Abstract
Western pine beetle (Dendroctonus brevicomis LeConte) is a major cause of ponderosa pine (Pinus ponderosa Dougl. ex. Laws.) mortality in western North America. Twenty-first century epidemics are among the largest in history and have affected hundreds of thousands of hectares. We synthesize literature on the chemical ecology of western pine beetle and on efforts to exploit our understanding of the western pine beetle-ponderosa pine system to reduce host tree losses. This literature dates back to the early 20th century and focuses on populations in California and Oregon, U.S., where western pine beetle exerts its largest impacts. Research in the 1960s–1970s yielded an effective semiochemical attractant (exo-brevicomin, frontalin, and myrcene) that helped inform understanding of the biology, ecology, and management of this species. Later, research focused on isolation and identification of semiochemical repellents. To date, Verbenone Plus (acetophenone, (E)-2-hexen-1-ol + (Z)-2-hexen-1-ol, and verbenone) is the only semiochemical repellent demonstrated effective for protecting ponderosa pines from mortality attributed to western pine beetle in multiple studies in Canada and the U.S.
1. Introduction
Bark beetles (Coleoptera: Curculionidae, Scolytinae) represent a large and diverse group of insects consisting of ~550 species in North America [1]. However, only ~20 species are important disturbance agents in conifer forests [2]. Trees of all species, ages, and sizes may be colonized and killed by bark beetles, but each species exhibits unique host preferences, life history traits, and impacts. Some species, such as California fivespined ips (Ips paraconfusus Lanier), primarily colonize stressed, dead, or dying host trees at small spatial scales (<100 ha). Other species, such as western pine beetle (Dendroctonus brevicomis LeConte), can kill large numbers of host trees over extensive areas (>100,000 ha) [2]. At endemic population levels, bark beetles create small gaps in the forest canopy by killing trees stressed by age, drought, defoliation, or other factors. This differs from epidemic population levels wherein timber and fiber production, water quality and quantity, fish and wildlife populations, recreation, grazing capacity, real estate values, biodiversity, carbon storage, cultural resources, and endangered species, among other resources, may be affected [3].
Bark beetle epidemics develop when weather conducive to beetle survival and population growth (i.e., typically warm and dry weather) co-occurs with an abundance of susceptible host trees [4]. In recent years, this was demonstrated for several species, including western pine beetle, when large epidemics occurred in the central and southern Sierra Nevada, California, U.S. In the most heavily affected areas, ~49% of trees died during 2014–2017 [5] (Figure 1). Ponderosa pine (Pinus ponderosa Dougl. ex Laws.) (Pinales: Pinaceae) had the highest levels of mortality (~90%), which was concentrated in larger-diameter trees (>32 cm dbh, and a diameter of 1.37 m) and attributed primarily to western pine beetle [5]. Levels of tree mortality observed are considered by some experts to be unprecedented [6]. A decade earlier a notable western pine beetle epidemic occurred in the Transverse and Peninsula Ranges in southern California [7] where the majority of ponderosa and Coulter pines (Pinus coulteri D. Don), the two primary hosts, were killed. Tree mortality was >80% in some areas [8]. Western pine beetle occurs from British Columbia, Canada southward through parts of Washington, Oregon, Idaho, Nevada, Montana, and California, U.S. [9]. Dendroctonus barberi Hopkins occurs in eastern Nevada, Utah, Colorado, Arizona, New Mexico, and Texas, U.S., and Mexico. We caution the reader that D. barberi was synonymized with D. brevicomis in 1963 but was reinstated as a separate species in 2019 based on differences in morphological characteristics, chemical ecology, and other factors [10,11,12,13].

Figure 1.
Tree mortality in the Sequoia National Forest, California, 2017. Most of the faded trees were colonized and killed by western pine beetle (Dendroctonus brevicomis) in 2016. Photo credit: C.J. Fettig, USDA Forest Service.
2. Host Finding, Host Selection, and Host Colonization
Bark beetles maintain limited energy reserves and are susceptible to adverse weather, starvation, and predation when searching for host trees. As such, it is important that bark beetles efficiently locate the correct habitat, correct tree species, and the most susceptible trees within these species [14,15]. During host finding and host selection, western pine beetle uses a combination of random landings, visual orientations, and direct assessments of hosts (and nonhosts) via olfactory and gustatory cues [16,17]. If the host is selected, pioneering (female) western pine beetles bore through the outer bark and initiate gallery construction in the phloem. Successful colonization of host trees may require overcoming formidable constitutive and induced physical and chemical defenses [18]. Healthy pines are capable of mobilizing large amounts of resin upon wounding of the tree by beetles. Pioneers are often killed by drowning or immobilization in resin that fills portions of the gallery and collects at the entrance hole (Figure 2). Smith [19] proposed that resistance of ponderosa pine to western pine beetle could be calculated as a function of resin quantity and resin quality/attack density and beetle quality. At the time, he indicated that there were “no known reliable measures of beetle quality” but suggested that the prevalence of nematodes and/or variations in fecundity might serve as useful measures [19]. Resin terpenes are highly toxic to beetles at high vapor concentrations [20,21].

Figure 2.
A western pine beetle (Dendroctonus brevicomis) killed in resin accumulated at the entrance hole, California. Photo credit: C.J. Fettig, USDA Forest Service.
Like most major tree-killing bark beetle species, western pine beetle uses a complex system of semiochemical communication in host location, host selection, host colonization, and mating behaviors (Table 1). Acoustic communication may also play a role [22] but has not been adequately studied. During the early stages of colonization, female western pine beetle release exo-brevicomin synthesized de novo, which in combination with host terpenes is attractive to conspecifics [23]. Frontalin, synthesized de novo by males [24], enhances attraction and facilitates mass attack (i.e., an adequate number of beetles to overcome host tree defenses) [25,26]. Both sexes orient upwind to these stimuli [27]. Verbenone, trans-verbenol, and ipsdienol are also produced by western pine beetle (Table 1) and inhibit the response of western pine beetle to aggregation pheromones thus reducing attraction to the host tree [28,29]. The relative abundance of these and other semiochemicals fluctuates during the colonization sequence (Table 1), with ratios influencing behavioral responses. There are one to four generations per year throughout most of the range. Voltinism may be increasing in some populations of western pine beetle due to warming attributed to climate change [30].

Table 1.
Major semiochemicals associated with western pine beetle (Dendroctonus brevicomis) used in management and/or research.
3. Management Tools and Tactics
Substantial basic and applied research has been devoted to the development of tools and tactics for mitigating undesirable levels of tree mortality attributed to western pine beetle. Most of this work has focused on western pine beetle populations in California and Oregon where the beetle causes substantial host tree losses [9,31]. Much of the early knowledge gained was published in a seminal book, “Biology and Control of the Western Pine Beetle”, by noted USDA Forest Service scientists J.M. Miller and F.P. Keen [31]. Direct control (suppression) involves short-term tactics designed to address current infestations and includes the use of insecticides (bole sprays and systemic injections), semiochemicals (attractants and repellents, as shown below), sanitation harvests, or a combination of these and other treatments. Indirect control (prevention) reduces the probability and severity of future infestations by manipulating stand, forest, and/or landscape conditions by reducing the number of susceptible hosts, primarily through thinning and prescribed burning [32]. It is important to note that killing weakened trees is an important ecosystem function fulfilled by western pine beetle during endemic populations [9].
3.1. Semiochemical Attractants
In 1931, Person [33] suggested that aggregation of western pine beetle resulted from chemically mediated attraction. This was informed by his studies in California and Oregon on the beetle’s preference for colonizing medium-large diameter ponderosa pines (~51–76 cm dbh) with small crowns [34]. He concluded that “…an initial weak attraction is due to the formation of volatile oils, such as aldehydes or esters, which are by-products of a respiratory fermentation or abnormal enzyme activity in subnormal trees. This attracts beetles from the immediate vicinity, these in turn introduce a yeast into the inner bark which produces a fermentation strong enough to attract other beetles from a wider radius.” Substantial advances in understanding of bark beetle attraction have been made since the work of Person [34]. However, it was not until the mid-1960s that the first bark beetle pheromones were isolated and identified [35,36]. Today, commercial lures and baits containing the aggregation pheromones exo-brevicomin and frontalin and the host volatile myrcene are readily available and effective attractants for surveying and detecting western pine beetle populations and for inducing mass attacks [37] (Table 1). Semiochemical attractants are not registered with the U.S. Environmental Protection Agency or the Health Canada Pest Management Regulatory Agency.
3.1.1. Baited Traps for Survey, Detection, and Experimental Purposes
Baited traps are used for surveying and detecting western pine beetle and for monitoring of diurnal and seasonal flight for a variety of purposes [38], including timing of control tactics. Unbaited traps catch very few beetles (e.g., <1/d) even during severe epidemics. Baited traps are also commonly used in assays evaluating the effectiveness of semiochemical repellents for use as tree protectants (e.g., [39,40]). Baited traps should be placed >20 m from susceptible host trees to avoid spillover (i.e., a situation in which beetles are attracted to baited traps (or baited trees) but colonize adjacent host trees) (Figure 3), which may confound trap catches and result in undesirable tree mortality. It is important to note that studies in California have shown that the use of baited multiple-funnel traps is not effective for predicting future levels of tree mortality attributed to western pine beetle [41]. Higher doses of attractants generally result in higher trap catches [42] but increase the risk of spillover.

Figure 3.
A multiple-funnel trap baited with exo-brevicomin, frontalin, and myrcene to collect western pine beetles (Dendroctonus brevicomis) for experimental purposes, California. Photo credit: L.A. Mortenson, USDA Forest Service.
Predatory beetles such as Temnoscheila chlorodia (Mannerheim) (Coleoptera: Trogossitidae) and Enoclerus lecontei (Wolcott) (Coleoptera: Cleridae) are among the first insects to respond to ponderosa pines colonized by western pine beetle [43]. These and other natural enemies are likely important in regulating western pine beetle populations at endemic levels but not at epidemic levels [44]. Some predatory beetles are attracted to western pine beetle pheromones and may be captured and killed in large numbers in baited traps [45]. For example, Fettig and Dabney [46] described the abundance of bark beetle predators collected in 40 multiple-funnel traps baited with exo-brevicomin, frontalin, and myrcene in California over two years. A total of 32,903 T. chlorodia, 79 E. lecontei, and 12 E. sphegeus (F.) were collected. Caution should be taken to limit the bycatch of these and other natural enemies as much as possible when using baited traps [47,48]. For example, screen filters may be used to prevent the passage of predators larger than western pine beetle (e.g., T. chlorodia and Enoclerus spp.) into the collection cup.
3.1.2. Mass Trapping for Population Suppression
Evaluating the effectiveness of mass trapping is difficult and hindered by the inability to accurately estimate bark beetle populations and to determine the effects of mass trapping on local populations. The first extensive study of mass trapping for suppression of western pine beetle was conducted in California in the early 1970s [49,50]. An area of ~65 km2 was divided into two plots treated with baited sticky traps [51] and two untreated plots. Over 400,000 western pine beetles were collected. Ponderosa pine losses were higher in treated plots than in untreated plots the year (1970) treatments were implemented. The following year there was some evidence of population suppression based on declines in levels of ponderosa pine mortality. Despite this, mass trapping is not viewed as an effective control tactic for western pine beetle [52]. However, the use of modern tools (traps, baits, etc.) and technologies for mass trapping of western pine beetle have not been adequately studied. In general, the use of intelligent traps (e.g., those that release attractants at optimal rates and times of day) and/or remotely sensed data have not been adequately explored for mass trapping of bark beetles [53].
3.1.3. Baited Trap Trees for Population Suppression
Limited research has focused on the use of baited trap trees whereby living host trees are treated with an insecticide and then baited to induce the mortality of attacking beetles. In California, Smith [54] demonstrated that treating the boles of ponderosa pines with carbaryl, lindane, permethrin, or deltamethrin and then inducing attack resulted in mortality of large numbers of western pine beetles (i.e., ~1075–5914 beetles/m2 of bark surface). However, spillover effects were observed on adjacent (unbaited) ponderosa pines [54]. In addition, there are concerns regarding the effects of insecticides on nontarget invertebrates, specifically natural enemies that may be attracted to baited trap trees. The use of baited trap trees is not viewed as a viable control tactic [52].
3.1.4. Baited Trees for Snag Creation
Snags are important components of forest ecosystems providing feeding substrates, nesting and roosting sites, and habitats for a variety of vertebrate and invertebrate species. In some situations, forests may be snag deficient. A common technique to create snags is mechanical girdling with a chainsaw which prevents the transport of water and nutrients within the tree resulting in mortality of the tree. However, this technique is costly, laborious, and requires substantial personal protective equipment (PPE). In many conifer forests, snags can be created by placing baits on individual trees to induce colonization by bark beetles (Figure 4). Research suggests that snags created by baiting for western pine beetles are more biologically rich than those created by mechanical girdling with a chainsaw. For example, in California the number of emergence holes from wood-boring insects was ~16–20X higher on baited than girdled snags [55]. Six years after baiting, 44% of baited snags had woodpecker cavities, while no cavities were found in girdled snags [55]. When baiting trees to create snags, caution should be taken to avoid spillover. This can be achieved by regular monitoring of baited and surrounding trees and removal of baits once the baited tree has been adequately colonized (e.g., >75 attacks/m2).

Figure 4.
A ponderosa pine (Pinus ponderosa) snag created by baiting several years earlier, California. The parental “S-shaped” galleries of western pine beetle (Dendroctonus brevicomis) are still partially visible on the bole. Photo credit: C.J. Fettig, USDA Forest Service.
3.1.5. Baited Trees for Experimental Purposes
Tree baits may be used for a variety of experimental purposes. The most common purpose is baiting of host trees to test the efficacy of semiochemical repellents (e.g., [56,57,58,59]) or insecticides (e.g., [60,61]) for tree protection. It is rare for a high proportion (>30%) of unbaited host trees to be attacked by western pine beetles in any given year, and if trees are not baited investigators run the risk of few experimental trees being challenged (attacked) by beetles. Typically, repellents or insecticides (and a control) are randomly assigned to 20–30 trees per treatment and later baited with the goal of inducing attack by western pine beetles. Efficacy is based on comparisons of attack densities (e.g., by counting numbers of successful attacks (i.e., oxidized phloem material present in pitch tubes or points of attack containing boring dust and/or dry frass) and unsuccessful attacks (i.e., pitch tubes without oxidized phloem material)) and/or levels of tree mortality among treatments.
3.2. Semiochemical Repellents
Since the 1980s, the bulk of research on the applied chemical ecology of western pine beetle has focused on development of semiochemical repellents for tree protection. Bedard et al. [62] showed that verbenone reduced the number of western pine beetles collected in baited traps in California. Trap catches were further reduced by higher releases of verbenone [62,63,64,65], and by combining verbenone with ipsdienol [66]. Ipsdienol is produced by the male western pine beetle [67,68] and sympatric Ips and Dendroctonus spp. [69] (Table 1). Shea and Wentz [70] evaluated the effect of chirality on response of western pine beetle to verbenone in California and found that racemic and 97%-(–) verbenone were most effective in reducing attraction. It is assumed that in nature verbenone reduces intraspecific competition, and in some cases interspecific competition, by altering adult beetle behavior to minimize overcrowding of brood within the host tree [28,29]. Lindgren et al. [71] proposed that verbenone is an indicator of host tissue quality and that its quantity is a function of microbial degradation.
Verbenone was the focus of early research evaluating semiochemicals for tree protection. Bertram and Paine [72] reported that verbenone and ipsdienol significantly reduced numbers of western pine beetles landing on ponderosa pines and attack densities on ponderosa pines in California. Unfortunately, levels of tree mortality were not determined [72]. Verbenone applied to the stem of individual ponderosa pines in a flake formulation was ineffective at preventing western pine beetle colonization in California [73]. In Oregon, the effectiveness of 5 g and 7 g verbenone pouches, within an integrated approach that included infested tree removals (sanitation) and mass trapping, for protecting old-growth stands of ponderosa pine were inconclusive (J.L. Hayes, USFS, pers. comm. in [59]). Research in California also showed that 5 g verbenone pouches were ineffective [57]. No significant differences in the levels of western pine beetle-caused tree mortality or in the percentage of unsuccessfully attacked host trees were found between verbenone-treated and untreated plots (Figure 5). As such, verbenone (alone) is not considered effective for protecting individual ponderosa pines or stands of ponderosa pine from western pine beetle [52] (Table 1).
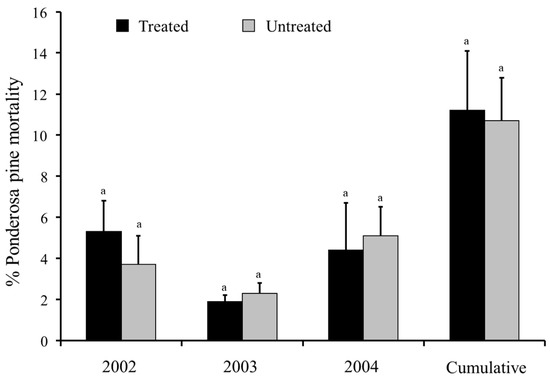
Figure 5.
Mean percentages (+SEM) of ponderosa pines (Pinus ponderosa) killed by western pine beetle (Dendroctonus brevicomis) in verbenone-treated and untreated plots, California. Bars preceded by the same letter within year and cumulatively (2002–2004) are not significantly different (p > 0.05; Tukey’s HSD). Adapted from Fettig et al. [57].
Zhang and Schlyter [74] introduced the semiochemical diversity hypothesis as one factor explaining why homogenous forest stands experience epidemics of insect pests more often than heterogeneous stands do. In the context of pest management, a diverse array of chemical cues and signals may disrupt bark beetles searching more than high doses of a single semiochemical (e.g., verbenone) or even mixtures of semiochemicals mimicking one type of signal (e.g., antiaggregation pheromones) because they represent heterogeneous stands. Poland et al. [39] examined the effects of nonhost angiosperm volatiles on western pine beetle attraction in British Columbia and reported that the green leaf aldehyde (E)-2-hexenal, and two green leaf alcohols (E)-2-hexen-1-ol and (Z)-2-hexen-1-ol significantly reduced numbers of males caught in baited traps (Table 1). (Z)-2-Hexen-1-ol also reduced numbers of females (pioneers) caught in baited traps. Fettig et al. [40] reported that combinations of angiosperm bark volatiles (benzaldehyde, benzyl alcohol, (E)-conophthorin, guaiacol, nonanal, and salicylaldehyde), green leaf volatiles ((E)-2-hexenal, (E)-2-hexen-1-ol, and (Z)-2-hexen-1-ol), or the nine compounds combined did not affect the response of western pine beetle to baited traps in California. However, when bark and green leaf volatiles were combined with verbenone trap catches were reduced to levels significantly below that of verbenone alone. No difference was observed between males and females [40]. A nine-component blend (benzyl alcohol, benzaldehyde, guaiacol, nonanal, salicylaldehyde, (E)-2-hexenal, (E)-2-hexen-1-ol, (Z)-2-hexen-1-ol and verbenone) reduced trap catches by ~87% compared to the baited control (Figure 6). Based on this research, Fettig et al. [56] were first to demonstrate the successful application of semiochemical repellents for protecting ponderosa pine from mortality attributed to western pine beetle (Figure 7). Additional research confirmed the effect [58].
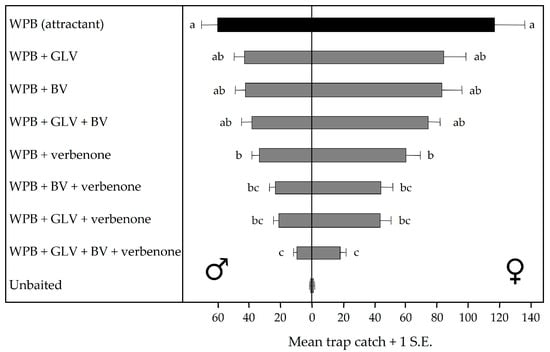
Figure 6.
Daily captures of western pine beetle (Dendroctonus brevicomis) in multiple-funnel traps baited with an attractant (WPB), or with an attractant and green leaf volatile blend (GLV), bark volatile blend (BV), nonhost angiosperm volatile blend (GLV + BV), and verbenone, California. Bars preceded (for males) and followed (for females) by the same letter are not significantly different (n = 30; Tukey’s HSD; p > 0.05). Adapted from Fettig et al. [40].

Figure 7.
Fettig et al. [56] examined nonhost angiosperm volatiles and verbenone for protecting individual ponderosa pines (Pinus ponderosa) from western pine beetle (Dendroctonus brevicomis) in California. A combination of verbenone and nonhost angiosperm volatiles significantly reduced attack densities and mortality of baited trees. Components were formulated in four bubble-capsule bands based on similarity in chemical structure (i.e., alcohols, aldehydes, guaiacol (a phenol), and verbenone (a ketone)). This research informed development of Verbenone Plus in subsequent years [59]. Photo credit: C.J. Fettig, USDA Forest Service.
Fettig et al. [75] further examined the response of western pine beetle to several blends of nonhost angiosperm volatiles and verbenone in baited traps with the goal of improving the efficacy of their blend and reducing the number of components. They documented the inhibitory effect of a five-component blend (nonanal, (E)-2-hexenal, (E)-2-hexen-1-ol, (Z)-2-hexen-1-ol, and verbenone), and later demonstrated that adding acetophenone [76,77] allowed the removal of the aldehydes (nonanal and (E)-2-hexenal) without compromising levels of inhibition. The resulting blend (acetophenone, (E)-2-hexen-1-ol + (Z)-2-hexen-1-ol, and verbenone), “Verbenone Plus”, has been demonstrated to inhibit the response of western pine beetle to baited traps and baited trees in multiple studies in California and British Columbia.
Related efforts to add acetophenone, (E)-2-hexen-1-ol, and (Z)-2-hexen-1-ol to SPLAT® Verb (ISCA Technologies Inc., Riverside, CA, USA) (=“SPLAT® Verbenone Plus”) were ineffective for protecting stands of ponderosa pine from western pine beetle in California and Oregon (C.J.F. and R.A.P., unpubl. data) (Figure 8). SPLAT® Verb is a flowable, controlled-release emulsion [78] containing verbenone for management of mountain pine beetle (Dendroctonus ponderosae Hopkins). It was registered by the U.S. Environmental Protection Agency for use on pines in 2013 [79]. An advantage of SPLAT® Verb is that, unlike other formulations that release semiochemicals through plastic membranes (i.e., requiring retrieval of release devices), SPLAT® Verb biodegrades in ~1–2 years under normal field conditions. A gas chromatograph with a flame ionization detector (GC-FID) was used to quantify the amount of active ingredients (acetophenone, (E)-2-hexen-1-ol, (Z)-2-hexen-1-ol, and verbenone) remaining in samples of SPLAT® Verbenone Plus harvested from experimental plots after 60 d. (E)-2-hexen-1-ol and (Z)-2-hexen-1-ol were nearly undetectable, indicating that sufficient releases of (E)-2-hexen-1-ol and (Z)-2-hexen-1-ol were not maintained throughout the flight activity period of western pine beetle (C.J.F. et al., unpubl. data). Alternatives are being explored in the laboratory to increase the longevity of release of (E)-2-hexen-1-ol and (Z)-2-hexen-1-ol within the SPLAT® matrix, but have not been evaluated in the field. The rheological properties of SPLAT® can be adjusted to create emulsions with a wide range of physical properties [78].

Figure 8.
An early prototype of SPLAT® Verbenone Plus (acetophenone, (E)-2-hexen-1-ol, (Z)-2-hexen-1-ol, and verbenone; ISCA Technologies Inc., Riverside, CA, USA) applied to the bole of a ponderosa pine (Pinus ponderosa), California. Photo credit: C.J. Fettig, USDA Forest Service.
Several new semiochemical repellents were evaluated in trapping assays for western pine beetle in California and Idaho in 2022, but none appeared to be more effective than Verbenone Plus (C.S.H. et al., unpubl. data). Another tactic for managing bark beetles with semiochemicals is “push-pull,” which causes beetles to disperse from a stand by combining semiochemical repellents within the stand (=push) with semiochemical attractants along the periphery of the stand (=pull) [37]. Push–pull has not been adequately studied for western pine beetle.
Verbenone Plus is the only semiochemical repellent that has been demonstrated to be effective for protecting ponderosa pines from mortality attributed to western pine beetle in multiple studies [80], but has yet to be commercialized. This is due, in part, to semiochemical repellents requiring registration with the U.S. Environmental Protection Agency and the Health Canada Pest Management Regulatory Agency. This takes time and is costly relative to the value of the marketplace (i.e., ponderosa pine is a minor-use crop with unpredictable fluctuations in western pine beetle populations). Efforts to develop Verbenone Plus for its use as a general bark beetle repellent, with the goal of increasing the marketplace, have been met with limited success. Verbenone Plus was only as effective as verbenone (alone) for protecting lodgepole pine (Pinus contorta Dougl. ex. Laws.) from mortality attributed to the mountain pine beetle in Idaho [81]. Furthermore, levels of inhibition were similar between Verbenone Plus and verbenone (alone) in trapping assays for pine engraver (Ips pini (Say)) in Arizona [82]. However, the addition of acetophenone, (E)-2-hexen-1-ol, and (Z)-2-hexen-1-ol to MCH (3-methyl-2-cyclohexen-1-one), an antiaggregation pheromone of spruce beetle (Dendroctonus rufipennis Kirby), is promising for tree protection based on research in Wyoming, U.S. [83], with additional spruce beetle studies underway in Alaska, U.S., Colorado, and Utah (J.P.A. et al., unpubl. data).
4. Conclusions
Much progress has been made concerning the development of semiochemicals for management of bark beetles since discovery of the first bark beetle pheromone more than half a century ago. Some notable successes include effective repellents for Douglas-fir beetle (Dendroctonus pseudotsugae Hopkins; MCH) and mountain pine beetle (verbenone) [37]. Since the early 2000s, MCH and verbenone have been applied to thousands of hectares of forest in western North America. Effective attractants for western pine beetle were identified in the 1960s–1970s, and their use in research has helped inform understanding of the biology, ecology, and management of western pine beetle. Unfortunately, development of an effective repellent for western pine beetle has been more challenging. It is our hope that with continued research, an effective semiochemical repellent will be commercialized as natural resource managers are increasingly challenged to limit host tree losses to western pine beetle and associated effects on a variety of resource objectives and ecosystem goods and services, a trend likely to continue in the future [4,84].
Author Contributions
Conceptualization, C.J.F. and J.P.A., investigation, C.J.F., J.P.A., C.S.H. and R.A.P.; writing—original draft preparation, C.J.F. and J.P.A.; writing—review and editing, C.J.F., J.P.A., C.S.H. and R.A.P.; project administration, C.J.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
We thank numerous colleagues in North America, Europe, and Asia who have shaped our thinking and research on the chemical ecology of western pine beetle and other bark and twig beetles. We thank Shakeeb Hamud for his critiques of earlier drafts of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), A Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982. [Google Scholar]
- Fettig, C.J.; Asaro, C.; Nowak, J.T.; Dodds, K.J.; Gandhi, K.J.K.; Moan, J.E.; Robert, J. Trends in bark beetle impacts in North America during a period (2000–2020) of rapid environmental change. J. For. 2022, 120, 693–713. [Google Scholar] [CrossRef]
- Morris, J.L.; Cottrell, S.; Fettig, C.J.; DeRose, R.J.; Mattor, K.M.; Carter, V.A.; Clear, J.; Clement, J.; Hansen, W.D.; Hicke, J.A.; et al. Bark beetles as agents of change in social-ecological systems. Front. Ecol. Environ. 2018, 16, S34–S43. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Fettig, C.J.; Mortenson, L.A.; Bulaon, B.M.; Foulk, P.B. Tree mortality following drought in the central and southern Sierra Nevada, California, U.S. For. Ecol. Manag. 2019, 432, 164–178. [Google Scholar] [CrossRef]
- Stephens, S.L.; Collins, B.M.; Fettig, C.J.; Finney, M.A.; Hoffman, C.E.; Knapp, E.E.; North, M.E.; Safford, H. Drought, tree mortality, and wildfire in forests adapted to frequent fire. Bioscience 2018, 68, 77–88. [Google Scholar] [CrossRef]
- Fettig, C.J. Socioecological impacts of the western pine beetle outbreak in southern California: Lessons for the future. J. For. 2019, 117, 138–143. [Google Scholar] [CrossRef]
- Walker, R.; Rosenberg, M.; Warbington, R.; Schwind, B.; Beardsley, D.; Ramirez, C.; Fischer, L.; Frerich, B. Inventory of Tree Mortality in Southern California Mountains (2001–2004) Due to Bark Beetle Impacts. 2006. Available online: https://qcpages.qc.cuny.edu/~cyi/Michelle-Park-papers/Walker%20et%20al.,%202006.pdf (accessed on 9 March 2023).
- Homicz, C.S.; Fettig, C.J.; Munson, A.S.; Cluck, D.R. Western Pine Beetle; FS-1192; U.S. Department of Agriculture, Forest Service: Portland, OR, USA, 2022.
- Valerio-Mendoza, O.; Armendáriz-Toledano, F.; Cuéllar-Rodríguez, G.; Negrón, J.F.; Zúñiga, G. The current status of the distribution range of the western pine beetle, Dendroctonus brevicomis (Curculionidae: Scolytinae) in Northern Mexico. J. Insect Sci. 2017, 17, 92. [Google Scholar] [CrossRef][Green Version]
- Valerio-Mendoza, O.; García-Román, J.; Becerril, M.; Armendáriz-Toledano, F.; Cuéllar-Rodríguez, G.; Negrón, J.F.; Sullivan, B.T.; Zúñiga, G. Cryptic species discrimination in western pine beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), based on morphological characters and geometric morphometrics. Insects 2019, 10, 377. [Google Scholar] [CrossRef]
- Sullivan, B.T.; Grady, A.M.; Hofstetter, R.W.; Pureswaran, D.S.; Brownie, C.; Cluck, D.R.; Coleman, T.W.; Graves, A.; Willhite, E.; Spiegel, L.; et al. Evidence for semiochemical divergence between sibling bark beetle species: Dendroctonus brevicomis and Dendroctonus barberi. J. Chem. Ecol. 2021, 47, 10–27. [Google Scholar] [CrossRef]
- Armendáriz-Toledano, F.; Zúñiga, G. Illustrated key to species of genus Dendroctonus (Coleoptera: Curculionidae) occurring in Mexico and Central America. J. Insect Sci. 2017, 17, 34. [Google Scholar] [CrossRef]
- Byers, J.A. Host-tree chemistry affecting colonization in bark beetles. In Chemical Ecology of Insects 2; Carde, R.T., Bell, W.J., Eds.; Springer: Boston, MA, USA, 1995; pp. 154–213. [Google Scholar]
- Borden, J.H. Disruption of semiochemical-mediated aggregation in bark beetles. In Insect Pheromone Research; Carde, R.T., Minks, A.K., Eds.; Springer: Boston, MA, USA, 1997; pp. 421–438. [Google Scholar]
- Strom, B.L.; Goyer, R.A.; Shea, P.J. Visual and olfactory disruption of orientation by the western pine beetle to attractant-baited traps. Entomol. Exp. Appl. 2001, 100, 63–67. [Google Scholar] [CrossRef]
- Huber, D.P.W.; Fettig, C.J.; Borden, J.H. Disruption of coniferophagous bark beetle (Coleoptera: Curculionidae: Scolytinae) mass attack using angiosperm nonhost volatiles: From concept to operational use. Can. Entomol. 2021, 153, 19–35. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.H. Formula for describing effect of insect and host tree factors on resistance to western pine beetle attack. J. Econ. Entomol. 1975, 68, 841–844. [Google Scholar] [CrossRef]
- Smith, R.H. Effect of monoterpene vapors on the western pine beetle. J. Econ. Entomol. 1965, 58, 509–510. [Google Scholar] [CrossRef]
- Seybold, S.J.; Huber, D.P.W.; Lee, J.C.; Graves, A.D.; Bohlmann, J. Pine monoterpenes and pine bark beetles: A marriage of convenience for defense and chemical communication. Phytochem. Rev. 2006, 5, 143–178. [Google Scholar] [CrossRef]
- Bedoya, C.L.; Hofstetter, R.W.; Nelson, X.J.; Hayes, M.; Miller, D.R.; Brockerhoff, E.G. Sound production in bark and ambrosia beetles. Bioacoustics 2021, 30, 58–73. [Google Scholar] [CrossRef]
- Bedard, W.D.; Tilden, P.E.; Wood, D.L.; Silverstein, R.M.; Brownlee, R.G.; Rodin, J.O. Western pine beetle: Field response to its sex pheromone and a synergistic host terpene, myrcene. Science 1969, 164, 1284–1285. [Google Scholar] [CrossRef]
- Kinzer, G.W.; Fentiman, A.F.; Page, T.F.; Foltz, R.L.; Vité, J.P.; Pitman, G.B. Bark beetle attractants: Identification, synthesis and field bioassay of a new compound isolated from Dendroctonus. Nature 1969, 221, 477–478. [Google Scholar] [CrossRef]
- Wood, D.L. Selection and colonization of ponderosa pine by bark beetles. In Insect/Plant Relationships, Symposia of the Royal Entomological Society of London, Vol. 6; Emden van, F., Ed.; Blackwell Scientific: Oxford, UK, 1972; pp. 101–117. [Google Scholar]
- Bedard, W.D.; Lindahl, K.Q., Jr.; Tilden, P.E.; Wood, D.L. Behavior of the western pine beetle during host colonization. J. Chem. Ecol. 1985, 11, 1249–1261. [Google Scholar] [CrossRef]
- Byers, J.A. Upwind flight orientation to pheromone in western pine beetle tested with rotating wind vane traps. J. Chem. Ecol. 1988, 14, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.A.; Wood, D.L. Interspecific inhibition of the response of the bark beetles Dendroctonus brevicomis and Ips paraconfusus to their pheromones in the field. J. Chem. Ecol. 1980, 6, 149–164. [Google Scholar] [CrossRef]
- Byers, J.A.; Wood, D.L.; Craig, J.; Hendry, L.B. Attractive and inhibitory pheromones produced in the bark beetle, Dendroctonus brevicomis, during host colonization: Regulation of inter- and intraspecific competition. J. Chem. Ecol. 1984, 10, 861–877. [Google Scholar] [CrossRef]
- Robbins, Z.J.; Xu, C.; Aukema, B.H.; Buotte, P.C.; Chitra-Tarak, R.; Fettig, C.J.; Goulden, M.L.; Goodsman, D.W.; Hall, A.D.; Koven, C.D.; et al. Warming increased bark beetle-induced tree mortality by 30% during an extreme drought in California. Glob. Change Biol. 2022, 28, 509–523. [Google Scholar] [CrossRef]
- Miller, J.M.; Keen, P.P. Biology and Control of the Western Pine Beetle; Misc. Publ. 800; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1960.
- Fettig, C.J.; Hilszczański, J. Management strategies for bark beetles in conifer forests. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 555–584. [Google Scholar] [CrossRef]
- Person, H.L. Theory in explanation of the selection of certain trees by the western pine beetle. J. For. 1931, 29, 696–699. [Google Scholar] [CrossRef]
- Person, H.L. Tree selection by the western pine beetle. J. For. 1928, 26, 564–578. [Google Scholar] [CrossRef]
- Silverstein, R.M.; Rodin, J.O.; Wood, D.L. Sex attractants in frass produced by male Ips confusus in ponderosa pine. Science 1966, 154, 509–510. [Google Scholar] [CrossRef]
- Renwick, J.A. Identification of two oxygenated terpenes from the bark beetles, Dendroctonus frontalis and Dendroctonus brevicomis. Contrib. Boyce Thompson Inst. 1967, 23, 355–360. [Google Scholar]
- Seybold, S.J.; Bentz, B.J.; Fettig, C.J.; Lundquist, J.E.; Progar, R.A.; Gillette, N.E. Management of western North American bark beetles with semiochemicals. Annu. Rev. Entomol. 2018, 63, 407–432. [Google Scholar] [CrossRef]
- Fettig, C.J.; Shea, P.J.; Borys, R.R. Spatial and temporal distributions of four bark beetle species (Coleoptera: Scolytidae) along two elevational transects in the Sierra Nevada. Pan-Pac. Entomol. 2005, 81, 6–19. [Google Scholar]
- Poland, T.M.; Borden, J.H.; Stock, A.J.; Chong, L.J. Green leaf volatiles disrupt responses by the spruce beetle, Dendroctonus rufipennis, and the western pine beetle, Dendroctonus brevicomis (Coleoptera: Scolytidae) to attractant-baited traps. J. Entomol. Soc. Brit. Columbia 1998, 95, 17–24. [Google Scholar]
- Fettig, C.J.; McKelvey, S.R.; Huber, D.P.W. Nonhost angiosperm volatiles and verbenone disrupt response of western pine beetle, Dendroctonus brevicomis (Coleoptera: Scolytidae), to attractant-baited traps. J. Econ. Entomol. 2005, 98, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.J.; Fettig, C.J.; Merrill, L.D. Evaluation of multiple funnel traps and stand characteristics for estimating western pine beetle-caused tree mortality. J. Econ. Entomol. 2009, 102, 2170–2182. [Google Scholar] [CrossRef] [PubMed]
- Tilden, P.E.; Bedard, W.D.; Lindahl, K.Q.; Wood, D.L. Trapping Dendroctonus brevicomis: Changes in attractant release rate, dispersion of attractant, and silhouette. J. Chem. Ecol. 1983, 9, 311–321. [Google Scholar] [CrossRef]
- Stephen, F.M.; Dahlsten, D.L. The arrival sequence of the arthropod complex following attack by Dendroctonus brevicomis (Coleoptera: Scolytidae) in ponderosa pine. Can. Entomol. 1976, 108, 283–304. [Google Scholar] [CrossRef]
- Wegensteiner, R.; Wermelinger, B.; Herrmann, M. Natural enemies of bark beetles: Predators, parasitoids, pathogens, and nematodes. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 247–304. [Google Scholar] [CrossRef]
- Pitman, G.B.; Vité, J.P. Predator-prey response to western pine beetle attractants. J. Econ. Entomol. 1971, 64, 402–404. [Google Scholar] [CrossRef]
- Fettig, C.J.; Dabney, C.P. Seasonal abundance of Temnochila chlorodia (Mannerheim) (Coleoptera: Trogossitidae) collected in western pine beetle pheromone-baited traps in northern California. J. Entomol. Sci. 2006, 41, 75–83. [Google Scholar] [CrossRef]
- Aukema, B.H.; Dahlsten, D.L.; Raffa, K.F. Improved population monitoring of bark beetles and predators by incorporating disparate behavioral responses to semiochemicals. Environ. Entomol. 2000, 29, 618–629. [Google Scholar] [CrossRef]
- Ross, D.W.; Daterman, G.E. Pheromone-baited traps for Dendroctonus pseudotsugae (Coleoptera: Scolytidae): Influence of selected release rates and trap designs. J. Econ. Entomol. 1998, 91, 500–506. [Google Scholar] [CrossRef]
- DeMars, C.J.; Slaughter, G.W.; Bedard, W.D.; Norick, N.X.; Roettgering, B. Estimating western pine beetle-caused tree mortality for evaluating an attractive pheromone treatment. J. Chem. Ecol. 1980, 6, 853–860. [Google Scholar] [CrossRef]
- Smith, R.H. Direct Control of Western Pine Beetle (Dendroctonus brevicomis LeConte): Review and Assessment; PSW-GTR-121; U.S. Department of Agriculture, Forest Service: Berkeley, CA, USA, 1990. [CrossRef]
- Browne, L.W. A trapping system for the western pine beetle using attractive pheromones. J. Chem. Ecol. 1978, 4, 261–275. [Google Scholar] [CrossRef]
- Fettig, C.J. Native bark beetles and wood borers in Mediterranean forests of California. In Insects and Diseases of Mediterranean Forest Systems; Lieutier, F., Paine, T.D., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 499–528. [Google Scholar]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.J.; Svoboda, M.; et al. Bark beetle outbreaks in Europe: State of knowledge and ways forward for management. Curr. Forestry Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Smith, R.H. Trapping Western Pine Beetles With Baited Toxic Trees; PSW-RN-382; U.S. Department of Agriculture, Forest Service: Berkeley, CA, USA, 1986. [CrossRef]
- Shea, P.J.; Laudenslayer, W.F., Jr.; Ferrell, G.; Borys, R. Girdled versus bark beetle-created ponderosa pine snags: Utilization by cavity-dependent species and differences in decay rate and insect diversity. In Proceedings of the Symposium Ecology and Management of Dead Wood in Western Forests; PSW-GTR-181; U.S. Department of Agriculture, Forest Service: Berkeley, CA, USA, 2002; pp. 145–153. [Google Scholar]
- Fettig, C.J.; Dabney, C.P.; McKelvey, S.R.; Huber, D.P.W. Nonhost angiosperm volatiles and verbenone protect individual ponderosa pines from attack by western pine beetle and red turpentine beetle (Coleoptera: Curculionidae, Scolytinae). W. J. Appl. For. 2008, 23, 40–45. [Google Scholar] [CrossRef]
- Fettig, C.J.; McKelvey, S.R.; Borys, R.R.; Dabney, C.P.; Hamud, S.M.; Nelson, L.J.; Seybold, S.J. Efficacy of verbenone for protecting ponderosa pine stands from western pine beetle (Coleoptera: Curculionidae: Scolytinae) attack in California. J. Econ. Entomol. 2009, 102, 1846–1858. [Google Scholar] [CrossRef]
- Fettig, C.J.; McKelvey, S.R.; Dabney, C.P.; Borys, R.R.; Huber, D.P.W. Response of Dendroctonus brevicomis to different release rates of nonhost angiosperm volatiles and verbenone in trapping and tree protection studies. J. Appl. Entomol. 2009, 133, 143–154. [Google Scholar] [CrossRef]
- Fettig, C.J.; McKelvey, S.R.; Dabney, C.P.; Huber, D.P.W.; Lait, C.G.; Fowler, D.L.; Borden, J.H. Efficacy of “Verbenone Plus” for protecting ponderosa pine trees and stands from Dendroctonus brevicomis (Coleoptera: Curculionidae) attack in British Columbia and California. J. Econ. Entomol. 2012, 105, 668–1680. [Google Scholar] [CrossRef] [PubMed]
- Fettig, C.J.; Allen, K.K.; Borys, R.R.; Christopherson, J.; Dabney, C.P.; Eager, T.J.; Gibson, K.E.; Hebertson, E.G.; Long, D.F.; Munson, A.S.; et al. Effectiveness of bifenthrin (Onyx™) and carbaryl (Sevin® SL) for protecting individual, high-value trees from bark beetle attack (Coleoptera: Curculionidae: Scolytinae) in the western United States. J. Econ. Entomol. 2006, 99, 1691–1698. [Google Scholar] [CrossRef]
- Grosman, D.M.; Fettig, C.J.; Jorgensen, C.L.; Munson, A.S. Effectiveness of two systemic insecticides for protecting western conifers from mortality due to bark beetle attack. W. J. Appl. For. 2010, 25, 181–185. [Google Scholar] [CrossRef]
- Bedard, W.D.; Tilden, P.E.; Lindahl, K.Q., Jr.; Wood, D.L.; Rauch, P.A. Effects of verbenone and trans-verbenol on the response of Dendroctonus brevicomis to natural and synthetic attractant in the field. J. Chem. Ecol. 1980, 6, 997–1013. [Google Scholar] [CrossRef]
- Bedard, W.D.; Wood, D.L.; Tilden, P.E.; Lindahl, K.Q., Jr.; Silverstein, R.M.; Rodin, J.O. Field response of the western pine beetle and one of its predators to host- and beetle-produced compounds. J. Chem. Ecol. 1980, 6, 625–641. [Google Scholar] [CrossRef]
- Tilden, P.E.; Bedard, W.D. Effect of verbenone on response of Dendroctonus brevicomis to exo-brevicomin, frontalin, and myrcene. J. Chem. Ecol. 1988, 14, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.L.; Paine, T.D. Response of Dendroctonus brevicomis LeConte (Coleoptera: Scolytidae) to different release rates and ratios of aggregation semiochemicals and the inhibitors verbenone and ipsdienol. J. Chem. Ecol. 1994, 20, 2931–2941. [Google Scholar] [CrossRef] [PubMed]
- Paine, T.D.; Hanlon, C.C. Response of Dendroctonus brevicomis and Ips paraconfusus (Coleoptera: Scolytidae) to combinations of synthetic pheromone attractants and inhibitors verbenone and ipsdienol. J. Chem. Ecol. 1991, 17, 2163–2176. [Google Scholar] [CrossRef]
- Byers, J.A. Male-specific conversion of the host plant compound, myrcene, to the pheromone, (+)-ipsdienol, in the bark beetle, Dendroctonus brevicomis. J. Chem. Ecol. 1982, 8, 363–371. [Google Scholar] [CrossRef]
- Seybold, S.J.; Teale, S.A.; Wood, D.L.; Zhang, A.; Webster, F.X.; Lindahl, K.Q.; Kubo, I. The role of lanierone in the chemical ecology of Ips pini (Coleoptera: Scolytidae) in California. J. Chem. Ecol. 1992, 18, 2305–2309. [Google Scholar] [CrossRef]
- Borden, J.H. Aggregation pheromones. In Comprehensive Insect Physiology, Biochemistry & Pharmacology, Vol. 9; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 257–285. [Google Scholar]
- Shea, P.J.; Wenz, J.M. Bark beetle research in California. In Proceedings of the Symposium on Management of Western Bark Beetles with Pheromones: Research and Development; GTR-PSW-150; Shea, P.J., Ed.; U.S. Department of Agriculture, Forest Service: Berkeley, CA, USA, 2004; pp. 46–52. [Google Scholar]
- Lindgren, B.S.; Nordlander, G.; Birgersson, G. Feeding deterrence of verbenone to the pine weevil, Hylobius abietis (L.) (Col., Curculionidae). J. Appl. Entomol. 1996, 120, 397–403. [CrossRef]
- Bertram, S.L.; Paine, T.D. Influence of aggregation inhibitors (verbenone and ipsdienol) on landing and attack behavior of Dendroctonus brevicomis (Coleoptera: Scolytidae). J. Chem. Ecol. 1994, 20, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Gillette, N.E.; Stein, J.D.; Owen, D.R.; Webster, J.N.; Fiddler, G.O.; Mori, S.R.; Wood, D.L. Verbenone-releasing flakes protect individual Pinus contorta trees from attack by Dendroctonus ponderosae and Dendroctonus valens (Coleoptera: Curculionidae, Scolytinae). Agric. For. Entomol. 2006, 8, 243–251. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Schlyter, F. Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 2004, 6, 1–20. [Google Scholar] [CrossRef]
- Fettig, C.J.; McKelvey, S.R.; Dabney, C.P.; Huber, D.P.W. Responses of Dendroctonus brevicomis (Coleoptera: Curculionidae) in behavioral assays: Implications to development of a semiochemical-based tool for tree protection. J. Econ. Entomol. 2012, 105, 149–160. [Google Scholar] [CrossRef]
- Erbilgin, N.; Gillette, N.E.; Mori, S.R.; Stein, J.D.; Owen, D.R.; Wood, D.L. Acetophenone as an anti-attractant for the western pine beetle, Dendroctonus brevicomis LeConte (Coleoptera: Scolytidae). J. Chem. Ecol. 2007, 33, 817–823. [Google Scholar] [CrossRef]
- Erbilgin, N.; Gillette, N.E.; Owen, D.R.; Mori, S.R.; Nelson, A.S.; Uzoh, F.; Wood, D.L. Acetophenone superior to verbenone for reducing attraction of western pine beetle Dendroctonus brevicomis to its aggregation pheromone. Agric. For. Entomol. 2008, 10, 433–441. [Google Scholar] [CrossRef]
- Mafra-Neto, A.; de Lame, F.M.; Fettig, C.J.; Munson, A.S.; Perring, T.M.; Stelinski, L.L.; Stoltman, L.; Mafra, L.E.J.; Borges, R.; Vargas, R.I. Manipulation of insect behavior with Specialized Pheromone & Lure Application Technology (SPLAT®). In Pest Management with Natural Products; Beck, J., Coats, J., Duke, S., Koivunen, M., Eds.; American Chemical Society: Washington, DC, USA, 2013; pp. 31–58. [Google Scholar]
- Fettig, C.J.; Munson, A.S.; Reinke, M.; Mafra-Neto, A. A novel semiochemical tool for protecting lodgepole pine from mortality attributed to mountain pine beetle (Coleoptera: Curculionidae). J. Econ. Entomol. 2015, 108, 173–182. [Google Scholar] [CrossRef]
- Borden, J.H. Management of bark and ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) with semiochemicals: Letter to a prospective graduate student. Can. Entomol. 2021, 153, 13–18. [Google Scholar] [CrossRef]
- Fettig, C.J.; Munson, A.S. Efficacy of verbenone and a blend of verbenone and nonhost volatiles for protecting lodgepole pine from mountain pine beetle (Coleoptera: Curculionidae). Agric. For. Entomol. 2020, 22, 373–378. [Google Scholar] [CrossRef]
- Gaylord, M.A.; Audley, J.P.; McMillin, J.D.; Fettig, C.J. Acetophenone and green leaf volatiles do not enhance the efficacy of verbenone for inhibiting attraction of Ips pini (Coleoptera: Curculionidae) to pheromone-baited traps in northern Arizona. J. Econ. Entomol. 2023, 6, toad016. [Google Scholar] [CrossRef]
- Audley, J.P.; Fettig, C.J.; Munson, A.S.; Blackford, D.C.; Mortenson, L.A.; Mafra-Neto, A. MCH-based semiochemical repellents for protecting Engelmann spruce trees from Dendroctonus rufipennis (Coleoptera: Curculionidae). J. Econ. Entomol. 2022, 115, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Robbins, Z.; Xu, C.; Jonko, A.; Chitra-Tarak, R.; Fettig, C.J.; Costanza, J.; Mortenson, L.A.; Aukema, B.H.; Kueppers, L.M.; Scheller, R.M. Carbon stored in live ponderosa pines in the Sierra Nevada will not return to pre-drought (2012) levels during the 21st century due to bark beetle outbreaks. Front. Environ. Sci. 2023, 11, 1112756. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).