
The Impact of Tree Species on Microbial Community Structure and Soil Function on Forest Plantations in the Central Hardwoods Region (CHR)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Sampling
2.2. Physical and Chemical Analysis
2.3. Enzyme Activity (EA)
2.4. EL-FAME Microbial Structure
2.5. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. Soil Microbial Community Structure Based on FAME Analysis
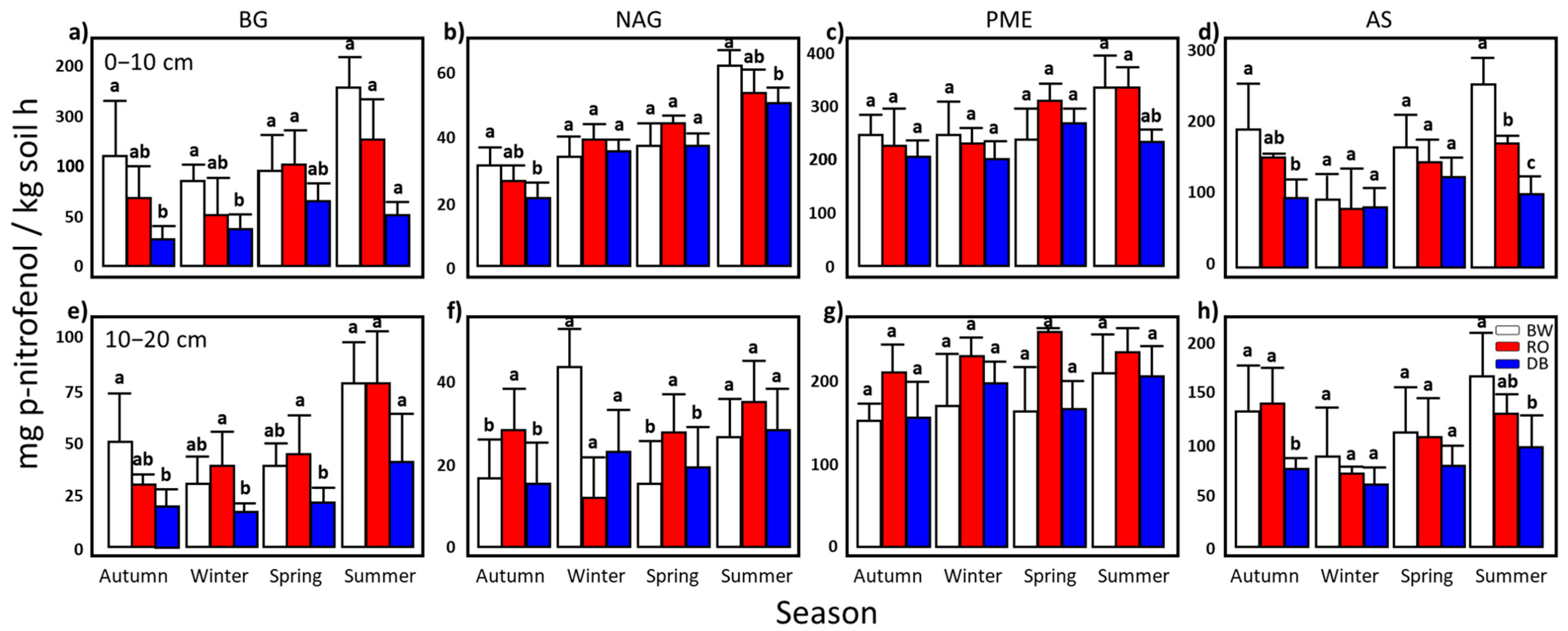
3.3. Soil Biochemical Cycles Measured by Soil Enzyme Activities
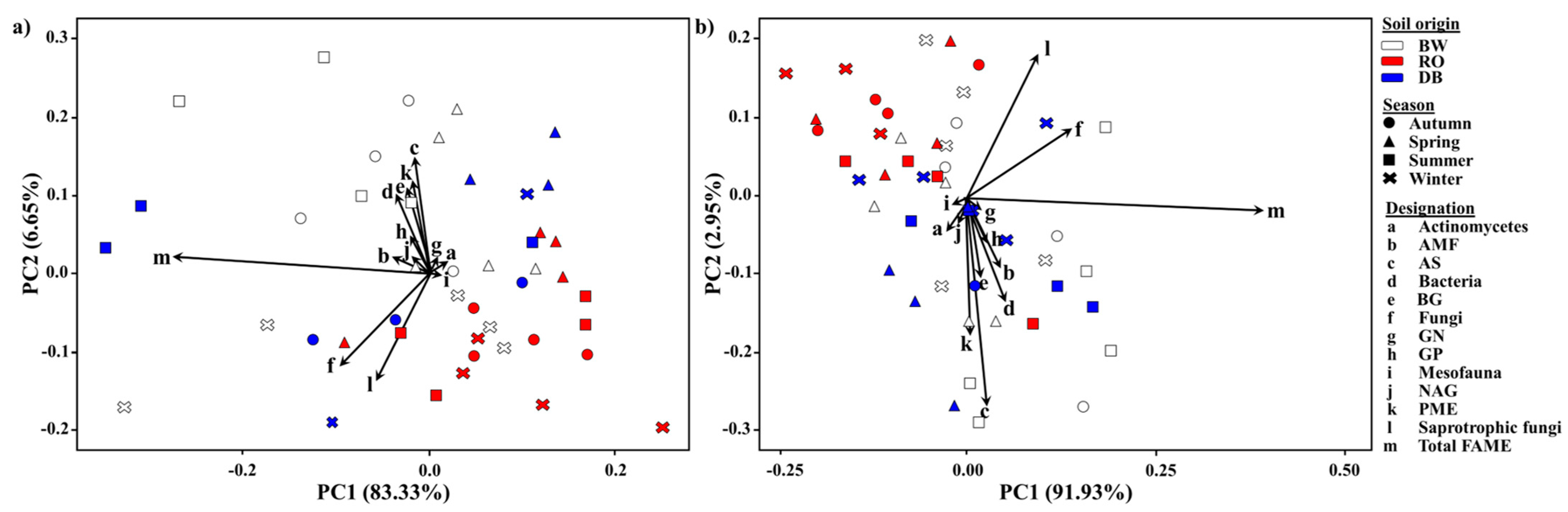
3.4. Principal Component Analysis between Soil Microbial Communities and Soil Enzyme Activities
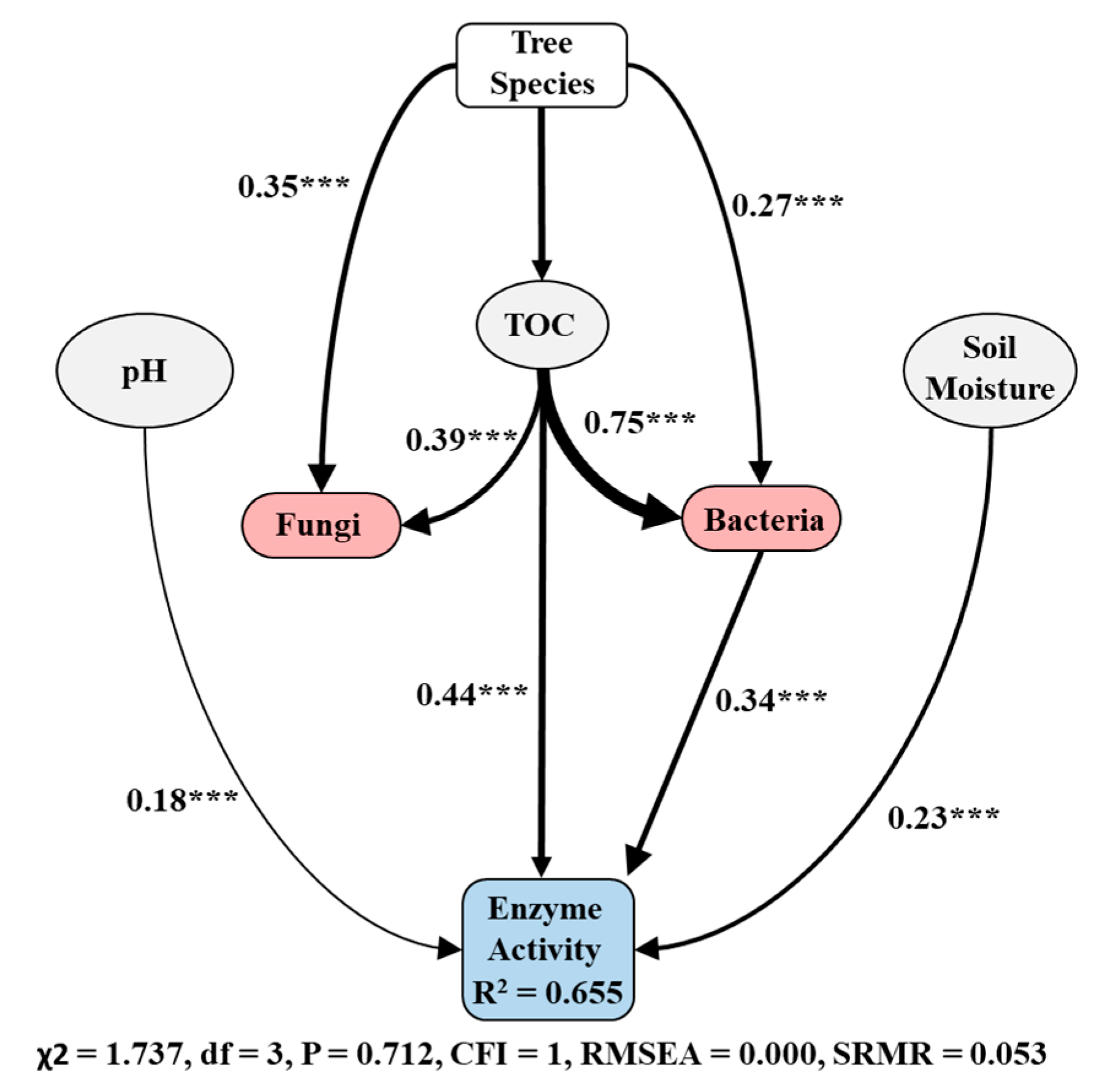
3.5. Association between Soil Microbial Community and Soil Chemical and Ecosystem Functions
4. Discussion
4.1. Soil C and N Responses to Tree Species
4.2. Tree Species Influenced the Soil Microbial Communities and Biogeochemical Cycles
4.3. Heterogeneous Seasonal Variation in Soil Microbial Communities and Biogeochemical Cycles
4.4. Linking among Ecosystem Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ehring, A.; Keller, O. The black walnut tree (Juglans nigra), valuable but with high demands. Wald Holz 2010, 91, 25–28. (In German) [Google Scholar]
- Nicolescu, V.N.; Hernea, C.; Bakti, B.; Keserű, Z.; Antal, B.; Rédei, K. Black locust (Robinia pseudoacacia L.) as a multi-purpose tree species in Hungary and Romania, a review. J. For. Res. 2018, 29, 1449–1463. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Chmura, D.; Dylewski, Ł.; Horodecki, P.; Jagodziński, A.M.; Pietras, M.; Woziwoda, B. Biological flora of the British Isles, Quercus rubra. J. Ecol. 2020, 108, 1199–1225. [Google Scholar] [CrossRef]
- Nicolescu, V.N.; Rédei, K.; Vor, T.; Bastien, J.C.; Brus, R.; Benčať, T.; Štefančik, I. A review of black walnut (Juglans nigra L.) ecology and management in Europe. Trees 2020, 34, 1087–1112. [Google Scholar] [CrossRef]
- Knight, K.S.; Oleksyn, J.; Jagodziński, A.M.; Reich, P.B.; Kasprowicz, M. Overstorey tree species regulate colonization by native and exotic plants, a source of positive relationships between understorey diversity and invisibility. Divers. Distrib. 2008, 14, 666–675. [Google Scholar] [CrossRef]
- Dobrylovská, D. Litter decomposition of red oak, larch and lime tree and its effect on selected soil characteristics. J. For. Sci. 2001, 47, 477–485. [Google Scholar]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties, a common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Davis, A.S. Genetic considerations in the operational production of hardwood nursery stock in the eastern USA. Nativ. Plants J. 2005, 6, 4–13. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—Current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Uroz, S.; Buee, M.; Deveau, A.; Mieszkin, S.; Martin, F. Ecology of the forest microbiome, highlights of temperate and boreal ecosystems. Soil Biol. Biochem. 2016, 103, 471–488. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Shade, A.; Violle, C. When, where and how does microbial community composition matter? Front. Microbiol. 2014, 5, 497. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function, are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef] [PubMed]
- Thoms, C.; Gattinger, A.; Jacob, M.; Thomas, F.M.; Gleixner, G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biol. Biochem. 2010, 42, 1558–1565. [Google Scholar] [CrossRef]
- Smith, J.M.; Paritsis, J.; Veblen, T.T.; Chapman, T.B. Permanent forest plots show accelerating tree mortality in subalpine forests of the Colorado Front Range from 1982 to 2013. For. Ecol. Manag. 2015, 341, 8–17. [Google Scholar] [CrossRef]
- Landesman, W.J.; Freedman, Z.B.; Nelson, D.M. Seasonal, sub-seasonal and diurnal variation of soil bacterial community composition in a temperate deciduous forest. FEMS Microbiol. Ecol. 2019, 95, fiz002. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Voříšková, J.; Větrovský, T.; Baldrian, P. The bacterial community inhabiting temperate deciduous forests is vertically stratified and undergoes seasonal dynamics. Soil Biol. Biochem. 2015, 87, 43–50. [Google Scholar] [CrossRef]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Seasonal variation in functional properties of microbial communities in beech forest soil. Soil Biol. Biochem. 2013, 60, 95–104. [Google Scholar] [CrossRef]
- Ma, W.; Yang, Z.; Liang, L.; Ma, Q.; Wang, G.; Zhao, T. Seasonal Changes in Soil Microbial Community and Co-Occurrence Network of Species of the Genus Corylus. Microorganisms 2021, 9, 2228. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, L.; Wang, Z.; Fan, J. Temporal variations in soil juglone and soil microbial community structure under Manchurian walnut (Juglans mandshurica Maxim.) plantations. Allelopath. J. 2013, 31, 169–180. [Google Scholar]
- Talbot, J.M.; Finzi, A.C. Differential effects of sugar maple, red oak, and hemlock tannins on carbon and nitrogen cycling in temperate forest soils. Oecologia 2008, 155, 583–592. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis, Part 2. Microbiological and Biochemical Properties; SSSA Book Series; Bottomley, P.S., Angle, J.S., Weaver, R.W., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Parham, J.A.; Deng, S.P. Detection, quantification and characterization of β-glucosaminidase activity in soil. Soil Biol. Biochem. 2000, 32, 1183–1190. [Google Scholar] [CrossRef]
- Schutter, M.E.; Dick, R.P. Comparison of fatty acid methyl ester (FAME) methods for characterizing microbial communities. Soil Sci. Soc. Am. J. 2000, 64, 1659–1668. [Google Scholar] [CrossRef]
- Gardner, T.; Acosta-Martinez, V.; Senwo, Z.; Dowd, S.E. Soil Rhizosphere Microbial Communities and Enzyme Activities under Organic Farming in Alabama. Diversity 2011, 3, 308–328. [Google Scholar] [CrossRef]
- Zelles, L. Fatty Acid Patterns of Microbial Phospholipids and Lipopolysaccharides. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 80–93. [Google Scholar]
- Zelles, L. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 1997, 35, 275–294. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of microbial phospholipids and lipopolysaccharides in the characterization of microbial communities in soil: A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Oksanen, F.G.B.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.3-0; 2015; Available online: https://www.researchgate.net/publication/311493644_Vegan_commity_ecology_package_R_package_version_23-0 (accessed on 24 July 2021).
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. Available online: http://www.jstatsoft.org/v48/i02/ (accessed on 24 July 2021). [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
- You, Y.; Wang, J.; Huang, X.; Tang, Z.; Liu, S.; Sun, O. Relating microbial community structure to functioning in forest soil organic carbon transformation and turnover. Ecol. Evol. 2014, 4, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.J.; Hester, A.J.; Campbell, C.D.; Chapman, S.J.; Cameron, C.M.; Hewison, R.L. Is vegetation composition or soil chemistry the best predictor of the soil microbial community? Plant Soil 2010, 333, 417–430. [Google Scholar] [CrossRef]
- Garau, G.; Morillas, L.; Robles, J.; Castaldi, P.; Mangia, N.P.; Spano, D.; Mereu, S. Effect of monospecific and mixed Mediterranean tree plantations on soil microbial community and biochemical functioning. Appl. Soil Ecol. 2019, 140, 78–88. [Google Scholar] [CrossRef]
- Wu, Y.T.; Gutknecht, J.; Nadrowski, K.; Geißler, C.; Kühn, P.; Scholten, T.; Both, S.; Erfmeier, A.; Böhnke, M.; Bruelheide, H.; et al. Relationships between soil microorganisms, plant communities and soil characteristics in Chinese subtropical forests. Ecosystems 2012, 15, 624–636. [Google Scholar] [CrossRef]
- Chemidlin Prevost-Boure, N.; Maron, P.A.; Ranjard, L.; Nowak, V.; Dufrene, E.; Damesin, C.; Soudani, K.; Lata, J.-C. Seasonal dynamics of the bacterial community in forest soils under different quantities of leaf litter. Appl. Soil Ecol. 2011, 47, 14–23. [Google Scholar] [CrossRef]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Schimel, J.P.; VanCleve, K.; Cates, R.G.; Clausen, T.P.; Reichardt, P.B. Effects of balsam poplar (Populus balsamifera) tannins and low molecular weight phenolics on microbial activity in taiga floodplain soil: Implications for changes in N cycling during succession. Can. J. Bot. 1996, 74, 84–90. [Google Scholar] [CrossRef]
- Schroth, G. Decomposition and nutrient supply from biomass. In Trees, Crops and Soil Fertility: Concepts and Research Methods; Schroth, G., Sinclair, F.L., Eds.; CAB International: Wallingford, UK, 2003; pp. 131–150. [Google Scholar]
- Fox, J.C.; Yosi, C.K.; Nimiago, P.; Oavika, F.; Pokana, J.N.; Lavong, K.; Keenan, R.J. Assessment of Aboveground Carbon in Primary and Selectively Harvested Tropical Forest in Papua New Guinea. Biotropica 2010, 42, 410–419. [Google Scholar] [CrossRef]
- Witherspoon, A.; Thevathasan, N.V.; Gordon, A.M.; Voroney, R.P. Carbon sequestration potential of five tree species in a 25-year-old temperate tree-based intercropping system in southern Ontario, Canada. Agrofor. Syst. 2014, 88, 631–643. [Google Scholar] [CrossRef]
- Mungai, N.W.; Motavalli, P.P. Litter quality effects on soil carbon and nitrogen dynamics in temperate alley cropping systems. Appl. Soil Ecol. 2006, 31, 32–42. [Google Scholar] [CrossRef]
- Rousk, J.; Frey, S.D.; Bååth, E. Temperature adaptation of bacterial communities in experimentally warmed forest soils. Glob. Chang. Biol. 2012, 18, 3252–3258. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E. Groth of saprotrophic fungi and bacteria in soil. FEMS Microbiol. Ecol. 2011, 78, 17–30. [Google Scholar] [CrossRef]
- Bauhus, J.; Pare, D.; Cote, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Finzi, A.C.; Van Breemen, N.; Canham, C.D. Canopy tree–soil interactions within temperate forests: Species effects on soil carbon and nitrogen. Ecol. Appl. 1998, 8, 440–446. [Google Scholar]
- Kraus, T.E.C.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems—A review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.; Ferre, C.; Carderelli, E.; Montagnani, C.; Bogliani, G.; Citterio, S.; Comolli, R. Comparing negative impacts of Prunus serotina, Quercus rubra and Robinia pseudoacacia on native forest ecosystems. Forest 2019, 10, 842. [Google Scholar] [CrossRef]
- Stanek, M.; Stefanowicz, A.M. Invasive Quercus rubra negatively affected soil microbial communities relative to native Quercus robur in a semi-natural forest. Sci. Total Environ. 2019, 696, 133977. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Bastida, F.; Nicolás, C.; Moreno, J.L.; del Cerro, A.; Andrés, M. Influence of forest cover and herbaceous vegetation on the microbiological and biochemical properties of soil under Mediterranean humid climate. Eur. J. Soil Biol. 2010, 46, 273–279. [Google Scholar] [CrossRef]
- Mincheva, T.; Barni, E.; Varese, G.C.; Brusa, G.; Cerabolini, B.; Siniscalco, C. Litter quality, decomposition rates and saprotrophic mycoflora in Fallopia japonica (Houtt.) Ronse Decraene and in adjacent native grassland vegetation. Acta Oecolog. 2014, 54, 29–35. [Google Scholar] [CrossRef]
- Berg, B.; Johansson, M.B.; Anta, R.C.D.; Escudero, A.; Gärdenäs, A.; Laskowski, R.; Santo, A.V.D. The chemical composition of newly shed needle litter of Scots pine and some other pine species in a climatic transect. X Long-term decomposition in a Scots pine forest. Can. J. Bot. 1995, 73, 1423–1435. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Veresoglou, S.D.; Chen, B.D.; Rillig, M.C. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol. Biochem. 2012, 46, 53–62. [Google Scholar] [CrossRef]
- Carteron, A.; Beiges, M.; Joly, S.; Turner, B.L.; Laliberte, E. Temperate forests dominated by Arbuscular or Ectomycorrhizal fungi are characterized by strong shifts from saprotrophic to Mycorrhizal fungi with increasing soil depth. Microb. Ecol. 2020, 82, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Cates, L.L.; Dietze, M.C.; Bhatnagar, J.M. Spatial vs. temporal controls over soil fungal community similarity at continental and global scales. ISME J. 2019, 13, 2082–2093. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wheeler, E.; Phillips, R. Root-induced changes in nutrient in forest depend on exudation rates. Soil Biol. Biochem. 2014, 78, 213–321. [Google Scholar] [CrossRef]
- McGuire, K.L.; Allison, S.D.; Fierer, N.; Treseder, K.K. Ectomycorrhizal-Dominated Boreal and Tropical Forests Have Distinct Fungal Communities, but Analogous Spatial Patterns across Soil Horizons. PLoS ONE 2013, 8, e68278. [Google Scholar] [CrossRef]
- Yokobe, T.; Hyodo, F.; Tokuchi, N. Seasonal effects on microbial community structure and nitrogen dynamics in temperate forest soil. Forest 2018, 9, 153. [Google Scholar] [CrossRef]
- Voříšková, J.; Brabcova, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Pettersson, M.; Bååth, E. Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol. Ecol. 2005, 52, 49–58. [Google Scholar]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef]
- Baldrian, P. Microbial activity and the dynamics of ecosystem processes in forest soils. Curr. Opin. Microbiol. 2017, 37, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Söderström, B.E. Seasonal fluctuations of active fungal biomass in horizons of a podzolized pine-forest soil in Central Sweden. Soil Biol. Biochem. 1979, 11, 149–154. [Google Scholar] [CrossRef]
- Talbot, J.M.; Bruns, T.D.; Smit, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vilgalys, R.; Peay, K.G. Independent roles of ectomycorrhizal and saprotrophic communities in soil organic matter decomposition. Soil Biol. Biochem. 2013, 57, 282–2091. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Siccama, T.G. Plant-soil microbial interactions in a northern hardwood forest. Ecology 2001, 82, 965–978. [Google Scholar]
- Rastin, N.; Rosenplänter, K.; Hüttermann, A. Seasonal variation of enzyme activity and their dependence on certain soil factors in a beech forest soil. Soil Biol. Biochem. 1988, 20, 637–642. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought decreases soil enzyme activity in a Mediterranean Quercus ilex L. forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol. Biochem. 2013, 54, 60–68. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Grayston, S.J.; Prescott, C.E. Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Biol. Biochem. 2005, 37, 1157–1167. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry 2013, 114, 313–326. [Google Scholar] [CrossRef]
- Manzoni, S.; Katul, G. Invariant soil water potential at zero microbial respiration explained by hydrological discontinuity in dry soils. Geophys. Res. Lett. 2014, 41, 7151–7158. [Google Scholar] [CrossRef]
- Turner, B.L. Variation in pH optima of hydrolytic enzyme activities in tropical rain in forest soil. Appl. Environ. Microbiol. 2010, 76, 6485–6493. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Mackensen, J.; Bauhus, J.; Webber, E. Decomposition rates of coarse woody debris—A review with particular emphasis on Australian tree species. Aust. J. Bot. 2003, 51, 27–37. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Season | Plant | TOC (%) | TC (%) | TN (%) | C:N | pH | Soil Moisture (%) |
|---|---|---|---|---|---|---|---|
| 0–10 cm | |||||||
| Autumn | BW | 1.59 (0.34) | 1.84 (0.29) a | 0.15 (0.02) a | 11.87 (1.44) | 6.65 (0.07) a | 19.75 (3.17) |
| RO | 1.13 (0.23) | 1.19 (0.23) b | 0.11 (0.01) b | 10.74 (0.99) | 5.80 (0.26) b | 17.57 (1.82) | |
| DB | 1.40 (0.12) | 1.57 (0.07) ab | 0.13 (0.00) ab | 11.36 (0.18) | 6.42 (0.42) a | 20.48 (2.14) | |
| Winter | BW | 1.42 (0.26) | 1.72 (0.33) | 0.13 (0.03) | 12.97 (1.74) | 6.56 (0.19) | 21.98 (1.62) |
| RO | 1.10 (0.27) | 1.30 (0.17) | 0.11 (0.02) | 11.20 (0.47) | 6.09 (0.38) | 20.42 (2.43) | |
| DB | 1.38 (0.18) | 1.59 (0.26) | 0.14 (0.02) | 11.09 (0.48) | 6.26 (0.50) | 22.16 (2.34) | |
| Spring | BW | 1.70 (0.29) | 1.91 (0.24) | 0.15 (0.02) | 12.88 (1.30) | 6.15 (0.06) ab | 18.71 (1.70) |
| RO | 1.45 (0.29) | 1.55 (0.31) | 0.12 (0.03) | 12.41 (0.77) | 5.85 (0.25) b | 16.82 (1.76) | |
| DB | 1.58 (0.16) | 1.73 (0.10) | 0.15 (0.01) | 11.09 (0.50) | 7.09 (0.70) a | 18.31 (2.28) | |
| Summer | BW | 1.99 (0.34) a | 2.30 (0.30) a | 0.18 (0.03) a | 12.92 (1.33) | 6.42 (0.05) a | 7.00 (2.98) |
| RO | 1.18 (0.10) b | 1.30 (0.05) b | 0.10 (0.01) b | 12.48 (0.85) | 5.81 (0.20) b | 4.91 (0.88) | |
| DB | 1.57 (0.03) ab | 1.78 (0.05) b | 0.14 (0.00) ab | 12.15 (0.60) | 6.32 (0.32) a | 7.86 (1.39) | |
| 10–20 cm | |||||||
| Autumn | BW | 1.13 (0.22) a | 1.38 (0.27) a | 0.12 (0.02) | 11.14 (1.37) | 6.51 (0.10) a | 17.64 (3.92) |
| RO | 0.80 (0.21) b | 0.93 (0.16) b | 0.09 (0.01) | 10.36 (0.52) | 5.97 (0.30) b | 18.81 (0.94) | |
| DB | 0.74 (0.17) b | 0.93 (0.14) b | 0.09 (0.01) | 10.38 (0.09) | 6.30 (0.25) ab | 17.76 (2.53 | |
| Winter | BW | 1.25 (0.34) | 1.38 (0.40) | 0.10 (0.03) | 13.20 (2.94) | 6.45 (0.18) | 19.91 (1.94) |
| RO | 0.85 (0.20) | 0.94 (0.17) | 0.08 (0.02) | 11.76 (1.29) | 6.32 (0.35) | 18.05 (1.91) | |
| DB | 0.88 (0.15) | 1.02 (0.18) | 0.09 (0.01) | 10.96 (0.70) | 6.17 (0.16) | 16.71 (2.69) | |
| Spring | BW | 1.40 (0.31) | 1.53 (0.35) | 0.12 (0.02) | 12.16 (2.21) | 6.23 (0.07) ab | 16.52 (2.58) a |
| RO | 0.87 (0.18) | 0.98 (0.12) | 0.09 (0.01) | 10.78 (0.88) | 5.95 (0.27) b | 11.44 (1.14) b | |
| DB | 1.03 (0.22) | 1.32 (0.34) | 0.13 (0.03) | 10.10 (0.34) | 6.78 (0.48) a | 14.82 (1.39) ab | |
| Summer | BW | 1.60 (0.29) a | 1.82 (0.38) a | 0.15 (0.02) a | 12.13 (2.10) | 6.34 (0.16) ab | 9.00 (2.52) |
| RO | 0.85 (0.24) b | 0.96 (0.19) b | 0.08 (0.02) b | 11.31 (0.38) | 5.90 (0.33) b | 8.12 (3.39) | |
| DB | 1.00 (0.14) b | 1.24 (0.21) ab | 0.11 (0.01) ab | 10.92 (0.84) | 7.16 (0.49) a | 7.89 (0.93) | |
| Season | Area | FAME (ttl) | Bacteria (ttl) | Fungi (ttl) | Gram (+) | Gram (–) | ATB | AMF | SF |
|---|---|---|---|---|---|---|---|---|---|
| 0–10 cm | |||||||||
| Autumn | BW | 1592.75 (297.72) | 227.32 (58.43) | 439.94 (191.34) | 113.89 (28.11) | 44.99 (13.58) | 68.44 (16.88) | 148.01 (35.49) | 291.92 (164.47) |
| RO | 1108.13 (141.23) | 167.53 (25.50) | 262.99 (69.48) | 78.91 (10.48) | 37.13 (7.95) | 51.47 (7.45) | 55.92 (32.81) | 207.07 (16.84) | |
| DB | 1529.03 (233.53) | 245.27 (36.99) | 419.43 (69.48) | 117.65 (17.88) | 49.59 (8.10) | 78.01 (11.52) | 101.10 (35.40) | 318.32 (83.06) | |
| Winter | BW | 1509.69 (395.57) | 193.67 (52.60) | 401.74 (136.79) | 103.68 (26.40) | 33.86 (9.28) | 56.12 (17.39) | 124.94 (36.66) | 276.79 (105.69) |
| RO | 1041.50 (221.51) | 153.049 (44.14) | 282.94 (40.30) | 76.90 (21.77) | 32.25 (11.72) | 43.88 (11.34) | 35.02 (5.49) | 247.92 (45.56) | |
| DB | 1025.96 (110.88) | 209.71 (15.02) | 401.65 (88.27) | 104.0 (11.06) | 40.57 (2.26) | 64.33 (4.37) | 99.43 (58.27) | 302.22 (73.37) | |
| Spring | BW | 1776.09 (242.53) | 210.71 (38.12) | 257.02 (29.66) | 108.86 (18.19) | 41.75 (8.72) | 60.09 (11.71) | 81.39 (9.84) | 175.62 (28.76) |
| RO | 1025.04 (84.14) | 194.52 (10.94) | 270.11 (81.35) | 82.88 (18.88) | 46.64 (3.09) | 64.99 (11.45) | 54.21 (42.33) | 215.89 (96.03) | |
| DB | 1025.96 (84.14) | 201.70 (24.40) | 173.94 (68.83) | 97.77 (13.26) | 41.51 (5.78) | 62.40 (7.37) | 51.42 (23.34) | 122.51 (45.49) | |
| Summer | BW | 1629.21 (201.35) | 290.49 (72.05) | 408.80 (70.20) | 145.20 (18.19) | 59.67 (16.68) | 85.61 (21.65) | 165.52 (47.61) | 243.37 (31.87) |
| RO | 1126.96 (218.54) | 184.48 (22.56) | 303.60 (91.04) | 88.56 (11.62) | 51.21 (4.21) | 54.70 (6.92) | 59.91 (34.25) | 243.69 (57.16) | |
| DB | 1776.09 (115.69) | 289.40 (66.99) | 499.87 (175.51) | 144.76 (38.57) | 57.07 (9.75) | 88.57 (18.67) | 186.86 (96.74) | 313.01 (78.79) | |
| ANOVA | TS | 0.0056 | 0.0141 | n.s. | 0.0019 | n.s. | 0.0272 | 5.56 × 10−5 | n.s. |
| Season | n.s. | 0.0173 | 0.0306 | 0.03114 | 0.007 | 0.0215 | 0.0107 | n.s. | |
| TS × S | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |
| 10–20 cm | |||||||||
| Autumn | BW | 1123.43 (378.36) | 165.71 (55.88) | 301.62 (153.66) | 80.08 (26.48) | 35.36 (13.28) | 50.25 (16.32) | 110.74 (37.24) | 190.88 (134.41) |
| RO | 585.53 (173.74) | 102.18 (31.99) | 135.51 (53.87) | 47.42 (16.19) | 22.91 (6.28) | 31.84 (10.25) | 29.22 (18.99) | 106.28 (44.10) | |
| DB | 1372.21 (510.28) | 208.01 (71.17) | 408.54 (174.40) | 97.80 (32.45) | 42.74 (15.42) | 67.45 (23.33) | 95.69 (11.45) | 312.84 (176.97) | |
| Winter | BW | 820.62 (124.26) | 117.71 (23.94) | 186.87 (63.32) | 58.79 (10.22) | 24.16 (4.85) | 34.75 (9.15) | 78.04 (26.31) | 108.83 (38.48) |
| RO | 432.96 (113.79) | 82.80 (34.02) | 186.62 (63.32) | 37.70 (14.04) | 18.57 (8.91) | 26.53 (11.26) | 21.85 (12.23) | 84.77 (17.29) | |
| DB | 863.15 (156.39) | 150.65 (23.34) | 106.63 (12.67) | 73.56 (12.80) | 29.52 (3.90) | 47.57 (7.13) | 68.24 (20.78) | 166.96 (55.75) | |
| Spring | BW | 746.97 (132.8) | 143.51 (52.06) | 235.20 (38.27) | 68.86 (23.25) | 30.45 (11.62) | 44.18 (17.43) | 58.57 (19.82) | 74.14 (21.23) |
| RO | 602.13 (161.46) | 117.08 (30.11) | 135.13 (42.32) | 52.73 (12.71) | 27.61 (7.20) | 36.73 (10.35) | 21.69 (9.44) | 113.45 (42.52) | |
| DB | 620.19 (80.52) | 145.58 (27.40) | 130.98 (13.84) | 66.52 (12.99) | 31.063 (5.92) | 48.00 (9.17) | 48.07 (12.81) | 82.89 (13.98) | |
| Summer | BW | 1063.77 (181.78) | 178.27 (15.68) | 276.15 (85.57) | 85.57 (7.35) | 37.45 (4.27) | 55.21 (6.96) | 122.16 (50.85) | 153.99 (45.91) |
| RO | 704.48 (207.05) | 143.45 (53.16) | 166.73 (49.01) | 66.02 (25.38) | 32.21 (11.59) | 45.21 (16.26) | 54.64 (41.90) | 112.09 (28.29) | |
| DB | 967.64 (229.77) | 162.77 (21.59) | 278.17 (91.30) | 76.62 (13.58) | 33.40 (2.23) | 52.51 (5.91) | 118.70 (57.07) | 159.46 (35.62) | |
| ANOVA | TS | 0.0018 | 0.023 | 0.0209 | 0.0073 | n.s. | 0.0218 | 0.0002 | n.s. |
| Season | 0.027 | n.s. | 0.0145 | n.s. | n.s. | n.s. | 0.0079 | n.s. | |
| TS × S | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Variable | FAME (ttl) | Bacteria (ttl) | Fungi (ttl) | Gram (+) | Gram (–) | AMY | AMF | SF | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp | 0.03 | 0.23 | * | −0.07 | 0.18 | 0.30 | ** | 0.24 | * | 0.02 | −0.10 | ||
| Soil Moisture | −0.27 | −0.25 | −0.08 | −0.30 | * | −0.18 | −0.16 | 0.07 | −0.14 | ||||
| pH | 0.23 | 0.26 | 0.18 | 0.35 | * | 0.01 | 0.23 | 0.5 | *** | −0.04 | |||
| TOC | 0.15 | 0.46 | *** | −0.05 | 0.5 | *** | 0.36 | * | 0.36 | * | 0.1 | −0.13 | |
| TC | 0.28 | 0.37 | ** | 0.13 | 0.46 | *** | 0.21 | 0.25 | 0.26 | 0.02 | |||
| TN | 0.22 | 0.56 | *** | 0.04 | 0.59 | *** | 0.43 | ** | 0.46 | *** | 0.2 | −0.05 | |
| C:N | 0.17 | −0.25 | 0.18 | −0.13 | −0.35 | ** | −0.33 | * | 0.15 | 0.15 | |||
| BG | 0.05 | 0.33 | * | −0.02 | 0.38 | ** | 0.15 | 0.29 | * | 0.43 | *** | −0.26 | |
| NAG | −0.13 | 0.01 | −0.02 | 0.02 | −0.07 | 0.02 | 0.14 | −0.11 | |||||
| PME | −0.02 | 0.26 | −0.07 | 0.21 | 0.27 | 0.26 | 0.09 | −0.15 | |||||
| AS | −0.02 | 0.33 | * | −0.11 | 0.34 | * | 0.26 | 0.29 | * | 0.18 | −0.23 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardner, T.G.; Frene, J.P.; Lawson, S.S.; Lue Sue, N.D.; Handy, J.; Crawford, R.H. The Impact of Tree Species on Microbial Community Structure and Soil Function on Forest Plantations in the Central Hardwoods Region (CHR). Forests 2023, 14, 859. https://doi.org/10.3390/f14050859
Gardner TG, Frene JP, Lawson SS, Lue Sue ND, Handy J, Crawford RH. The Impact of Tree Species on Microbial Community Structure and Soil Function on Forest Plantations in the Central Hardwoods Region (CHR). Forests. 2023; 14(5):859. https://doi.org/10.3390/f14050859
Chicago/Turabian StyleGardner, Terrence G., Juan P. Frene, Shaneka S. Lawson, Niall D. Lue Sue, Jeffery Handy, and Ralph H. Crawford. 2023. "The Impact of Tree Species on Microbial Community Structure and Soil Function on Forest Plantations in the Central Hardwoods Region (CHR)" Forests 14, no. 5: 859. https://doi.org/10.3390/f14050859
APA StyleGardner, T. G., Frene, J. P., Lawson, S. S., Lue Sue, N. D., Handy, J., & Crawford, R. H. (2023). The Impact of Tree Species on Microbial Community Structure and Soil Function on Forest Plantations in the Central Hardwoods Region (CHR). Forests, 14(5), 859. https://doi.org/10.3390/f14050859