Abstract
Soil macrofauna is an important component of soil biodiversity and plays a key role in soil ecosystem function. Little work, however, has explored how macrofauna may contribute, directly or indirectly, to soil microfauna communities. In this study, we explored the impact of two classes of common soil macrofauna, earthworms (Eisenia foetida) and millipedes (Spirobolus walkeri), on soil nematode community diversity and structure. We found that earthworms and millipedes play an important role in maintaining soil nematode communities and increasing soil nutrients. Both earthworms and millipedes act as hosts for soil nematodes, and play a valuable role in dispersing and distributing these important microfauna throughout the soil landscape. We tested the efficacy of this strategy on multiple soil layers and found that layer does not have a significant influence on soil nematode community. These results suggest that soil nematode abundance, diversity, and structure are tightly linked to the existence of soil macrofauna, which may contribute to the maintenance mechanism of forest soil biodiversity.
1. Introduction
Greater soil biodiversity helps to maintain ecosystem multifunction [1,2]. A wide variety of soil fauna (e.g., macrofauna and microfauna) contribute to this biodiversity and are key regulators of soil biogeochemical cycles [3]. Soil macrofauna can directly affect soil structure and physicochemical properties through their lifetimes, mainly by feeding, moving, and excreting. Similarly, soil microfauna prey upon soil microbes, leading to microbial turnover and, potentially, shifts in community composition [4,5]. Soil macro- and microfauna also play an irreplaceable role in the mineralization of soil organic matter [6,7,8,9]. Despite a universal understanding that soil fauna are important players underlying a host of soil processes, the role they play in mediating forest soil biodiversity is unexplored and poorly described.
Earthworms and millipedes are typical and common soil macrofauna in forest ecosystems [10,11]. Both play an important role in leaf litter decomposition and nutrient cycling [12]. Earthworms live mainly in the soil and survive by feeding on soil organic matter and plant residues. Earthworms digging in the soil layer can change the soil structure and air and water state, increase available nutrients in the soil, and promote nutrient mineralization [13]. Millipedes live on the soil surface and mainly feed on leaf litter, accelerating decomposition and indirectly providing a nutrient source for the growth of other organisms [14]. Considering that these two macrofauna have distinct mechanical interfaces with the soil environment, we anticipate that earthworms and millipedes may have understandably different contributions to forest soil biodiversity.
Soil nematodes are one of the most abundant and widely distributed soil microfauna in terrestrial ecosystems, and have multiple trophic positions in the soil food web [15]. Due to the close connection of soil nematodes to the soil environment and their sensitivity to environmental disturbances, soil nematode communities are often used as bioindicators of the changes in the structure and function of soil food webs [16,17,18]. The diversity and community composition of nematode communities can be important indicators of soil biodiversity and nutrient status [19].
Some evidence suggests that these important soil nematode communities may be regulated by important soil macrofauna [20]. For example, recent work has demonstrated that a decrease in nematode abundance is often associated with increases in the number and biomass of earthworms [21,22]. Similarly, Domínguez et al. [23] found that earthworm activity significantly reduced the abundance of nematodes (by more than 50%) in fresh organic waste. Other work has suggested that earthworms may indirectly regulate nematode communities. For example, earthworm mucus residue has been shown to affect the behavior and physiology of bacterivorous nematodes [24]. However, these studies are often contradicting. Senapati [25], for example, found that earthworms (largely through their high-nitrogen excretion material, mucus, and dead earthworm organization) may be responsible for the stimulation of the growth of the nematode population.
Still more work has suggested that the increased number of soil nematode individuals may be related to the intestinal transmission of nematode parasites within earthworms. For instance, studies have shown that earthworms transport nematodes through the gut harmlessly, and that digestion does not affect the hatching of the nematode eggs, nor does it kill the larvae or the adults [26]. This survival and gut transport may indicate a possible mechanism by which macrofauna maintains soil nematode communities. For example, Shapiro et al. [27] speculated that the spread of nematodes in the soil may be increased by body surface or the digestive tract transport of earthworms. Some of these hypotheses have been borne out, and Villenave et al. [28] observed that when earthworms were artificially ‘inoculated’ with nematodes in the field, the overall density of nematodes on-site increased, especially fungivorous and bacterivorous nematodes.
Although there are few relevant studies on the relationship between millipedes and nematodes [29], nematodes can obtain food that has been ingested and digested by millipede hosts without significant harm to millipedes [30]. Nematodes can also survive in the guts of millipedes, and are thought to play an important role in regulating intestinal bacteria [29]. In the millipede gut, nematodes can promote nutrient cycling and are important for the overall health of the gut microbiome [29]. Though less work has been dedicated to understanding how millipede populations may impact and regulate soil nematode communities, they are a useful contrast to earthworms and are valuable to explore. Both earthworms and millipedes may also indirectly promote nematode community health by increasing soil nutrient availability [25,31,32].
Biodiversity restoration in the forest is usually carried out aboveground; however, direct and effective methods for restoring belowground biodiversity remain poorly studied. In addition, our knowledge regarding the maintenance mechanism of forest soil biodiversity is limited by the complicated interaction effects between diverse soil communities. Soil macrofauna is known as a group of ecosystem engineers, and plays an important role in soil processes. It is necessary to better understand how soil macrofauna inoculation affects forest soil biodiversity. To that end, we set up a microcosm experiment studying the effects of the soil macrofauna (earthworms and millipedes) and their interactions with soil nematodes. We hypothesized that: (1) both earthworm addition and millipede addition could increase the abundance of soil nematodes; and (2) simultaneous earthworm addition and millipede addition would lead to the greatest biodiversity in the soil nematode community.
2. Materials and Methods
2.1. Study Site and Experimental Materials
The microcosm experiment was conducted in the laboratory of the South China Botanical Garden, Chinese Academy of Sciences, Guangzhou (23°13′ N, 113°28′ E), Guangdong Province, China. The region has a subtropical monsoon climate, with an annual average temperature of 23 °C and an annual average rainfall of 1623.6–1899.8 mm. The millipede Spirobolus walkeri was collected in a Juglans regia orchard in Tianjin, China (40°05′ N, 117°41′ E) on 26 September 2021. On 10 October 2021, Eisenia foetida (a common endogeic earthworm) were purchased from a farm in Guangzhou and brought back to the laboratory. On 16 October 2021, we collected a 0–10 cm layer of soil from a Dimocarpus longan forest in the South China Botanical Garden, sifted these soils through a 2 mm sieve, picked out gravel, roots, leaves, and soil animals, and then re-homogenized our soils to create a well-mixed soil system devoid of macrofauna competitors and large physical obstructions.
2.2. Experiment Design
We approached the experiment using a four-part design: (1) a control treatment without soil macrofauna (CK), (2) an earthworm addition (E), (3) a millipede addition (M), and (4) a simultaneous earthworm addition and millipede addition (E + M). Each treatment was replicated 5 times for a total of 20 microcosms. Each microcosm was constructed from a single PVC cylinder 15 cm in diameter and 20 cm in height. A sample of 1800 g of fresh soil was sterilized using an autoclave pot (126 °C, 0.142 MPa, 40 min), then added to each microcosm until the soil columns were approximately 10 cm in length. These soils were then ‘topped off’ with 13 g of dried and autoclaved litter leaves spread evenly on the soil surface. Once the microcosm was established, we added our soil animals. Considering the survival rate and necessary living space of animals, 6 earthworms were added for the earthworm addition treatment, 3 millipedes were added for the millipede addition treatment, and 6 earthworms and 3 millipedes were added for the simultaneous addition treatment. We then covered the PVC column with 3 layers of 2 mm aperture nylon mesh to prevent our macrofauna from escaping. During the incubation period, the room temperature was kept between 25–28 °C, the air humidity was 40%–60%, and small and equal amounts of deionized water were regularly sprayed into all microcosms to maintain surface soil moisture. The experiment ran for just over 1 month, beginning on 19 October 2021 and ending on 30 November, for a total of 6 weeks.
2.3. Sampling and Analysis
At the end of the incubation, the remaining litter was collected from the soil surface with tweezers. Then, part of the litter was extracted by the Baermann funnel method and ultimately fixed with 4% formaldehyde [33]. The litter used for nematode extraction was dried and weighed to calculate the abundance of nematodes. While collecting litter, millipedes were collected from each PVC column and each placed into a 50 mL centrifuge tube. We then used a spoon to directly collect earthworm feces and millipede fecal pellets on the soil surface in the PVC soil column. We collected fecal pellets to directly measure parasite load. Remaining casts and fecal pellets were weighed and dried to measure the water content; thus, we were able to scale our one-pellet measurements to both the individual and community levels. After collecting all the litter and animal casts on the soil surface, we sampled the soil directly. We collected samples from both the 0–5 cm and 5–10 cm layers, carefully picking out the surviving earthworms and placing them into a 50 mL centrifuge tube. Collected soils in each layer were mixed and divided into two homogenous subsamples. One was used to determine water content and extract soil nematodes; the other was dried in a cool and ventilated place to determine other physiochemical properties of the soil. All of the millipedes and most of the earthworms added were alive.
To collect soil nematodes from the body surface, 50 mL deionized water was added into a 50 mL centrifuge tube containing earthworms or millipedes, placed on a shaking table (200 rpm), and vibrated for 10 min. After this homogenization, we removed visible earthworms and millipedes with tweezers, and let the remaining suspension settle for 2 h. After the suspension had settled, we removed and discarded the supernatant, thus retaining about 8 mL of liquid, to which we added 95% alcohol of the same volume to artificially fix all samples to a 16 mL volume.
To collect nematodes from the gut, earthworms and millipedes were removed from the soil, dissected and chopped, loaded into another 50 mL centrifuge tube with deionized water, and placed onto a shaking table (200 rpm) for 10 min. Afterwards, we removed the animal body debris and left the remaining suspension to settle. After settling, we discarded the supernatant, retained about 8 mL, and added 95% equal volume of alcohol to raise the total volume to 16 mL.
All collected nematodes were counted with an inverted microscope and identified to the genus level. According to their feeding habits, they were divided into bacterivores (Ba), fungivores, herbivores, and omnivore predators [34]. Nematode functional guilds were determined according to their colonizer–persister (c-p) value [35] (http://nemaplex.ucdavis.edu/Ecology/EcophysiologyParms/GenusParmsQuery.aspx (accessed on 23 May 2022)). For instance, the Ba1 guild is a group of r-strategy bacterivorous nematodes with a c-p value of 1. The first 100 nematodes were identified for each sample; if there were fewer than 100 individuals in the sample, all nematodes were identified.
We used biomass measurements before and after a drying oven treatment (105 °C, 48 h) to measure soil water content (SWC), and also used a pH meter to measure the soil solution, with the soil water ratio of 1:2.5 (w/v). All samples also used the element analyzer to measure both soil organic carbon (SOC) and soil total nitrogen (STN).
2.4. Statistical Analysis
The abundance of soil nematodes was calculated based on the number of nematodes in soil, litter, or feces per 100 g dry weight. Three-way ANOVAs were used to test the effects of earthworm addition, millipede addition, and soil layer on the soil properties and the abundance of nematodes in the soil. Two-way ANOVAs were used to test the effects of earthworm addition and millipede addition on the abundance of nematodes in the litter. Post hoc LSD tests were used to compare the differences across treatments. One-way ANOVAs were used to compare the abundance of nematodes in control soil, treated soil, and animal feces. A nonmetric multidimensional scaling (NMDS) representing Bray–Curtis dissimilarity was used to examine whether earthworm addition or millipede addition differentially influenced nematode community composition [36]. The significance level was set to p = 0.05.
3. Results
3.1. Soil Properties
Earthworm addition significantly increased the soil nutrient content of SOC (F = 30.266, p < 0.001; Table 1; Figure 1A) and STN (F = 27.263, p < 0.001; Table 1; Figure 1B), while simultaneously increasing pH (F = 31.744, p < 0.001; Table 1; Figure 1C) and SWC (F = 40.931, p < 0.001; Table 1; Figure 1D). Millipede addition also significantly increased SOC (F = 9.313, p = 0.005; Table 1; Figure 1A), and pH (F = 9.580, p = 0.004; Table 1; Figure 1B), as well as STN (F = 49.874, p < 0.001; Table 1; Figure 1C). A significant interactive effect of earthworm addition and soil depth on soil pH was also detected (F = 18.259, p < 0.001; Table 1; Figure 1C). Furthermore, SWC at 5–10 cm depth was significantly higher than that at 0–5 cm (F = 38.377, p < 0.001; Table 1; Figure 1D).

Table 1.
Effects of earthworm addition (E), millipede addition (M), and soil depth (depth) on soil properties, soil nematode abundance, and diversity.
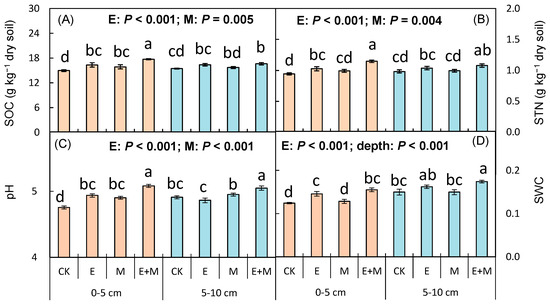
Figure 1.
Soil properties after the addition of earthworms and millipedes to two soil layers. CK, control; E, earthworm addition; M, millipede addition; E + M, earthworm addition + millipede addition. (A) SOC: soil organic carbon (g·kg−1 dry soil); (B) STN: soil total nitrogen (g·kg−1 dry soil); (C) pH: soil pH; (D) SWC: soil water content (ratio of water to dry soil). Data represent means ± SE (n = 5). Significant (p < 0.05) effects based on one-way ANOVA are presented by lowercase letters.
3.2. Nematode Abundances and Species Richness
Ten nematode genera were recorded in the present study, and six of them were bacterivores. No omnivore predators were found in the present study. Due to the low abundances of fungivores and herbivores, only abundances of bacterivores and Ba1 guild were analyzed. The two-way ANOVA results showed no significant effects of earthworm addition or millipede addition on either nematode abundance or species richness in the leaf litter (Table 2; Figure 2A–D). However, earthworm addition significantly increased total nematode abundance (F = 42.515, p < 0.001; Table 1; Figure 2E) in the soil, as well as the abundance of bacterivores (F = 47.880, p < 0.001; Table 1; Figure 2F) and the Ba1 guild (F = 29.823, p < 0.001; Table 1; Figure 2G). Earthworm addition also ultimately increased soil community species richness (F = 30.215, p < 0.001; Table 1; Figure 2H, 5–10 cm). Earthworm addition also increased the abundance of nematodes in both earthworm casts and soil when compared to soil receiving the control treatment (Figure 3A–D). We did not observe any significant difference in nematode abundance or species richness in millipede fecal pellets. These pellets were indistinguishable from the soil receiving the millipede addition treatment. Nematodes were found both on the body surface and in the gut of the earthworms and millipedes. The mean value of the nematode individuals found on the body surfaces and in the guts of the millipedes was higher than that of earthworms (Figure 4).
Table 2.
Effects of earthworm addition (E) and millipede addition (M) on nematodes in leaf litter.
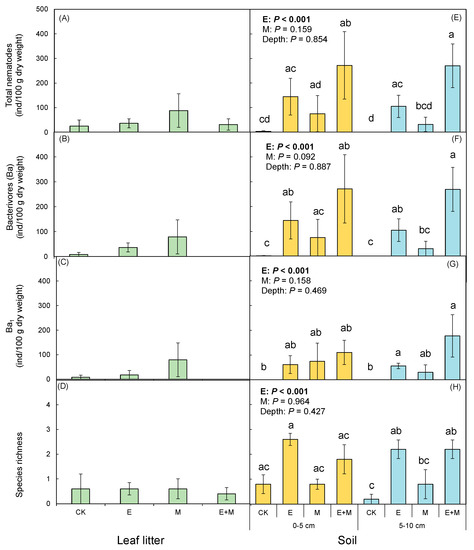
Figure 2.
Abundances and species richness of nematodes in the leaf litter and soil, as affected by earthworms and millipedes. (A,E) Total nematodes abundance; (B,F) the abundance of bacterivores; (C,G) the abundance of Ba1 guild of the bacterivores; (D,H) number of the nematode species. Data represent means ± SE (n = 5). Significant (p < 0.05) effects based on one-way ANOVA are presented by lowercase letters.
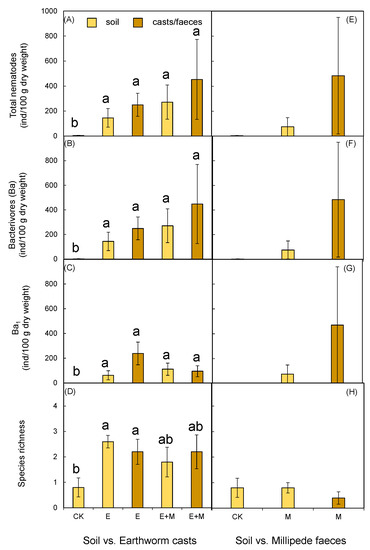
Figure 3.
Comparison of the abundance and species richness of the nematodes between the soil and earthworm casts and the millipede feces. (A,E) Total nematodes abundance; (B,F) the abundance of bacterivores; (C,G) the abundance of Ba1 guild of the bacterivores; (D,H) number of the nematode species. Data represent means ± SE (n = 5). Significant (p < 0.05) effects based on one-way ANOVA are presented by lowercase letters. No millipede feces was found sample receiving E + M treatment.
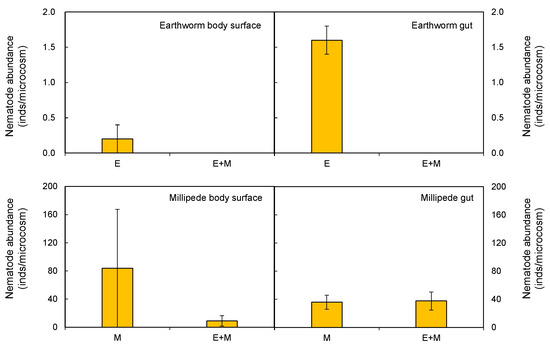
Figure 4.
Abundances of the nematodes on the body surfaces or in the guts of earthworms and millipedes in each microcosm. Data represent means ± SE (n = 5).
3.3. Treatment Effects on Soil Nematode Taxonomic Community Structure
An NMDS ordination based on the distribution of soil samples after macrofauna addition showed strong associations with nematode taxonomic community structure among treatments (df = 3, r2 = 0.27, F= 3.07, p = 0.002; Figure 5). The earthworm addition + millipede addition treatment was associated with a larger number of nematode genera compared to individual earthworm or millipede addition (Figure 5). Meanwhile, the earthworm addition treatment and the millipede addition treatment were both related to bacterivore taxa (Ba1 groups). Specifically, the earthworm addition treatment was more related to Diplogasteridae, while the millipede addition treatment was more related to Rhabditis.
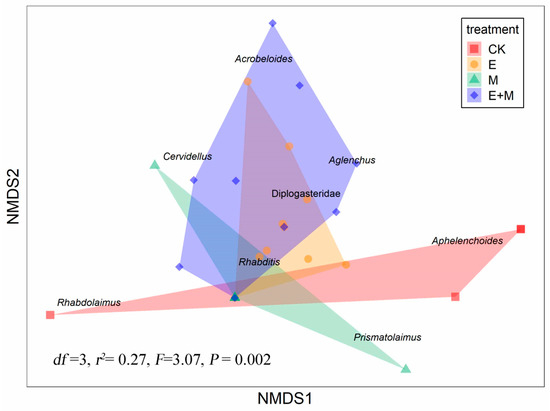
Figure 5.
Nonmetric multidimensional scaling (NMDS) plot of nematode genera/family composition as a function of treatments (Bray–Curtis).
4. Discussion
This study mainly explored the transport of soil nematodes by soil macrofauna in the gut and body surface, as well as the dispersal of nematodes at different interfaces (litter, soil, body surface, and feces) of soil ecosystems. We found that soil macrofauna was able to significantly increase the abundance of soil nematodes, and that the community structures of the nematodes reconstructed in the soil by different macrofauna varied significantly, partly confirming our hypotheses.
How various soil fauna influence each other, through direct and indirect effects, remains a fundamental question in ecology [37]. Previous studies have largely explored the influence of adding a single macrofauna class (earthworm or millipede) to soil nematode communities [26,29]. Fewer studies, however, have focused on the mechanisms of and interactions between soil macrofauna and soil nematode communities, especially in forest ecosystems. In line with our first hypothesis, the addition of earthworms significantly increased the abundance of total nematodes and bacterivorous nematodes in the soil. The possible explanation is that earthworm activities may have spread the nematode individuals and eggs throughout the soil, and then promoted the assemblage process of the soil nematode community [27]. Another possible explanation is that earthworms increased the available nutrients in the soil by accelerating the soil organic matter mineralization. In fact, previous studies have shown that soil nematode abundance was positively correlated with the content of available phosphorus in the soil after earthworm addition [20,38]. In our study, earthworms and millipedes increased the soil nutrients, and may have stimulated soil nematode abundances through bottom-up effects. However, negative effects of earthworms on soil nematodes have also been reported, mainly because of their feeding habits (epigeic, endogeic, or anecic) and substrate conditions [39]. Furthermore, it should be noted that exotic earthworms may cause a decrease in the soil available nutrients and the abundances of soil nematodes compared with native earthworms in subtropical forests [38]. Surprisingly, the combined earthworm and millipede treatment did not produce the most diverse soil nematode community of our experiment, contradicting our second hypothesis. It is possible that this is because inter-individual variation in the nematode community was extremely high. A given millipede may actually be extremely similar to an earthworm, depending on the individual. In this case, the combined treatment may not have been effectively different than treatment with either worms or millipedes.
Despite this variability, we found that earthworms and millipedes promoted distinctly different soil nematode genera assemblage. There are likely two explanations for this difference: promotion and inhibition. Earthworms and millipedes host different varieties of soil nematodes, which they disperse throughout the soil. By propagating these nematodes, they likely shift the direction of the ultimate soil community. Secondly, some studies have shown that earthworms may inhibit the abundance of specific nematodes in soil. For instance, Yu et al. [24] found that the mucus produced by the earthworm body surface has negative effects on the behavior and physiology of some bacterivorous nematodes.
There were no significant differences between the nematode abundances in the 0–5 cm soil layer and those in the 5–10 cm soil layer (Table 1, Figure 2), suggesting that the soil nematode community is more strongly controlled by biotic interactions with macrofauna than by the physical and chemical properties associated with the soil layer. This was an interesting result. Millipedes usually live in the surface layer of the soil, and are thought to only impact decomposition and resource acquisition at the soil surface [40]. Conversely, endogenic earthworms are mostly distributed in the subsoil layer, and their indirect effects on nematodes are usually mediated by changes in soil structure, moisture status, and nutrient cycling [21]. That said, over the course of the experiment, we did observe burrowing millipedes and surface-dwelling earthworms, suggesting that the previously assumed vertical stratification of their impacts may be inaccurate in this setting. In addition, while the 42 days of the experiment may have been a sufficient duration to allow for the diffusion of soil nematodes throughout the mesocosm, it may have been insufficient to allow for the complete development of a ‘top’ and ‘bottom’ community. It is possible that, over a longer experimental period, the abiotic variation in soil layers would have been sufficient to observe two distinct communities. Overall, we found that soil macrofauna inoculation may be an effective means to reconstruct soil communities in defaunated soil, such as severely degraded forest soils. In other words, belowground macrofauna inoculation can be a potential complementary method to aboveground afforestation in the restoration processes of degraded forest ecosystems.
5. Conclusions
In this study, we explored and compared the influences of two classes of soil macrofauna (earthworms and millipedes) on soil nematode community composition and biodiversity. Both earthworms and millipedes demonstrated the potential to carry and spread soil nematodes on the body surface, in the gut, and, ultimately, in feces. This study further advances our understanding of how soil macrofauna and their interactions affect soil nematode community composition, and highlights the important role that soil macrofauna plays in maintaining soil biodiversity and soil nutrients in forest ecosystems.
Author Contributions
Conceptualization, T.L. and X.L.; methodology, T.L.; formal analysis, T.L. and B.Z.; data curation, Z.W., H.G., Y.W. and Y.Q.; writing—original draft preparation, T.L. and Z.W.; writing—review and editing, T.L., X.H., W.Y. and X.L.; visualization, B.Z.; supervision, X.L.; funding acquisition, T.L. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (32101393, 32271687), The Key Research and Development Program of Guangdong Province (2022B1111230001), Joint Funds of the National Natural Science Foundation of China (U21A20187), and China Postdoctoral Science Foundation (2021M703257).
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to thank Guoxiang Niu and Xuebing Zhangfor their assistance in the field work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Luo, Y.H.; Cadotte, M.W.; Liu, J.; Burgess, K.S.; Tan, S.L.; Ye, L.J.; Zou, J.Y.; Chen, Z.Z.; Jiang, X.L.; Li, J.; et al. Multitrophic diversity and biotic associations influence subalpine forest ecosystem multifunctionality. Ecology 2022, 103, e3745. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.; Ineson, P.; Heal, O.W.; Dhillion, S. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Lavelle, P. Functional domains in soils. Ecol. Res. 2002, 17, 441–450. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A., Jr. Effects of microfauna on the seasonal dynamics of nutrients in forest litter. Soil Biol. Biochem. 1980, 12, 337–342. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Crossley, D.A., Jr. Nutrients in forest litter treated with naphthalene and simulated throughfall: A field microcosm study. Soil Biol. Biochem. 1983, 15, 159–165. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Chan, K.F. Experimental evidence that soil fauna enhance nutrient mineralization and plant nutrient uptake in montane grassland ecosystems. Soil Biol. Biochem. 1999, 31, 1007–1014. [Google Scholar] [CrossRef]
- Hasegawa, M.; Takeda, H. Carbon and nutrient dynamics in decomposing pine needle litter in relation to fungal and fauna abundances. Pedobiologia 1996, 40, 171–184. [Google Scholar]
- Topp, W.; Kappes, H.; Kulfan, J.; Zach, P. Distribution pattern of woodlice (Isopoda) and millipedes (Diplopoda) in four primeval forests of the Western Carpathians (Central Slovakia). Soil Biol. Biochem. 2006, 38, 43–50. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Partsch, S.; Parkinson, D.; Scheu, S. Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods and vegetation. Soil Biol. Biochem. 2007, 39, 1099–1110. [Google Scholar] [CrossRef]
- Scheu, S.; Wolters, V. Influence of fragmentation and bioturbation on the decomposition of 14C-labelled beech leaf litter. Soil Biol. Biochem. 1991, 23, 1029–1034. [Google Scholar] [CrossRef]
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms, 3rd ed.; Chapman and Hall: London, UK, 1996. [Google Scholar] [CrossRef]
- Edwards, C.A.; Fletcher, K.E. Interactions between earthworms and micro-organisms in organic matter breakdown. Agric. Ecosyst. Environ. 1988, 24, 235–247. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Liu, T.; Mao, P.; Shi, L.; Eisenhauer, N.; Liu, S.; Wang, X.; He, X.; Wang, Z.; Zhang, W.; Liu, Z.; et al. Forest canopy maintains the soil community composition under elevated nitrogen deposition. Soil Biol. Biochem. 2020, 143, 107733. [Google Scholar] [CrossRef]
- Kudrin, A.A.; Zuev, A.G.; Taskaeva, A.A.; Konakova, T.N.; Kolesnikova, A.A.; Gruzdev, I.V.; Gabov, D.N.; Yakovleva, E.V.; Tiunov, A.V. Spruce girdling decreases abundance of fungivorous soil nematodes in a boreal forest. Soil Biol. Biochem. 2021, 155, 108184. [Google Scholar] [CrossRef]
- Liu, T.; Mao, P.; Shi, L.; Wang, Z.; Wang, X.; He, X.; Tao, L.; Liu, Z.; Zhou, L.; Shao, Y.; et al. Contrasting effects of nitrogen deposition and increased precipitation on soil nematode communities in a temperate forest. Soil Biol. Biochem. 2020, 148, 107869. [Google Scholar] [CrossRef]
- Liu, T.; Wang, X.; Zhang, W.; Eisenhauer, N.; Shao, Y.; Zhao, J.; Li, J.; He, X.; Tao, L.; Fu, S.; et al. Earthworms regulate the nematode community by directly enhancing the bacterial-based energy channel rather than through the effect of casts. Pedobiologia 2022, 95, 150843. [Google Scholar] [CrossRef]
- Yeates, G.W. Soil nematode populations depressed in the presence of earthworms. Pedobiologia 1981, 22, 191–195. [Google Scholar]
- Hyvönen, R.; Andersson, S.; Clarholm, M.; Persson, T. Effects of lumbricids and enchytraeids on nematodes in limed and unlimed coniferous mor humus. Biol. Fertil. Soils 1994, 17, 201–205. [Google Scholar] [CrossRef]
- Domínguez, J.; Parmelee, R.W.; Edwards, C.A. Interaction between Eisenia andrei (Oligochaeta) and nematode populations during vermicomposting. Pedobiologia 2003, 47, 53–60. [Google Scholar] [CrossRef]
- Yu, F.; Li, C.K.; Liu, T.; Li, T.; Hu, F.; Li, H.X.; Jiao, J.G. Earthworm mucus interfere in the behavior and physiology of bacterial-feeding nematodes. Appl. Soil Ecol. 2019, 143, 107–115. [Google Scholar] [CrossRef]
- Senapati, B.K. Biotic interactions between soil nematodes and earthworms. Soil Biol. Biochem. 1992, 24, 1441–1444. [Google Scholar] [CrossRef]
- Boyer, J.; Reversat, G.; Lavelle, P.; Chabanne, A. Interactions between earthworms and plant-parasitic nematodes. Eur. J. Soil Biol. 2013, 59, 43–47. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Berry, E.C.; Lewis, L.C. Interactions between nematodes and earthworms: Enhanced dispersal of Steinernema carpocapsae. J. Nematol. 1993, 25, 189–192. [Google Scholar] [PubMed]
- Villenave, C.; Rabary, B.; Kichenin, E.; Djigal, D.; Blanchart, E. Earthworms and plant residues modify nematodes in tropical cropping soils (Madagascar): A mesocosm experiment. Appl. Environ. Soil Sci. 2010, 323640. [Google Scholar] [CrossRef]
- Phillips, G.; Yates, D.I.; Shelley, R.M.; Ortstadt, P.R.; Bernard, E.C. Investigating commensal relationships of nematodes in millipedes: Life in unexpected Places. Am. Biol. Teach. 2019, 81, 278–283. [Google Scholar] [CrossRef]
- Phillips, D.; Bernard, E.C.; Shelley, R.M.; Sun, X. Diversity, abundance and prevalence of nematodes inside the intestine of North American millipedes. J. Nematol. 2016, 48, 362. [Google Scholar]
- Senapati, B.K.; Dash, M.C. Effect of grazing on the elements of production in vegetation and Oligochaete components of a tropical pasture. Rev. Decol. Biol. Sol. 1981, 18, 487–505. [Google Scholar]
- Aira, M.; Sampedro, L.; Monroy, F.; Domínguez, J. Detritivorous earthworms directly modify the structure, thus altering the functioning of a microdecomposer food web. Soil Biol. Biochem. 2008, 40, 2511–2516. [Google Scholar] [CrossRef]
- Barker, K.R. Nematode extraction and bioassays. In An Advanced Treatise on Meloidogyne; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; Methodology; North Carolina State University: Raleigh, NC, USA, 1985; Volume 2, pp. 19–35. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.L.C.; Guan, P.; Cui, S.; de Tomasel, C.M.; Gherardi, L.A.; Sala, O.E.; Wall, D.H. Precipitation effects on nematode diversity and carbon footprint across grasslands. Glob. Chang. Biol. 2022, 28, 2124–2132. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Freckleton, R.P.; Godfray, H.; Beissinger, S.R.; Benton, T.; Cameron, D.D.; Carmel, Y.; Coomes, D.A.; Coulson, T.; Emmerson, M.C.; et al. Identification of 100 fundamental ecological questions. J. Ecol. 2013, 101, 58–67. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, X.; Zhang, W.; Lu, X.; Liu, T. Effects of exotic and native earthworms on soil micro-decomposers in a subtropical forest: A field mesocosm experiment. Forests 2022, 13, 1924. [Google Scholar] [CrossRef]
- Niu, X.; Zhai, P.; Zhang, W.; Gu, Y. Effects of Earthworms and Agricultural Plant Species on the Soil Nematode Community in a Microcosm Experiments. Sci. Rep. 2019, 9, 11660. [Google Scholar] [CrossRef]
- Anderson, J.M.; Bignell, D.E. Bacteria in the food, gut contents and faeces of the litter-feeding millipede Glomeris marginata (Villers). Soil Biol. Biochem. 1980, 12, 251–254. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).