slopes | 0.04 | 0.3% |
| Uu | Urban land-Udorthents complex | 0.0 | 0.1% |
| Totals for Area of Interest | 17.24 | 100.0% | |
| PLOT | LAT | LONG |
|---|---|---|
| Plot 1 | 36.0025106 | −83.94927621 |
| Plot 2 | 36.00197844 | −83.95000384 |
| Plot 3 | 36.00020893 | −83.95195902 |
| Plot 4 | 36.00213861 | −83.94895458 |
| Plot 5 | 36.00345394 | −83.94786174 |
| Plot 6 | 36.00155746 | −83.94590983 |
| Plot 7 | 35.99924814 | −83.9530034 |
| Plot 8 | 36.00040706 | −83.94916099 |
| Plot 9 | 36.00277919 | −83.94664926 |
| Plot 10 | 35.99985932 | −83.95189211 |
| Plot 11 | 36.00015073 | −83.95275281 |
| Plot 12 | 36.00057602 | −83.94823461 |
| Plot 13 | 35.9989089 | −83.9524609 |
| Plot 14 | 36.00200854 | −83.94619979 |
| Plot 15 | 36.00153933 | −83.94726715 |
| Plot 16 | 36.00022314 | −83.95121101 |
| Plot 17 | 36.0010602 | −83.95088963 |

| Plot | Aspect | Avg. Slope | Texture | Avg. Volumetric Water Content (%) | Avg. Bulk Density (g/mL) | Ideal (I), Affected (A), Restricted (R) * |
|---|---|---|---|---|---|---|
| Plot 1 | W | 6.2 | Loam | 51.25 | 1.77 | A |
| Plot 2 | NW | 19.2 | Silt Loam | 44.63 | 1.33 | I |
| Plot 3 | S | 13.8 | Loam | 51.38 | 1.62 | A |
| Plot 4 | W | 11.6 | Loam | 48.88 | 1.72 | A |
| Plot 5 | SW | 13.0 | Loam | 47.25 | 1.82 | R |
| Plot 6 | N | 24.0 | Loam | 35.50 | 1.30 | I |
| Plot 7 | NW | 22.8 | Sandy Loam | 33.50 | 1.12 | I |
| Plot 8 | NE | 25.1 | Sandy Loam | 34.13 | 1.06 | I |
| Plot 9 | W | 11.5 | Sandy Loam | 41.50 | 1.19 | I |
| Plot 10 | SW | 17.4 | Loam | 35.38 | 1.24 | I |
| Plot 11 | SW | 14.8 | Loam | 52.50 | 1.82 | R |
| Plot 12 | NW | 17.6 | Loam | 39.88 | 1.20 | I |
| Plot 13 | W | 21.6 | Loam | 31.13 | 1.39 | I |
| Plot 14 | W | 21.9 | Sandy Loam | 39.75 | 1.35 | I |
| Plot 15 | W | 23.6 | Silt Loam | 34.75 | 1.29 | I |
| Plot 16 | NW | 25.5 | Sandy Loam | 42.00 | 1.32 | I |
| Plot 17 | NE | 20.6 | Sandy Loam | 37.63 | 1.09 | I |
| Avg. | - | 18.2 | - | 41.24 | 1.39 | I |
| S.D. | - | 5.6 | - | 6.96 | .26 | - |
| n | 17 | 17 | - | 17 | 130 | - |
| Plot | Avg. % Organic Matter | Avg. % Total C | Avg. pH | P | K | Ca | Mg | B | Fe | Mn | Na | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (All Nutrients in kg/ha) | ||||||||||||
| Plot 1 | 4.4% | 2.6% | 4.31 | 9.3 | 195.6 | 470.8 | 113.2 | 0.76 | 155.3 | 201.2 | 11.5 | 11.5 |
| Plot 2 | 5.7% | 3.3% | 4.90 | 5.0 | 144.0 | 535.1 | 88.6 | 0.54 | 54.9 | 96.2 | 8.4 | 8.2 |
| Plot 3 | 6.0% | 3.5% | 5.72 | 3.1 | 209.6 | 1153.3 | 172.1 | 1.01 | 26.3 | 73.8 | 8.2 | 10.3 |
| Plot 4 | 4.0% | 2.3% | 5.17 | 5.9 | 129.5 | 310.9 | 82.7 | 0.45 | 48.2 | 82.4 | 9.0 | 7.0 |
| Plot 5 | 4.1% | 2.4% | 5.23 | 5.2 | 191.8 | 1116.6 | 212.3 | 0.78 | 38.5 | 44.2 | 10.1 | 8.4 |
| Plot 6 | 4.5% | 2.6% | 5.22 | 7.3 | 132.1 | 310.3 | 69.8 | 0.39 | 67.8 | 165.9 | 7.3 | 10.1 |
| Plot 7 | 5.2% | 3.0% | 5.87 | 2.7 | 173.6 | 1262.5 | 143.0 | 0.72 | 27.9 | 68.2 | 7.2 | 10.1 |
| Plot 8 | 5.7% | 3.3% | 5.56 | 5.0 | 213.3 | 792.5 | 125.3 | 0.65 | 34.5 | 64.5 | 6.2 | 11.7 |
| Plot 9 | 4.9% | 2.8% | 5.16 | 4.3 | 111.0 | 526.3 | 73.2 | 0.48 | 52.5 | 63.3 | 7.1 | 6.4 |
| Plot 10 | 7.4% | 4.3% | 6.08 | 7.6 | 251.1 | 1877.5 | 282.8 | 1.21 | 27.2 | 134.0 | 8.4 | 9.3 |
| Plot 11 | 6.0% | 3.5% | 5.65 | 2.0 | 200.7 | 839.4 | 163.4 | 0.73 | 44.8 | 23.9 | 8.2 | 10.8 |
| Plot 12 | 5.5% | 3.2% | 4.92 | 2.8 | 123.9 | 398.0 | 81.3 | 0.48 | 76.0 | 49.1 | 8.4 | 7.7 |
| Plot 13 | 7.4% | 4.3% | 5.88 | 3.7 | 158.1 | 1729.5 | 179.1 | 0.78 | 63.1 | 138.8 | 7.6 | 16.1 |
| Plot 14 | 4.5% | 2.6% | 4.99 | 3.1 | 123.9 | 194.5 | 47.1 | 0.37 | 76.2 | 31.9 | 7.6 | 4.8 |
| Plot 15 | 4.9% | 2.8% | 5.00 | 4.5 | 199.3 | 339.4 | 95.1 | 0.37 | 94.7 | 80.2 | 8.4 | 7.3 |
| Plot 16 | 6.7% | 3.9% | 6.03 | 7.1 | 173.5 | 2014.4 | 248.9 | 1.29 | 13.2 | 139.9 | 6.7 | 11.1 |
| Plot 17 | 9.8% | 5.7% | 5.96 | 12.7 | 315.0 | 3107.7 | 293.7 | 1.66 | 27.2 | 208.3 | 7.8 | 13.0 |
| Average | 5.7% | 3.3% | 4.31 | 5.4 | 179.1 | 998.7 | 145.4 | 0.7 | 54.6 | 98.0 | 8.1 | 9.6 |
| Std. Dev. | 0.01% | 0.01% | 4.90 | 2.7 | 52.6 | 793.2 | 76.7 | 0.3 | 33.9 | 56.8 | 1.3 | 2.7 |
| Avg. Nutrient *- | - | 5.72 | L | M | S | S | - | S | S | - | S | |
References
- Tratalos, J.; Fuller, R.A.; Warren, P.H.; Davies, R.G.; Gaston, K.J. Urban form, biodiversity potential and eco-system services. Landsc. Urban Plan. 2007, 83, 308–317. [Google Scholar] [CrossRef]
- Barrico, L.; Castro, H.; Coutinho, A.P.; Gonçalves, M.T.; Freitas, H.; Castro, P. Plant and microbial biodiversity in urban forests and public gardens: Insights for cities’ sustainable development. Urban For. Urban Green. 2018, 29, 19–27. [Google Scholar] [CrossRef]
- Pregitzer, C.C.; Ashton, M.S.; Charlop-Powers, S.; D’Amato, A.W.; Frey, B.R.; Gunther, B.; Hallett, R.A.; Pregitzer, K.S.; Woodall, C.W.; Bradford, M.A. Defining and assessing urban forests to inform management and policy. Environ. Res. Lett. 2019, 14, 085002. [Google Scholar] [CrossRef]
- Nowak, D.J.; Greenfield, E.J. US Urban Forest Statistics, Values, and Projections. J. For. 2018, 116, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Pregitzer, C.; Forgione, H.; King, K.; Charlop-Powers, S.; Greenfeld, J. Forest Management Framework for New York City; Natural Areas Conservancy: New York, NY, USA, 2018; pp. 1–44. [Google Scholar]
- McKinney, M.L.; Ingo, K.; Kendal, D. The contribution of wild urban ecosystems to liveable cities. Urban For. Urban Green. 2018, 29, 334–335. [Google Scholar] [CrossRef]
- Miller, J.R. Biodiversity conservation and the extinction of experience. Trends Ecol. Evol. 2005, 20, 430–434. [Google Scholar] [CrossRef]
- Kahn, P.H.; Kellert, S.R. Experiencing Nature: Affective, Cognitive, and Evaluative Development in Children. In Children and Nature: Psychological, Sociocultural, and Evolutionary Investigations; The MIT Press: Cambridge, MA, USA, 2002; p. 117151. [Google Scholar] [CrossRef]
- Bowler, D.E.; Buyung-Ali, L.M.; Knight, T.M.; Pullin, A.S. A systematic review of evidence for the added benefits to health of exposure to natural environments. BMC Public Health 2010, 10, 456. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.Y.; Jim, C.Y. Assessment and Valuation of the Ecosystem Services Provided by Urban Forests. In Ecology, Planning, and Management of Urban Forests International Perspectives; Carreiro, M.M., Song, Y.C., Wu, J., Eds.; Springer: New York, NY, USA, 2008; pp. 53–83. [Google Scholar] [CrossRef]
- Lovell, S.T.; Taylor, J.R. Supplying urban ecosystem services through multifunctional green infra-structure in the United States. Landsc. Ecol. 2013, 28, 1447–1463. [Google Scholar] [CrossRef]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Nielsen, A.B.; Bosch, M.V.D.; Maruthaveeran, S.; Bosch, C.K.V.D. Species richness in urban parks and its drivers: A review of empirical evidence. Urban Ecosyst. 2014, 17, 305–327. [Google Scholar] [CrossRef]
- Zefferman, E.P.; McKinney, M.L.; Cianciolo, T.; Fritz, B.I. Knoxville’s urban wilderness: Moving toward sus-tainable multifunctional management. Urban For. Urban Green. 2018, 29, 357–366. [Google Scholar] [CrossRef]
- Pregitzer, C.C.; Charlop-Powers, S.; Bibbo, S.; Forgione, H.M.; Gunther, B.; Hallett, A.; Bradford, M.A. A city-scale assessment reveals that nativeforest types and overstory species dominate New York City forests. Ecol. Appl. 2019, 29, e01819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dearborn, D.C.; Kark, S. Motivations for Conserving Urban Biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.J.; Crane, D.E.; Stevens, J.C.; Hoehn, R.E.; Walton, J.T.; Bond, J. A Ground-Based Method of Assessing Urban Forest Structure and Ecosystem Services. Arboric. Urban For. 2008, 34, 347–358. [Google Scholar] [CrossRef]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Konijnendijk, C.C. New Perspectives for Urban Forests: Introducing Wild Woodlands. In Wild Urban Woodlands; Springer: Berlin/Heidelberg, Germany, 2005; pp. 33–45. [Google Scholar] [CrossRef]
- Forgione, H.M.; Pregitzer, C.C.; Charlop-Powers, S.; Gunther, B. Advancing urban ecosystem governance in New York City: Shifting towards a unified perspective for conservation management. Environ. Sci. Policy 2016, 62, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Pavao-Zuckerman, M.A. The Nature of Urban Soils and Their Role in Ecological Restoration in Cities. Restor. Ecol. 2008, 16, 642–649. [Google Scholar] [CrossRef]
- Riddle, R. Urban Soils. [Online] United States Department of Agriculture, Natural Resources Conservation Service. 2010. Available online: https://www.nrcs.usda.gov/wps/-portal/nrcs/main/soils/use/urban/ (accessed on 8 July 2020).
- Blanchart, A.; Séré, G.; Cherel, J.; Warot, G.; Stas, M.; Noël, C.J.; Morel, J.L.; Schwartz, C. Towards an opera-tional methodology to optimize ecosystem services provided by urban soils. Landsc. Urban Plan. 2018, 176, 1–9. [Google Scholar]
- Scheyer, J.M.; Hipple, K.W. Urban Soil Primer. United States Department of Agriculture, Natural Resources Conservation Service, National Soil Survey Center, Lincoln, Nebraska. 2005. Available online: http://soils.usda.gov/use (accessed on 20 July 2020).
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Groffman, P.M.; Band, L.E.; Boone, C.G.; Burch, W.R.; Grimmond, C.S.B.; Hom, J.; Jenkins, J.C.; et al. Beyond Urban Legends: An Emerging Framework of Urban Ecology, as Illustrated by the Baltimore Ecosystem Study. Bioscience 2008, 58, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Pickett, S.T.A.; Cadenasso, M.L. Altered resources, disturbance, and heterogeneity: A framework for comparing urban and non-urban soils. Urban Ecosyst. 2009, 12, 23–44. [Google Scholar] [CrossRef]
- Delbecque, N.; Dondeyne, S.; Gelaude, F.; Mouazen, A.M.; Vermeir, P.; Verdoodt, A. Urban soil properties distinguished by parent material, land use, time since urbanization, and pre-urban geomorphology. Geoderma 2022, 413, 115719. [Google Scholar] [CrossRef]
- Craul, P.J. Urban Soils: Applications and Practices; John Wiley & Sons: Hoboken, NJ, USA, 1999. [Google Scholar]
- Sauerwein, M. Urban soils–characterization, pollution and relevance in urban ecosystems. In Urban Ecology: Patterns, Processes, and Applications; Cambridge University Press: Cambridge, UK, 2011; pp. 45–58. [Google Scholar]
- Gaston, K.J.; Davies, Z.G.; Edmondson, J.L. Urban environments and ecosystem functions. In Urban Ecology; Cambridge University Press: Cambridge, UK, 2010; pp. 35–52. [Google Scholar] [CrossRef]
- O’Riordan, R.; Davies, J.; Stevens, C.; Quinton, J.N.; Boyko, C. The ecosystem services of urban soils: A review. Geoderma 2021, 395, 115076. [Google Scholar] [CrossRef]
- Morgan, J.B.; Connolly, E.L. Plant-Soil Interactions: Nutrient Uptake. Nat. Educ. Knowl. 2013, 4, 2. [Google Scholar]
- Charlop-Powers, Z.; Pregitzer, C.C.; Lemetre, C.; Ternei, M.A.; Maniko, J.; Hover, B.M.; Calle, P.Y.; McGuire, K.L.; Garbarino, J.; Forgione, H.M.; et al. Urban park soil microbiomes are a rich reservoir of natural product biosynthetic diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 14811–14816. [Google Scholar] [CrossRef] [Green Version]
- Threlfall, C.G.; Kendal, D. The distinct ecological and social roles that wild spaces play in urban ecosystems. Urban For. Urban Green. 2018, 29, 348–356. [Google Scholar] [CrossRef]
- Guilland, C.; Maron, P.A.; Damas, O.; Ranjard, L. Biodiversity of urban soils for sustainable cities. Environ. Chem. Lett. 2018, 16, 1267–1282. [Google Scholar] [CrossRef]
- Mooney, L.; Mooney, D.; Trently, D.; Nicholson, C.P. “Sharp’s Ridge Memorial Park.” Tennessee Important Bird Areas, Tennessee Wildlife Resources Agency. 2006. Available online: https://www.tnbirds.org/IBA/SitePages/SharpsRidge.htm (accessed on 22 August 2019).
- USDA Natural Resource Conservation Service. Web Soil Survey. 2019. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 7 May 2019).
- The Natural Areas Conservancy, the Natural Resources Group of the New York City Parks Department, and the U.S. Forest Service. Natural Areas Conservancy Upland and Forest Assessment Field Protocol, Unpublished Document. Available online: https://fic.naturalareasnyc.org/docs/upland-and-forest-ecological-assessment-protocol-nyc (accessed on 22 August 2019).
- IBM Corp. IBM SPSS Statistics for Windows, Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Tabachnick, B.G.; Fidell, L.S.; Ullman, J.B. Using Multivariate Statistics; Pearson: Boston, MA, USA, 2007; Volume 5, pp. 481–498. [Google Scholar]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Kowarik, I.; Hiller, A.; Planchuelo, G.; Seitz, B.; Von Der Lippe, M.; Buchholz, S. Emerging Urban Forests: Opportunities for Promoting the Wild Side of the Urban Green Infrastructure. Sustainability 2019, 11, 6318. [Google Scholar] [CrossRef] [Green Version]
- Putz, J. The Biology of Vines; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Aryal, P.C.; Aryal, C.; Bhusal, K.; Chapagain, D.; Dhamala, M.K.; Maharjan, S.R.; Chhetri, P.K. Forest structure and anthropogenic disturbances regulate plant invasion in urban forests. Urban Ecosyst. 2022, 25, 367–377. [Google Scholar] [CrossRef]
- Sonti, N.F.; Pregitzer, C.C.; Hallett, R.A. Native tree seedling growth and physiology responds to variable soil conditions of urban natural areas. Restor. Ecol. 2022, 30, e13653. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service. Soil Health: Soil pH. [Online] Soil Health for Educators, United States Department of Agriculture. 2014; pp. 1–6. Available online: https://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs142p2_-051574.pdf (accessed on 20 July 2020).
- Natural Resources Conservation Service. Soil Health: Bulk Density, Moisture, Aeration. [Online] Soil Health for Edu-cators, United States Department of Agriculture. 2019; pp. 1–11. Available online: https://www.nrcs.usda.gov/Internet/FSE_-DOCUMENTS/nrcs142p2_053260.pdf (accessed on 20 July 2020).
- Osman, K.T. Organic Matter of Forest Soils. In Forest Soils; Springer: Cham, Switzerland, 2013; pp. 63–76. [Google Scholar] [CrossRef]
- Smeck, N.E. Phosphorus dynamics in soils and landscapes. Geoderma 1985, 36, 185–199. [Google Scholar] [CrossRef]
- Kimmins, J.P. Soil. In Forest Ecology; Prentice Hall, Inc.: Upper Saddle River, NJ, USA, 1997; pp. 228–268. [Google Scholar]
- Natural Resources Conservation Service. Soil Health: Soil Phosphorous. [Online] Soil Health for Educators, United States Department of Agriculture. 2014; pp. 1–6. Available online: https://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/-nrcs142p2_051878.pdf (accessed on 22 July 2020).
- Kaiser, D.E.; Rosen, C.J. Potassium for Crop Production. [Online] University of Minnesota Extension. 2018. Available online: https://extension.umn.edu/phosphorus-and-potassium/potassium-crop-production (accessed on 22 July 2020).
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheard, R.W. Understanding Turf Management: Sand, Silt, and Clay. [Online] Michigan State University Libraries, Sports Turf Newsletter. 1991, pp. 1–3. Available online: https://archive.lib.msu.edu/tic/stnew/article/1991sep4.pdf (accessed on 22 July 2020).
- Bárcenas-Argüello, M.L.; del Carmen Gutiérrez-Castorena, M.; Terrazas, T. The Role of Soil Properties in Plant Endemism—A Revision of Conservation Strategies. In Soil Processes and Current Trends in Quality Assessment; IntechOpen: Rijeka, Croatia, 2013; pp. 381–398. [Google Scholar] [CrossRef] [Green Version]
- Hulshof, C.M.; Spasojevic, M.J. The edaphic control of plant diversity. Glob. Ecol. Biogeogr. 2020, 29, 1634–1650. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Exotic invasive species in urban wetlands: Environmental correlates and implications for wetland management. J. Appl. Ecol. 2008, 45, 1160–1169. [Google Scholar] [CrossRef]
- Gornish, E.S.; Ambrozio dos Santos, P. Invasive species cover, soil type, and grazing interact to predict long-term grassland restoration success. Restor. Ecol. 2016, 24, 222–229. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Callaway, R.M.; Thelen, G.C.; Rodriguez, A.; Holben, W.E. Soil biota and exotic plant invasion. Nature 2004, 427, 731–733. [Google Scholar] [CrossRef]
- Tóth, J.A.; Nagy, P.T.; Krakomperger, Z.; Veres, Z.; Kotroczó, Z.; Kincses, S.; Fekete, I.; Papp, M. Effect of litter fall on soil nutrient content and pH, and its consequences in view of climate change (Síkfőkút DIRT Project). Acta Sil-Vatica Lignaria Hung. 2011, 7, 75–86. [Google Scholar]
| Forest Health Indicator | Mean | S.D. | Median | IQR |
|---|---|---|---|---|
| Diversity and Equitability | ||||
| (Shannon’s H ‡ and EH §) | ||||
| Overstory (>10 cm DBH) | 1.47 ‡ | 0.51 ‡ | ||
| 0.84 § | 0.11 § | |||
| Midstory (2–10 cm DBH) | 1.75 ‡ | 0.57 ‡ | ||
| 0.71 § | 0.14 § | |||
| Combined | 2.07 ‡ | 0.78 ‡ | ||
| 0.47 § | 0.09 § | |||
| Species richness | 29.8 | 7.09 | ||
| Abundance | ||||
| Overstory | 14 | 4.57 | ||
| Midstory | 73 | 39.43 | ||
| Relative basal area | 28.7 m2/ha | 13.56 | ||
| Percent of healthy | ||||
| overstory trees | 30.80% | 14.30% | ||
| Percent of overstory trees with vines in canopy | 63.70% | 27.10% | ||
| Percent of plot impacted | ||||
| Canopy gap | 36.30% | 24.20% | ||
| Invasive vegetation | - | - | 13.50% | 25.00% |
| Tree damage | 20.30% | 12.10% | ||
| Fine woody debris (n = 16) | 22.50% | 19.50% | ||
| Soil modification (n = 16) | - | - | 10.30% | 10.50% |
| Anthropogenic dumping | - | - | 1.00% | 2.70% |
| Ecological Indicator | F Change | DF | Adj R-sq | Indep Variables | Std Err | Beta | p |
|---|---|---|---|---|---|---|---|
| Total Species Richness * | 6.34 | 1, 14 | 0.478 | Avg bulk dens | 4.97 | 0.597 | 0.005 |
| Avg Zn | 0.527 | 0.455 | 0.025 | ||||
| Over+Mid Diversity ** | 11.17 | 1, 13 | 0.768 | Avg Zn | 0.028 | 0.473 | 0.006 |
| % Silt | 0.008 | 0.452 | 0.003 | ||||
| Avg Mg | 0.001 | 0.489 | 0.005 | ||||
| Over+Mid Equitability * | 5.88 | 1, 14 | 0.537 | Avg Mg | 0 | 0.729 | 0.001 |
| % Silt | 0.002 | 0.42 | 0.029 | ||||
| Over+Mid Native Abund ** | 13.64 | 1, 14 | 0.624 | Avg Zn | 2.756 | 0.77 | 0 |
| Avg P | 2.743 | −0.595 | 0.002 | ||||
| Overstory Diversity *** | 15.19 | 1, 15 | 0.47 | Avg Total C | 0.108 | 0.709 | 0.001 |
| Midstory Diversity * | 5.54 | 1, 13 | 0.689 | Avg Zn | 0.033 | 0.703 | 0.001 |
| % Silt | 0.012 | 0.512 | 0.005 | ||||
| Avg Fe | 0.003 | −0.36 | 0.035 | ||||
| Canopy Gaps * | 5.59 | 1, 15 | 0.223 | Avg Zn | 0.022 | 0.521 | 0.032 |
| Invasive Vegetation *** | 15.77 | 1, 14 | 0.649 | % Clay | 1.165 | 0.707 | 0.001 |
| Avg Fe | 0.126 | −0.644 | 0.001 | ||||
| Fine Woody Debris * | 15.97 | 1, 12 | 0.856 | Avg pH | 6.256 | 0.936 | 0.001 |
| Avg P | 1.08 | −0.594 | 0.001 | ||||
| Avg Na | 2.852 | 0.492 | 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wavrek, M.T.; Jean-Philippe, S.; McKinney, M.L. Ecological and Soil Data Applied to Conservation Management of an Urban Forest. Forests 2023, 14, 487. https://doi.org/10.3390/f14030487
Wavrek MT, Jean-Philippe S, McKinney ML. Ecological and Soil Data Applied to Conservation Management of an Urban Forest. Forests. 2023; 14(3):487. https://doi.org/10.3390/f14030487
Chicago/Turabian StyleWavrek, Mia T., Sharon Jean-Philippe, and Michael L. McKinney. 2023. "Ecological and Soil Data Applied to Conservation Management of an Urban Forest" Forests 14, no. 3: 487. https://doi.org/10.3390/f14030487
APA StyleWavrek, M. T., Jean-Philippe, S., & McKinney, M. L. (2023). Ecological and Soil Data Applied to Conservation Management of an Urban Forest. Forests, 14(3), 487. https://doi.org/10.3390/f14030487