
Interaction Processes of Environment and Plant Ecophysiology with BVOC Emissions from Dominant Greening Trees


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Area and Tree Species
2.2. BVOC Sampling and Analysis
2.3. Measurements of the Environment and Plant Ecophysiology
2.4. Statistical Analyses
3. Results
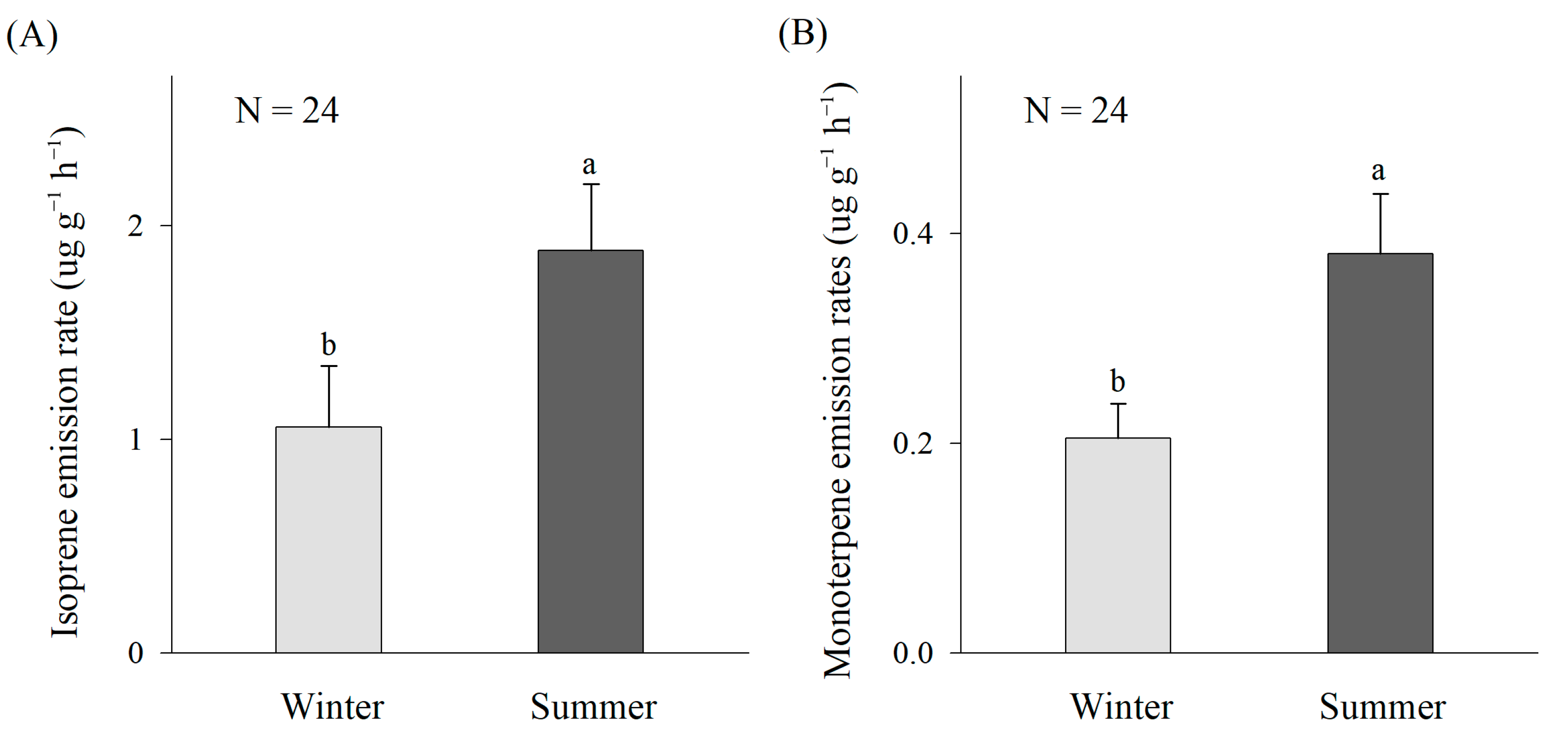
3.1. Seasonal Variations in Isoprene and Monoterpene Emission Rates
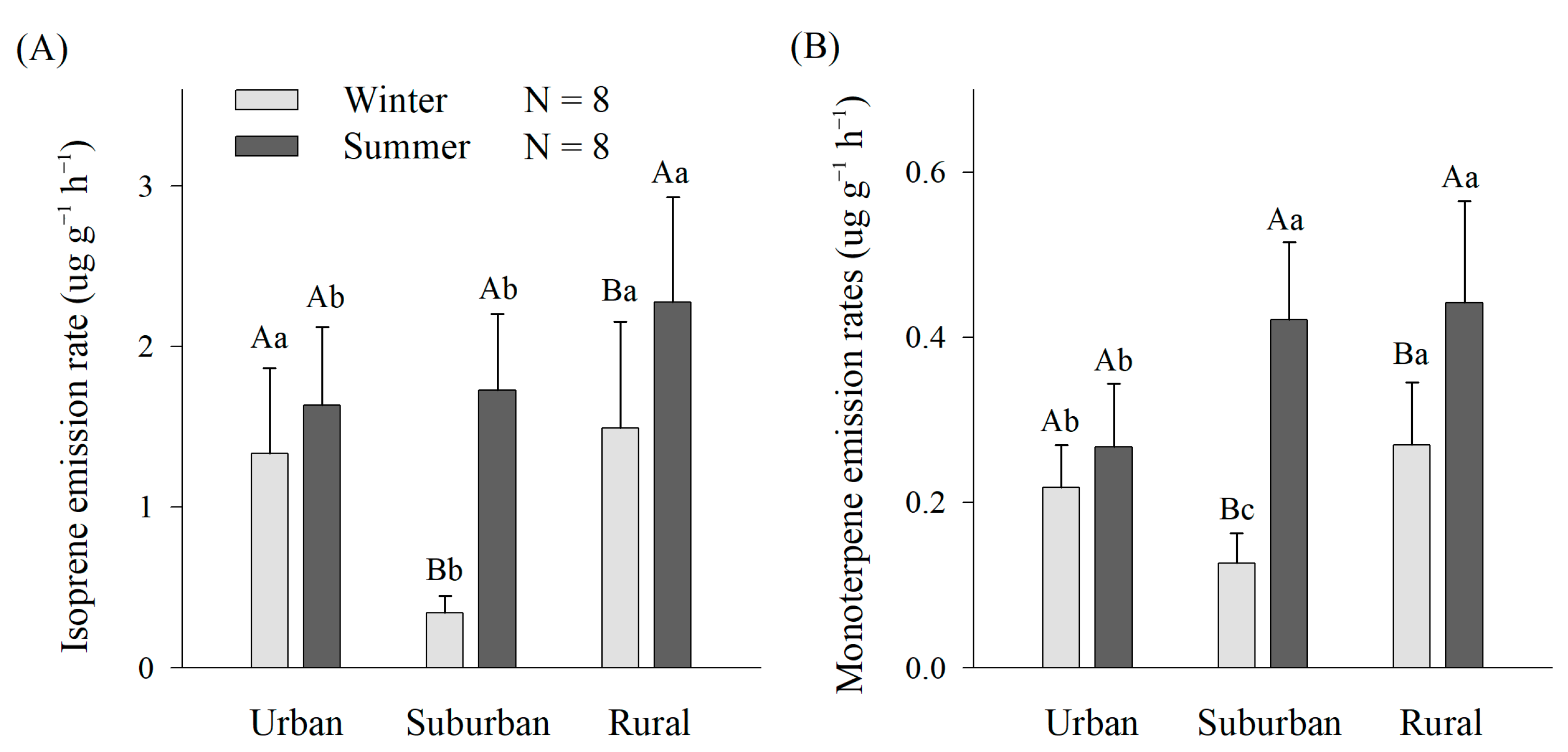
3.2. Differences of Isoprene and Monoterpene Emissions in Urban, Suburban and Rural Environments
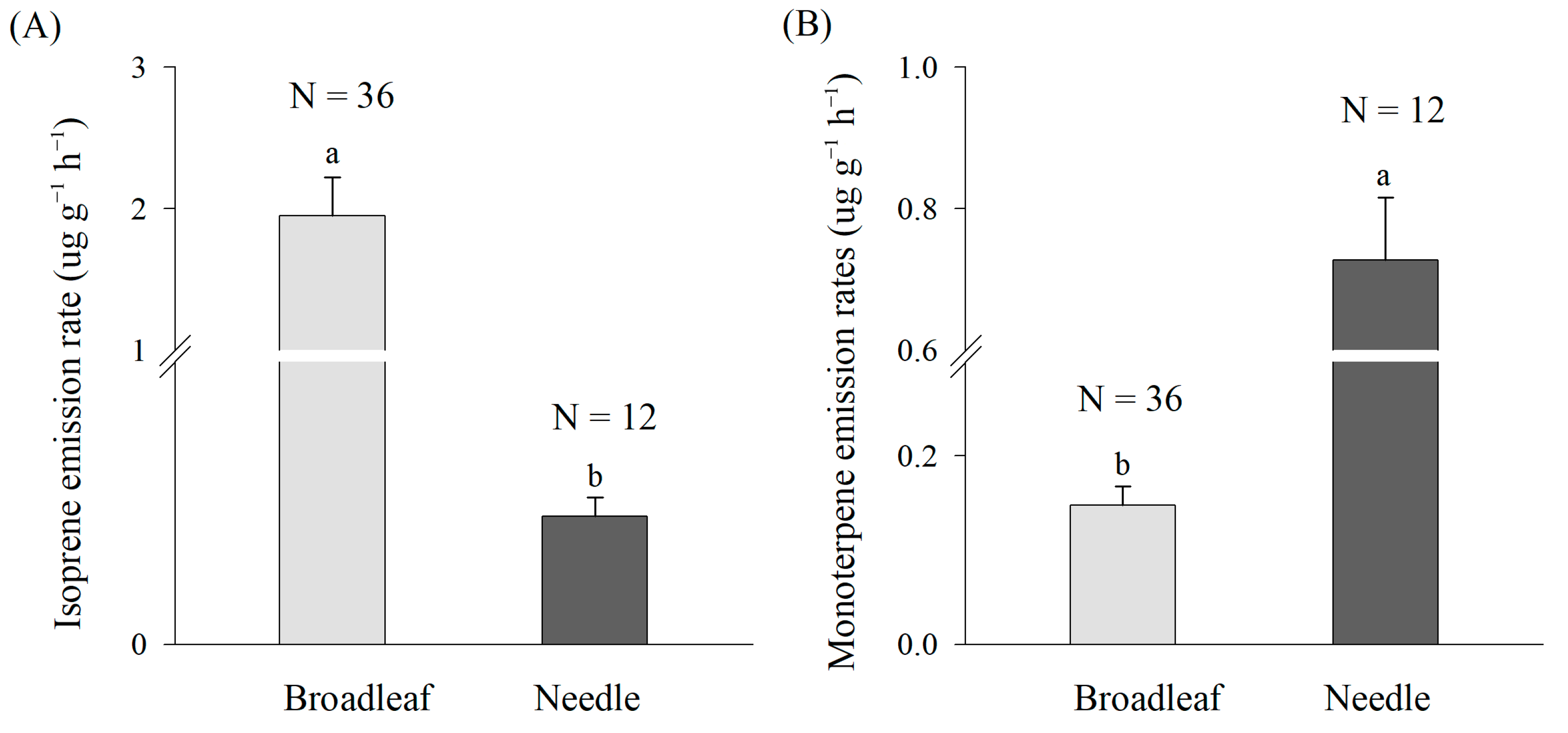
3.3. Differences of Isoprene and Monoterpene Emissions between Broadleaf and Needle
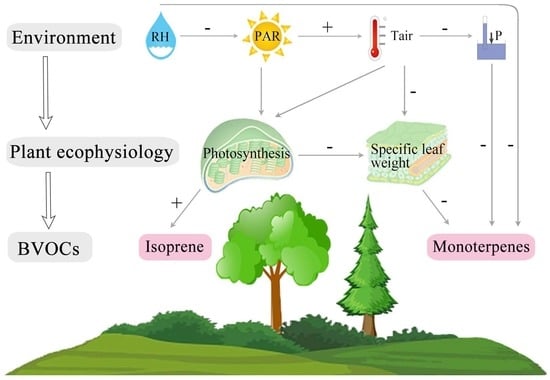
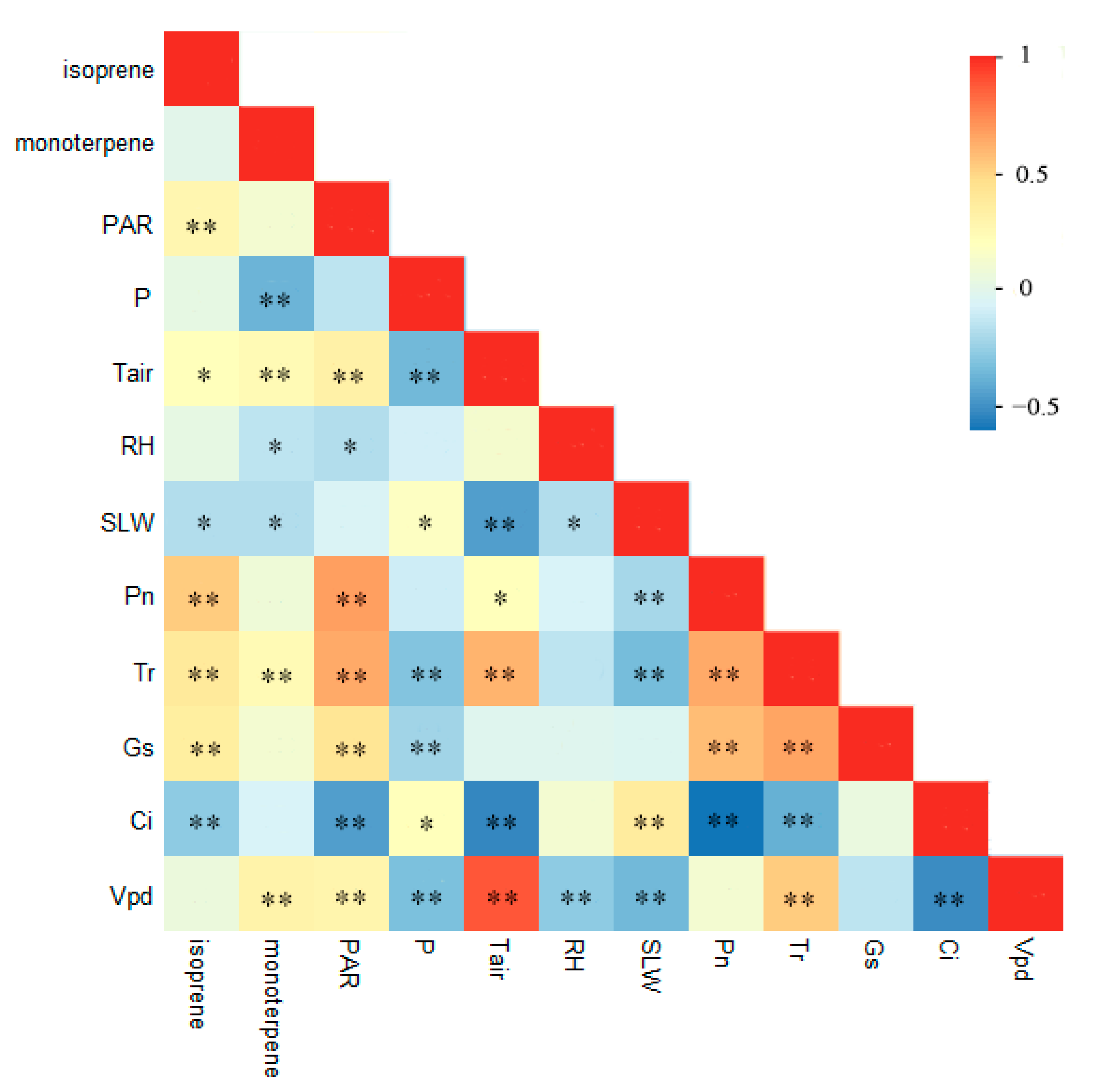
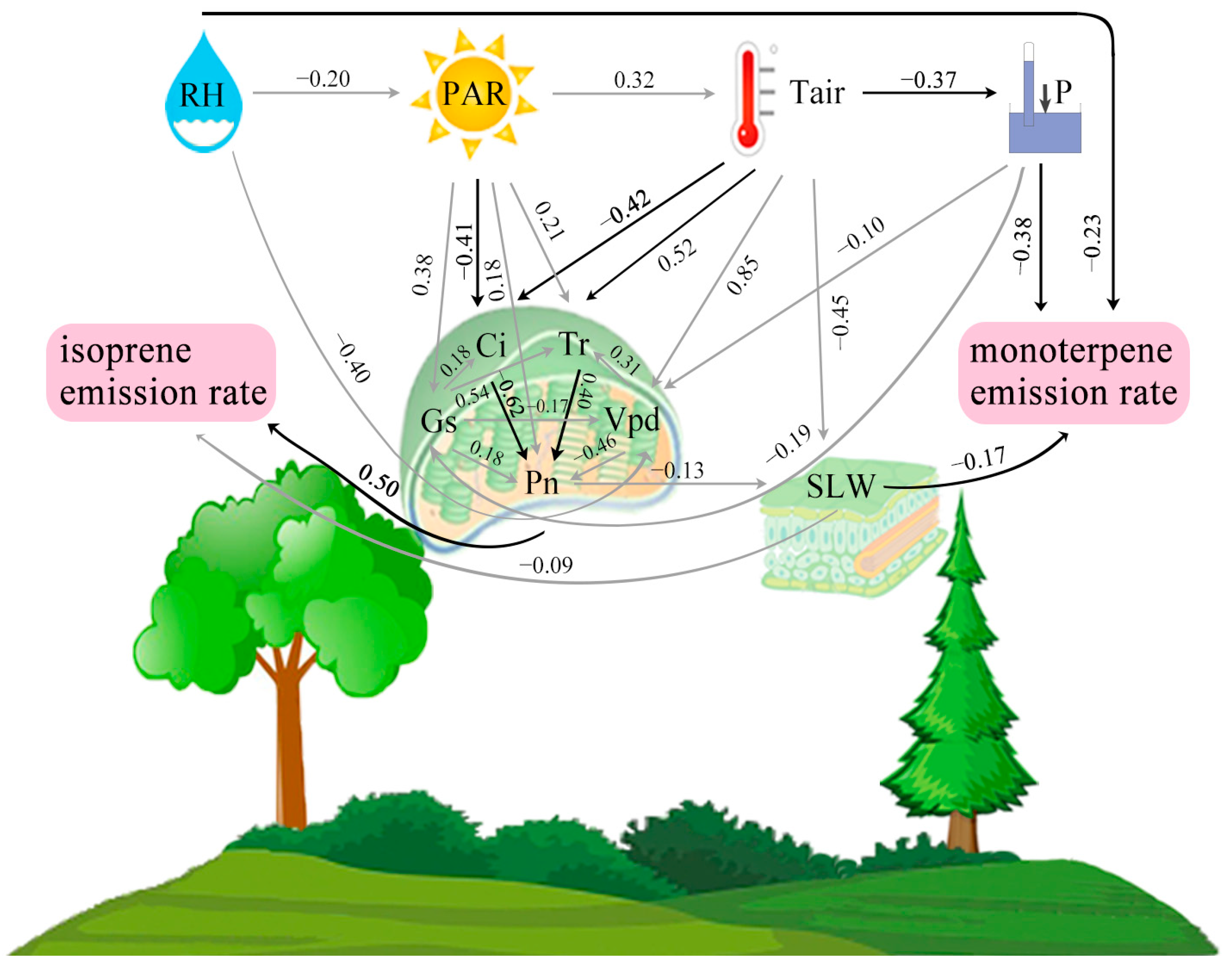
3.4. PA Model of Environment–Plant and Ecophysiology–BVOC Emissions Interaction
4. Discussion
4.1. BVOC Emission Characteristics
4.2. Direct and Indirect Effects of Environmental Factors and Plant Ecophysiology on Isoprene Emissions
4.3. Interaction among Environmental Factors, Plant Ecophysiology and Monoterpene Emissions
4.4. Implications of the PA Model on BVOC Emissions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Messina, P.; Lathière, J.; Sindelarova, K.; Vuichard, N.; Granier, C.; Ghattas, J.; Cozic, A.; Hauglustaine, D.A. Global biogenic volatile organic compound emissions in the ORCHIDEE and MEGAN models and sensitivity to key parameters. Atmos. Chem. Phys. 2016, 16, 14169–14202. [Google Scholar] [CrossRef] [Green Version]
- Duan, C.; Liao, H.; Wang, K.; Ren, Y. The research hotspots and trends of volatile organic compound emissions from anthropogenic and natural sources: A systematic quantitative review. Environ. Res. 2022, 216, 114386. [Google Scholar] [CrossRef] [PubMed]
- Atkinsona, R.; Arey, J. Gas-phase tropospheric chemistry of biogenic volatile organic compounds: A review. Atmos. Res. 2003, 37, 197–219. [Google Scholar] [CrossRef]
- Sindelarova, K.; Markova, J.; Simpson, D.; Huszar, P.; Karlicky, J.; Darras, S.; Granier, C. High-resolution biogenic global emission inventory for the time period 2000–2019 for air quality modelling. Earth Syst. Sci. Data 2022, 14, 251–270. [Google Scholar] [CrossRef]
- Wu, K.; Yang, X.; Chen, D.; Gu, S.; Lu, Y.; Jiang, Q.; Wang, K.; Ou, Y.; Qian, Y.; Shao, P.; et al. Estimation of biogenic VOC emissions and their corresponding impact on ozone and secondary organic aerosol formation in China. Atmos. Res. 2020, 231, 104656. [Google Scholar] [CrossRef]
- Yang, W.; Cao, J.; Wu, Y.; Kong, F.; Li, L. Review on plant terpenoid emissions worldwide and in China. Sci. Total Environ. 2021, 787, 147454. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Wang, X.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.; Chen, A.; Ciais, P.; Tømmervik, H.; et al. Characteristics, drivers and feedbacks of global greening. Earth Environ. 2019, 1, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Gu, S.; Guenther, A.; Faiola, C. Effects of anthropogenic and biogenic volatile organic compounds on Los Angeles air quality. Environ. Sci. Technol. 2021, 55, 12191–12201. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Q.; Guenther, A.; Yang, X.; Wang, L.; Xiao, T.; Li, J.; Feng, J.; Xu, Q.; Cheng, H. Land cover change dominates decadal trends of biogenic volatile organic compound (BVOC) emission in China. Atmos. Chem. Phys. 2020, preprint. [Google Scholar]
- Simon, E.; Kuhn, U.; Rottenberger, S.; Meixner, F.; Kesselmeier, J. Coupling isoprene and monoterpene emissions from Amazonian tree species with physiological and environmental parameters using a neural network approach. Plant Cell Environ. 2005, 28, 287–301. [Google Scholar] [CrossRef]
- Dindorf, T.; Kuhn, U.; Ganzeveld, L.; Schebeske, G.; Ciccioli, P.; Holzke, C.; Koble, R.; Seufert, G.; Kesselmeier, J. Significant light and temperature dependent monoterpene emissions from European beech (Fagus sylvatica L.) and their potential impact on the European volatile organic compound budget. J. Geophys. Res. 2006, 111, D16305. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, A.; Duan, Q.; Jansen, K.; Verena Junker, L.; Kammerer, B.; Rennenberg, H.; Ensminger, I.; Gessler, A.; Kreuzwieser, J. Drought effects on root and needle terpenoid content of a coastal and an interior Douglas fir provenance. Tree Physiol. 2017, 37, 1648–1658. [Google Scholar] [CrossRef] [Green Version]
- Tiiva, P.; Haikio, E.; Kasurinen, A. Impact of warming, moderate nitrogen addition and bark herbivory on BVOC emissions and growth of Scots pine (Pinus sylvestris L.) seedlings. Tree Physiol. 2018, 38, 1461–1475. [Google Scholar] [CrossRef]
- Simin, T.; Tang, J.; Holst, T.; Rinnan, R. Volatile organic compound emission in tundra shrubs—Dependence on species characteristics and the near-surface environment. Environ. Exp. Bot. 2021, 184, 104387. [Google Scholar] [CrossRef]
- Pikkarainen, L.; Nissinen, K.; Ghimire, R.; Kivimaenpaa, M.; Ikonen, V.; Kilpelainen, A.; Virjamo, V.; Yu, H.; Kirsikka-Aho, S.; Salminen, T.; et al. Responses in growth and emissions of biogenic volatile organic compounds in Scots pine, Norway spruce and silver birch seedlings to different warming treatments in a controlled field experiment. Sci. Total Environ. 2022, 821, 153277. [Google Scholar] [CrossRef] [PubMed]
- Kivimäenpää, M.; Riikonen, J.; Valolahti, H.; Elina, H.; Holopainen, J.; Holopainen, T. Effects of elevated ozone and warming on terpenoid emissions and concentrations of Norway spruce depend on needle phenology and age. Tree Physiol. 2022, 42, 1570–1586. [Google Scholar] [CrossRef]
- Rinnan, R.; Iversen, L.; Tang, J.; Vedel-Petersen, I.; Schollert, M.; Schurgers, G. Separating direct and indirect effects of rising temperatures on biogenic volatile emissions in the Arctic. Proc. Natl. Acad. Sci. USA 2020, 117, 32476–32483. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, Z.; Kännaste, A.; Guo, M.; Zhou, G.; Niinemets, Ü. Isoprenoid and aromatic compound emissions in relation to leaf structure, plant growth form and species ecology in 45 East-Asian urban subtropical woody species. Urban For. Urban Green. 2020, 53, 126705. [Google Scholar] [CrossRef]
- Chen, J.; Tang, J.; Yu, X. Environmental and physiological controls on diurnal and seasonal patterns of biogenic volatile organic compound emissions from five dominant woody species under field conditions. Environ. Pollut. 2020, 259, 113955. [Google Scholar] [CrossRef]
- Lahr, E.; Schade, G.; Crossett, C.; Watson, M. Photosynthesis and isoprene emission from trees along an urban-rural gradient in Texas. Glob. Chang. Biol. 2015, 21, 4221–4236. [Google Scholar] [CrossRef] [PubMed]
- Douma, J.C.; Shipley, B. A multigroup extension to piecewise path analysis. Ecosphere 2021, 12, e03502. [Google Scholar] [CrossRef]
- Ren, Y.; Deng, L.; Zuo, S.; Luo, Y.; Shao, G.; Hua, L.; Yang, Y. Geographical modeling of spatial interaction between human activity and forest connectivity in an urban landscape of southeast China. Landsc. Ecol. 2014, 29, 1741–1758. [Google Scholar] [CrossRef]
- Zhou, M.; He, D.; Qin, D.; You, W.; Wu, L.; Xiao, S. Impacts of land use change on vegetation coverage in Xiamen City from 1995 to 2015. J. For. Environ. 2017, 37, 440–445. [Google Scholar]
- Zhao, W.; Li, A.; Huang, Q.; Gao, Y.; Li, F.; Zhang, L. An improved method for assessing vegetation cooling service in regulating thermal environment: A case study in Xiamen, China. Ecol. Indic. 2019, 98, 531–542. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, Y.; Zhang, H.; Song, W.; Wu, Z.; Wang, X. Design and characterization of a semi-open dynamic chamber for measuring biogenic volatile organic compound (BVOC) emissions from plants. Atmos. Meas. Tech. 2022, 15, 79–93. [Google Scholar] [CrossRef]
- Jing, X.; Lun, X.; Fan, C.; Ma, W. Emission patterns of biogenic volatile organic compounds from dominant forest species in Beijing, China. J. Environ. Sci. 2020, 95, 73–81. [Google Scholar] [CrossRef]
- Ortega, J.; Helmig, D.; Daly, R.W.; Tanner, D.M.; Guenther, A.B.; Herrick, J.D. Approaches for quantifying reactive and low-volatility biogenic organic compound emissions by vegetation enclosure techniques—Part B: Applications. Chemosphere 2008, 72, 365–380. [Google Scholar] [CrossRef] [Green Version]
- Ortega, J.; Helmig, D. Approaches for quantifying reactive and low-volatility biogenic organic compound emissions by vegetation enclosure techniques—Part A. Chemosphere 2008, 72, 343–364. [Google Scholar] [CrossRef]
- Karlsson, T.; Rinnan, R.; Holst, T. Variability of BVOC emissions from commercially used willow (Salix spp.) varieties. Atmosphere 2020, 11, 356. [Google Scholar] [CrossRef] [Green Version]
- Mermet, K.; Sauvage, S.; Dusanter, S.; Salameh, T.; Léonardis, T.; Flaud, P.-M.; Perraudin, É.; Villenave, É.; Locoge, N. Optimization of a gas chromatographic unit for measuring biogenic volatile organic compounds in ambient air. Atmos. Meas. Tech. 2019, 12, 6153–6171. [Google Scholar] [CrossRef] [Green Version]
- Guenther, A. Isoprene and Monoterpene Emission Rate Variability Model Evaluations and Sensitivity Analyses. J. Geophys. Res. 1993, 98, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Volf, M.; Volfova, T.; Seifert, C.L.; Ludwig, A.; Engelmann, R.; Jorge, L.R.; Richter, R.; Schedl, A.; Weinhold, A.; Wirth, C.; et al. A mosaic of induced and non-induced branches promotes variation in leaf traits, predation and insect herbivore assemblages in canopy trees. Ecol. Lett. 2021, 25, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Ru, C.; Hu, X.; Chen, D.; Wang, W.; Song, T. Heat and drought priming induce tolerance to subsequent heat and drought stress by regulating leaf photosynthesis, root morphology, and antioxidant defense in maize seedlings. Environ. Exp. Bot. 2022, 202, 105010. [Google Scholar] [CrossRef]
- Du, Y.; Du, J.; Liu, X.; Yuan, Z. Multiple-to-multiple path analysis model. PLoS ONE 2021, 16, e0247722. [Google Scholar] [CrossRef]
- Thomas, J.A.; Ditchman, N.; Beedle, R.B. The impact of knowledge, self-efficacy, and stigma on STI testing intention among college students. J. Am. Coll. Health 2020, 70, 1415–1425. [Google Scholar] [CrossRef]
- Alves, E.; Jardine, K.; Tota, J.; Jardine, A.; Yãnez-Serrano, A.; Karl, T.; Tavares, J.; Nelson, B.; Gu, D.; Stavrakou, T.; et al. Seasonality of isoprenoid emissions from a primary rainforest in central Amazonia. Atmos. Chem. Phys. 2016, 16, 3903–3925. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Gamboa, J.; Paton-Walsh, C.; Galbally, I.; Simmons, J.; Guerette, E.-A.; Griffith, A.; Chambers, S.; Williams, A. Seasonal variation of biogenic and anthropogenic VOCs in a semi-urban area near Sydney, Australia. Atmosphere 2020, 12, 47. [Google Scholar] [CrossRef]
- Liu, Y.; Schallhart, S.; Taipale, D.; Tykkä, T.; Räsänen, M.; Merbold, L.; Hellén, H.; Pellikka, P. Seasonal and diurnal variations in biogenic volatile organic compounds in highland and lowland ecosystems in southern Kenya. Atmos. Chem. Phys. 2021, 21, 14761–14787. [Google Scholar] [CrossRef]
- Helmig, D.; Daly, R.; Milford, J.; Guenther, A. Seasonal trends of biogenic terpene emissions. Chemosphere 2013, 93, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Xu, X.; Liao, D.; Liu, T.; Ji, X.; Xu, K.; Liao, C.; Wang, T.; Lin, C.; Chen, J. Measurement report: Effects of anthropogenic emissions and environmental factors on the formation of biogenic secondary organic aerosol (BSOA) in a coastal city of southeastern China. Atmos. Chem. Phys. 2022, 22, 7827–7841. [Google Scholar] [CrossRef]
- Chen, G.; Liu, T.; Ji, X.; Xu, K.; Hong, Y.; Xu, L.; Li, M.; Fan, X.; Chen, Y.; Yang, C.; et al. Source Apportionment of VOCs and O3 Production Sensitivity at Coastal and Inland Sites of Southeast China. Aerosol Air Qual. Res. 2022, 22, 220289. [Google Scholar] [CrossRef]
- Tani, A.; Mochizuki, T. Review: Exchanges of volatile organic compounds between terrestrial ecosystems and the atmosphere. J. Agric. Meteorol. 2021, 77, 66–80. [Google Scholar] [CrossRef]
- Jason, Q.; Demi, G.; John, H.; Ian, E. Monoterpene synthases responsible for the terpene profile of anther glands in Eucalyptus polybractea R.T. Baker (Myrtaceae). Tree Physiol. 2020, 11, 849–864. [Google Scholar]
- Vanhatalo, A.; Ghirardo, A.; Juurola, E.; Schnitzler, J.-P.; Zimmer, I.; Hellén, H.; Hakola, H.; Bäck, J. Long-term dynamics of monoterpene synthase activities, monoterpene storage pools and emissions in boreal Scots pine. Biogeosciences 2018, 15, 5047–5060. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Murray, F.; Tom, L. Monoterpene and isoprene emissions from 15 Eucalyptus species in Australia. Atmos. Environ. 2000, 34, 645–655. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, Y.; Wang, Z.; Zhang, S. Studies on the emission rates of plants VOCs in China. Environ. Sci. 2004, 24, 654–657. [Google Scholar]
- Huang, A.; Li, N.; Guenther, A.; Greenberg, J.; Baker, B.; Graessli, M.; Bai, J. Investigation on emission properties of biogenic VOCs of landscape plants in Shenzhen. Environ. Sci. 2011, 32, 3555–3559. [Google Scholar]
- Yang, D.; Bai, Y.; Li, J.; Pan, N.; Yu, K.; Tang, L.; Peng, L.; Su, X. Study on hydrocarbon compounds from natural source in the Pearl River Delta area. Environ. Sci. 2001, 21, 422–426. [Google Scholar]
- Alves, E.; Harley, P.; Goncalves, J.; Moura, C.; Jardine, K. Effects of light and temperature on isoprene emission at different leaf developmental stages of Eschweilera coriacea in central Amazon. Acta Amaz. 2014, 44, 9–18. [Google Scholar] [CrossRef]
- Bison, J.; Cardoso-Gustavson, P.; De Moraes, R.; Da Silva Pedrosa, G.; Cruz, L.; Freschi, L.; De Souza, S. Volatile organic compounds and nitric oxide as responses of a Brazilian tropical species to ozone: The emission profile of young and mature leaves. Environ. Sci. Pollut. Res. Int. 2018, 25, 3840–3848. [Google Scholar] [CrossRef] [PubMed]
- Mozaffar, A.; Schoon, N.; Bachy, A.; Digrado, A.; Heinesch, B.; Aubinet, M.; Fauconnier, M.; Delaplace, P.; Du Jardin, P.; Amelynck, C. Biogenic volatile organic compound emissions from senescent maize leaves and a comparison with other leaf developmental stages. Atmos. Environ. 2018, 176, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Dudareva, N.; Klempien, A.; Muhlemann, J.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhou, Y.; Zeng, L.; Dong, F.; Mei, X.; Liao, Y.; Watanabe, N.; Yang, Z. Analytical method for metabolites involved in biosynthesis of plant volatile compounds. RSC Adv. 2017, 7, 19363–19372. [Google Scholar] [CrossRef] [Green Version]
- Widhalm, J.; Jaini, R.; Morgan, J.; Dudareva, N. Rethinking how volatiles are released from plant cells. Trends Plant Sci. 2015, 20, 545–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, J.; Xiong, F.; Sun, N.; Li, Y.; Liu, C.; Yin, S. Photosynthesis and related physiological parameters differences affected the isoprene emission rate among 10 typical tree species in subtropical metropolises. Int. J. Environ. Res. Public Health 2021, 18, 954. [Google Scholar] [CrossRef]
- Penuelas, J. Plant VOC emissions: Making use of the unavoidable. Trends Ecol. Evol. 2004, 19, 402–404. [Google Scholar] [CrossRef]
- Kesselmeier, J.; Staudt, M. Biogenic volatile organic compounds (VOC): An overview on emission, physiology and ecology. J. Atmos. Chem. 1999, 33, 23–88. [Google Scholar] [CrossRef]
- Guidolotti, G.; Rey, A.; Medori, M.; Calfapietra, C. Isoprenoids emission in Stipa tenacissima L.: Photosynthetic control and the effect of UV light. Environ. Pollut. 2016, 208, 336–344. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Veromann Jürgenson, L.L.; Niinemets, Ü. Gall- and erineum-forming Eriophyes mites alter photosynthesis and volatile emissions in an infection severity-dependent manner in broad-leaved trees Alnus glutinosa and Tilia cordata. Tree Physiol. 2021, 41, 1122–1142. [Google Scholar] [CrossRef]
- Oku, H.; Iwai, S.; Uehara, M.; Iqbal, A.; Mutanda, I.; Inafuku, M. Growth condition controls on G93 parameters of isoprene emission from tropical trees. J. Plant Res. 2021, 134, 1225–1242. [Google Scholar] [CrossRef] [PubMed]
- Sellers, P.; Berry, J.; Collatz, G.; Field, C.; Hall, F. Canopy reflectance, photosynthesis, and transpiration. III. a reanalysis using improved leaf models and a new canopy integration scheme. Remote Sens. Environ. 1992, 42, 187–216. [Google Scholar] [CrossRef]
- Zhang, N.; Su, X.; Zhang, X.; Yao, X.; Cheng, T.; Zhu, Y.; Cao, W.; Tian, Y. Monitoring daily variation of leaf layer photosynthesis in rice using UAV-based multi-spectral imagery and a light response curve model. Agric. For. Meterol. 2020, 291, 108098. [Google Scholar] [CrossRef]
- Kanazawa, A.; Chattopadhyay, A.; Kuhlgert, S.; Tuitupou, H.; Maiti, T.; Kramer, D. Light potentials of photosynthetic energy storage in the field: What limits the ability to use or dissipate rapidly increased light energy? R. Soc. Open Sci. 2021, 8, 211102. [Google Scholar] [CrossRef]
- Arena, C.; Tsonev, T.; Doneva, D.; De Micco, V.; Michelozzi, M.; Brunetti, C.; Centritto, M.; Fineschi, S.; Velikova, V.; Loreto, F. The effect of light quality on growth, photosynthesis, leaf anatomy and volatile isoprenoids of a monoterpene-emitting herbaceous species (Solanum lycopersicum L.) and an isoprene-emitting tree (Platanus orientalis L.). Environ. Exp. Bot. 2016, 130, 122–132. [Google Scholar] [CrossRef]
- Chen, J.; Bi, H.; Yu, X.; Fu, Y.; Liao, W. Influence of physiological and environmental factors on the diurnal variation in emissions of biogenic volatile compounds from Pinus tabuliformis. J. Environ. Sci. 2019, 81, 102–118. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Terry, N. Limiting factors in photosynthesis. Plant Physiol. 1984, 75, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernusak, L.; Cheesman, A. The benefits of recycling how photosynthetic bark can increase drought tolerance. New Phytol. 2015, 208, 995–997. [Google Scholar] [CrossRef] [PubMed]
- Slot, M.; Winter, K. In situ temperature relationships of biochemical and stomatal controls of photosynthesis in four lowland tropical tree species. Plant Cell Environ. 2017, 40, 3055–3068. [Google Scholar] [CrossRef]
- Lerdau, M.; Guenther, A.; Monson, R. Plant production and emission of volatile organic compounds. BioScience 1997, 47, 373–383. [Google Scholar] [CrossRef]
- Sulaiman, H.; Liu, B.; Kaurilind, E.; Niinemets, Ü. Phloem-feeding insect infestation antagonizes volatile organic compound emissions and enhances heat stress recovery of photosynthesis in Origanum vulgare. Environ. Exp. Bot. 2021, 189, 104551. [Google Scholar] [CrossRef]
- Guidolotti, G.; Pallozzi, E.; Gavrichkova, O.; Scartazza, A.; Mattioni, M.; Loreto, F.; Calfapietra, C. Emission of constitutive isoprene, induced monoterpenes, and other volatiles under high temperatures in Eucalyptus camaldulensis: A (13) C labelling study. Plant Cell Environ. 2019, 42, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Fasbender, L.; Romek, K.; Yáñez-Serrano, A.; Kreuzwieser, J. Heat waves change plant carbon allocation among primary and secondary metabolism altering CO2 assimilation, respiration, and VOC emissions. Front. Plant Sci. 2020, 11, 1242. [Google Scholar] [CrossRef] [PubMed]
- Riedlmeier, M.; Ghirardo, A.; Wenig, M.; Knappe, C.; Koch, K.; Georgii, E.; Dey, S.; Parker, J.; Schnitzler, J.; Vlot, A. Monoterpenes support systemic acquired resistance within and between plants. Plant Cell 2017, 29, 1440–1459. [Google Scholar] [CrossRef] [Green Version]
- Tiiva, P.; Tang, J.; Michelsen, A.; Rinnan, R. Monoterpene emissions in response to long-term night-time warming, elevated CO2 and extended summer drought in a temperate heath ecosystem. Sci. Total Environ. 2017, 580, 1056–1067. [Google Scholar] [CrossRef]
- Barbieri, S.; Elustondo, M.; Urbicain, M. Retention of aroma compounds in basil dried with low pressure superheated steam. J. Food Eng. 2004, 65, 109–115. [Google Scholar] [CrossRef]
- Babu, K.; Singh, B.; Joshi, V.; Singh, V. Essential oil composition of Damask rose (Rosa damascena Mill.) distilled under different pressures and temperatures. Flavour Fragr. J. 2002, 17, 136–140. [Google Scholar] [CrossRef]
- Widegren, J.; Bruno, T. Vapor pressure measurements on low-volatility terpenoid compounds by the concatenated gas saturation method. Environ. Sci. Technol. 2010, 44, 388–393. [Google Scholar] [CrossRef]
- Edelmann, H.; Neinhuis, C.; Bargel, H. Influence of hydration and temperature on the rheological properties of plant cuticles and their impact on plant organ integrity. J. Plant Growth Regul. 2005, 24, 116–126. [Google Scholar] [CrossRef]
- Ma, F.; Barrett-Lennard, E.; Tian, C. Changes in cell size and tissue hydration (‘succulence’) cause curvilinear growth responses to salinity and watering treatments in euhalophytes. Environ. Exp. Bot. 2019, 159, 87–94. [Google Scholar] [CrossRef]
- Duan, C.; Zuo, S.; Wu, Z.; Qiu, Y.; Wang, J.; Lei, Y.; Liao, H.; Ren, Y. A review of research hotspots and trends in biogenic volatile organic compounds (BVOCs) emissions combining bibliometrics with evolution tree methods. Environ. Res. Lett. 2020, 16, 013003. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Meischner, M.; Grun, M.; Yanez-Serrano, A.; Fasbender, L.; Werner, C. Drought affects carbon partitioning into volatile organic compound biosynthesis in Scots pine needles. New Phytol. 2021, 232, 17736. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, M.; Llusià, J.; Filella, I.; Niinemets, Ü.; Arneth, A.; Wright, I.; Loreto, F.; Peñuelas, J. Nutrient-rich plants emit a less intense blend of volatile isoprenoids. New Phytol. 2018, 220, 773–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, P.; Thompson, K.; Hodgson, J. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Ibrahim, M.; Maenpaa, M.; Hassinen, V.; Kontunen-Soppela, S.; Malec, L.; Rousi, M.; Pietikainen, L.; Tervahauta, A.; Karenlampi, S.; Holopainen, J.; et al. Elevation of night-time temperature increases terpenoid emissions from Betula pendula and Populus tremula. J. Exp. Bot. 2010, 61, 1583–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vescio, R.; Abenavoli, M.; Araniti, F.; Musarella, C.; Sofo, A.; Laface, V.; Spampinato, G.; Sorgonà, A. The assessment and the within-plant variation of the morpho-physiological traits and VOCs profile in endemic and rare salvia ceratophylloides Ard. (Lamiaceae). Plants 2021, 10, 474. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Hauff, K.; Bertin, N.; Tenhunen, J.; Steinbrecher, R.; Seufert, G. Monoterpene emissions in relation to foliar photosynthetic and structural variables in Mediterranean evergreen Quercus species. New Phytol. 2001, 153, 243–356. [Google Scholar] [CrossRef]
- Liao, P.; Ray, S.; Boachon, B.; Lynch, J.; Deshpande, A.; McAdam, S.; Morgan, J.; Dudareva, N. Cuticle thickness affects dynamics of volatile emission from petunia flowers. Nat. Chem. Biol. 2021, 17, 138–145. [Google Scholar] [CrossRef]
- Geron, C.; Arnts, R. Seasonal monoterpene and sesquiterpene emissions from Pinus taeda and Pinus virginiana. Atmos. Environ. 2010, 44, 4240–4251. [Google Scholar] [CrossRef]
- Tian, Z.; Luo, Q.; Zuo, Z. Seasonal emission of monoterpenes from four chemotypes of Cinnamomum camphora. Ind. Crops Prod. 2021, 163, 113327. [Google Scholar] [CrossRef]
- Kuhn, U.; Rottenberger, S.; Biesenthal, T.; Wolf, A.; Schebeske, G.; Ciccioli, P.; Brancaleoni, E.; Frattoni, M.; Tavares, T.; Kesselmeier, J. Seasonal differences in isoprene and light-dependent monoterpene emission by Amazonian tree species. Glob. Change Biol. 2004, 10, 663–682. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Seufert, G.; Steinbrecher, R.; Tenhunen, J. A model coupling foliar monoterpene emissions to leaf photosynthetic characteristics in Mediterranean evergreen Quercus species. New Phytol. 2001, 153, 257–275. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, C.; Wu, Z.; Liao, H.; Ren, Y. Interaction Processes of Environment and Plant Ecophysiology with BVOC Emissions from Dominant Greening Trees. Forests 2023, 14, 523. https://doi.org/10.3390/f14030523
Duan C, Wu Z, Liao H, Ren Y. Interaction Processes of Environment and Plant Ecophysiology with BVOC Emissions from Dominant Greening Trees. Forests. 2023; 14(3):523. https://doi.org/10.3390/f14030523
Chicago/Turabian StyleDuan, Chensong, Zhifeng Wu, Hu Liao, and Yin Ren. 2023. "Interaction Processes of Environment and Plant Ecophysiology with BVOC Emissions from Dominant Greening Trees" Forests 14, no. 3: 523. https://doi.org/10.3390/f14030523
APA StyleDuan, C., Wu, Z., Liao, H., & Ren, Y. (2023). Interaction Processes of Environment and Plant Ecophysiology with BVOC Emissions from Dominant Greening Trees. Forests, 14(3), 523. https://doi.org/10.3390/f14030523