Correlation between Changes in Soil Properties and Microbial Diversity Driven by Different Management in Artificial Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) Plantations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Overview of the Survey Area
2.2. Sample Plot Setting and Forest Stand Growth Survey
2.3. Soil Sample Collection
2.4. Investigation of Physical and Chemical Properties and the Enzyme Activities of Soil Samples
2.5. Soil Microbial Community Diversity Analysis
- (1)
- Removing reads containing more than 10% of unknown uncleotides (N);
- (2)
- Removing reads containing less than 50% of bases with quality (Q-value) >20.
- (1)
- Break raw tags from the first low-quality base site where the number of bases in the continuous low-quality value (the default quality threshold is ≤3) reaches the set length (the default length is 3 bp);
- (2)
- Then, filter tags whose continuous high-quality base length is less than 75% of the tag length.
2.6. Taxonomy Annotation and Community Composition, Diversity, Function and Environmental Factor Analysis
3. Results
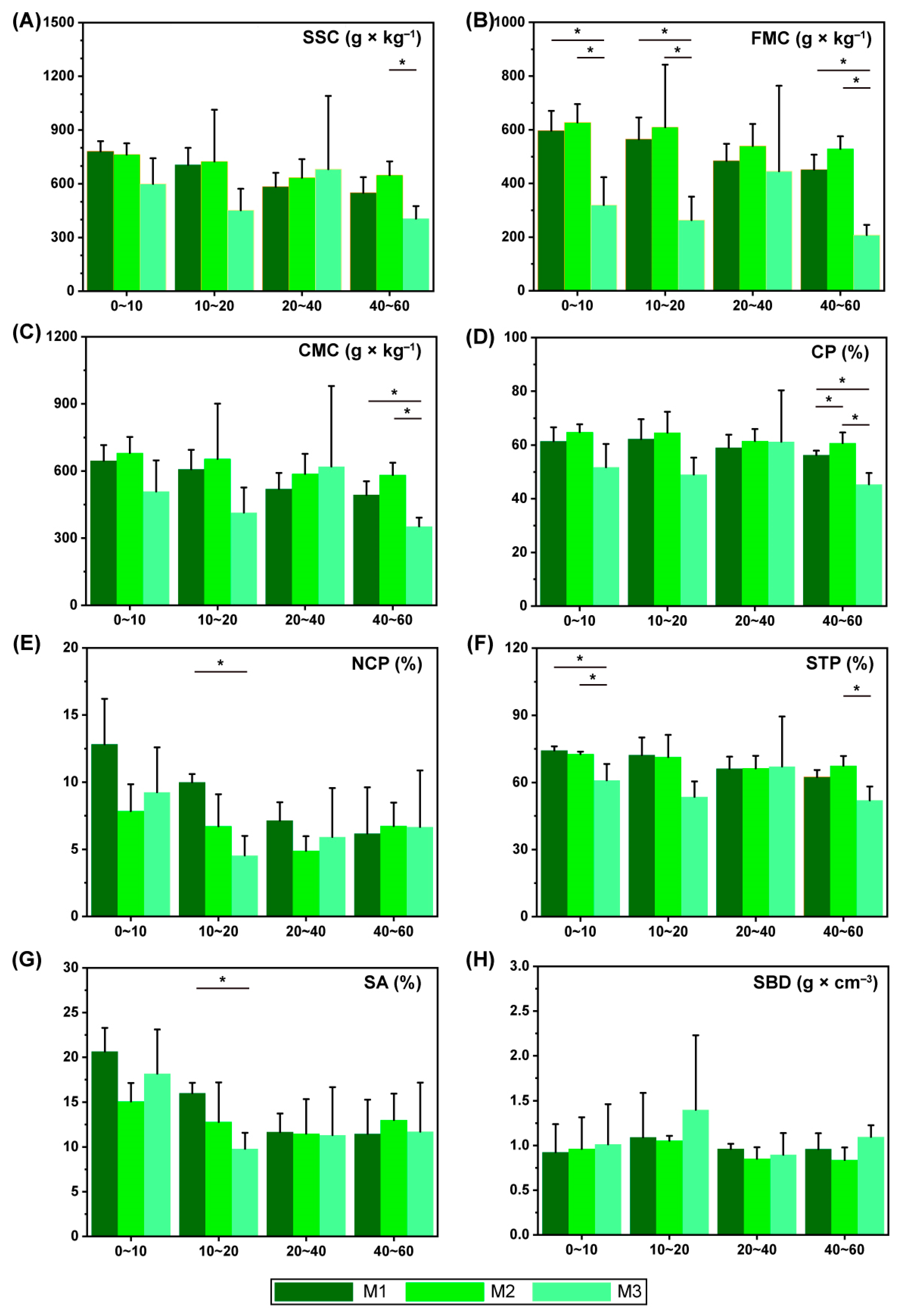
3.1. Effect of Different Models on Soil Physical Status Changes
3.2. Effect of Different Models on Soil Nutrient Content Changes
3.3. Effect of Different Models on Soil Enzyme Activities Changes
3.4. Diversity, Function, and Correlation with Environmental Factors of Soil Bacteria
3.5. Diversity, Function, and Correlation to Environmental Factors of Soil Fungi
4. Discussion
4.1. Soil Properties Change after Conversion of the Artificial Plantation Ecosystem
4.2. Role of Manual Intervention in Succession of the Soil Microorganism Population Structure
4.3. Correlation between Soil Properties and Microbial Composition
4.4. Limitation and Suggession for Sustainable C. lanceolata Plantation Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giles-Vernick, T. Leaving a person behind: History, personhood, and struggles over forest resources in the Sangha Basin of Equatorial Africa. Int. J. Afr. Hist. Stud. 1999, 32, 311–338. [Google Scholar] [CrossRef] [PubMed]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, M.; Li, X.; Liu, Z.; Ming, A.; Lan, H.; Ye, S. The Abundance and Structure of Deadwood: A Comparison of Mixed and Thinned Chinese Fir Plantations. Front. Plant Sci. 2021, 12, 614695. [Google Scholar] [CrossRef]
- Huang, H.-H.; Xu, L.-L.; Tong, Z.-K.; Lin, E.-P.; Liu, Q.-P.; Cheng, L.-J.; Zhu, M.-Y. De novo characterization of the Chinese fir (Cunninghamia lanceolata) transcriptome and analysis of candidate genes involved in cellulose and lignin biosynthesis. BMC Genom. 2012, 13, 648. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wu, H.; Ouyang, S.; Chen, L.; Fang, X.; Peng, C.; Liu, S.; Xiao, W.; Xiang, W. Ecosystem service multifunctionality of Chinese fir plantations differing in stand age and implications for sustainable management. Sci. Total Environ. 2021, 788, 147791. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, X.; Chhin, S.; Zhang, J. A Bimodal Pattern and Age-Related Growth of Intra-Annual Wood Cell Development of Chinese Fir in Subtropical China. Front. Plant Sci. 2021, 12, 757438. [Google Scholar] [CrossRef]
- Zeng, Q.; Yu, X.; Wei, N.; Wu, Z.; Liu, Q.; Chen, N.; Zhao, W. Effect of Impregnation with Natural Shellac Polymer on the Mechanical Properties of Fast-Growing Chinese Fir. Polymers 2022, 14, 3871. [Google Scholar] [CrossRef]
- Cui, H.O.; Liu, M. Analysis on the results of the 9th national forest inventory. J. West China For. Sci. 2020, 49, 90–95, (Chinese with English Abstract). [Google Scholar] [CrossRef]
- Wang, Q.; Wang, J.; Li, Y.; Chen, D.; Ao, J.; Zhou, W.; Shen, D.; Li, Q.; Huang, Z.; Jiang, Y. Influence of nitrogen and phosphorus additions on N2-fixation activity, abundance, and composition of diazotrophic communities in a Chinese fir plantation. Sci. Total Environ. 2018, 619–620, 1530–1537. [Google Scholar] [CrossRef]
- Li, R.; Yu, D.; Zhang, Y.; Han, J.; Zhang, W.; Yang, Q.; Gessler, A.; Li, M.-H.; Xu, M.; Guan, X.; et al. Investment of needle nitrogen to photosynthesis controls the nonlinear productivity response of young Chinese fir trees to nitrogen deposition. Sci. Total Environ. 2022, 840, 156537. [Google Scholar] [CrossRef]
- Yang, W.D.; Liu, J.S.; Li, H.Y.; Zhang, X.L.; Qi, Y.Z. Inhibition of the growth of Alexandrium tamarense by algicidal substances in Chinese fir (Cunninghamia lanceolata). Bull. Environ. Contam. Toxicol. 2009, 83, 537–541. [Google Scholar] [CrossRef]
- Huang, Z.; Liao, L.; Wang, S.; Lui, Y. Dynamics of phenolics content of Chinese fir stump-roots and the rhizosphere soil and it’s allelopathy. J. Appl. Ecol. 2000, 11, 190–192, (Chinese with English Abstract). [Google Scholar]
- Pachepsky, E.; Taylor, T.; Jones, S. Mutualism promotes diversity and stability in a simple artificial ecosystem. Artif. Life 2002, 8, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Jennings, S.; Melin, F.; Blanchard, J.L.; Forster, R.M.; Dulvy, N.K.; Wilson, R.W. Global-scale predictions of community and ecosystem properties from simple ecological theory. Proc. Biol. Sci. R. Soc. 2008, 275, 1375–1383. [Google Scholar] [CrossRef]
- Fauteux, D.; Stien, A.; Yoccoz, N.G.; Fuglei, E.; Ims, R.A. Climate variability and density-dependent population dynamics: Lessons from a simple High Arctic ecosystem. Proc. Natl. Acad. Sci. USA 2021, 118, e2106635118. [Google Scholar] [CrossRef]
- Bai, Y.; Chen, S.; Shi, S.; Qi, M.; Liu, X.; Wang, H.; Wang, Y.; Jiang, C. Effects of different management approaches on the stoichiometric characteristics of soil C, N, and P in a mature Chinese fir plantation. Sci. Total Environ. 2020, 723, 137868. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; He, L.; Zhang, X.; Kong, D.; Chen, Z.; Lin, J.; Wang, C. Bioremediation of petroleum-contaminated saline soil by Acinetobacter baumannii and Talaromyces sp. and functional potential analysis using metagenomic sequencing. Environ. Pollut. 2022, 311, 119970. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, X.; Hu, Y.; Wang, P.; Saeed, S.; Sun, Y. Short-term effects of thinning on the development and communities of understory vegetation of Chinese fir plantations in Southeastern China. PeerJ 2020, 8, e8536. [Google Scholar] [CrossRef]
- Peng, C.; Song, Y.; Li, C.; Mei, T.; Wu, Z.; Shi, Y.; Zhou, Y.; Zhou, G. Growing in Mixed Stands Increased Leaf Photosynthesis and Physiological Stress Resistance in Moso Bamboo and Mature Chinese Fir Plantations. Front. Plant Sci. 2021, 12, 649204. [Google Scholar] [CrossRef]
- Li, J.; Li, L.; Arif, M.; Ding, D.; Hu, X.; Zheng, J.; Yuan, Z.; Li, C. Artificial Plantation Responses to Periodic Submergence in Massive Dam and Reservoir Riparian Zones: Changes in Soil Properties and Bacterial Community Characteristics. Biology 2021, 10, 819. [Google Scholar] [CrossRef]
- Xiang, W.; Xu, L.; Lei, P.; Ouyang, S.; Deng, X.; Chen, L.; Zeng, Y.; Hu, Y.; Zhao, Z.; Wu, H.; et al. Rotation age extension synergistically increases ecosystem carbon storage and timber production of Chinese fir plantations in southern China. J. Environ. Manag. 2022, 317, 115426. [Google Scholar] [CrossRef]
- Gentile, J.M.; Pluymers, D.; Plewa, M.J. Promutagen activation as a function of growth curve dynamics in the plant cell/microbe coincubation assay. Mutat. Res. 1986, 173, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.; Deacon, L.; Grinev, D.; Harris, J.; Ritz, K.; Singh, B.; Young, I. Microbial diversity affects self-organization of the soil-microbe system with consequences for function. J. R. Soc. Interface 2012, 9, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Noutoshi, Y. The function of the plant cell wall in plant-microbe interactions. Plant Physiol. Biochem. 2022, 192, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, J.; Zheng, J.; Liu, J.; Liu, S.; Lin, W.; Wu, C. Soil microbial community structure and catabolic activity are significantly degenerated in successive rotations of Chinese fir plantations. Sci. Rep. 2017, 7, 6691. [Google Scholar] [CrossRef]
- Seifert, J. Intensity of nitrification in soil as a function of time and soil moisture. Folia Microbiol. 1973, 18, 386–389. [Google Scholar] [CrossRef]
- Gleeson, D.; Mathes, F.; Farrell, M.; Leopold, M. Environmental drivers of soil microbial community structure and function at the Avon River Critical Zone Observatory. Sci. Total Environ. 2016, 571, 1407–1418. [Google Scholar] [CrossRef]
- Bobul’ska, L.; Espindola, S.P.; Coelho, M.A.; Ferreira, A.S. Impact of land use on soil function and bacterial community in the Brazilian savanna. An. Acad. Bras. Cienc. 2021, 93, e20201906. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Liu, Y.; Chen, H.; Hu, Y. Response of Bacterial and Fungal Soil Communities to Chinese Fir (Cunninghamia lanceolate) Long-Term Monoculture Plantations. Front. Microbiol. 2020, 11, 181. [Google Scholar] [CrossRef]
- Chen, J.; Deng, Z.; Jiang, Z.; Sun, J.; Meng, F.; Zuo, X.; Wu, L.; Cao, G.; Cao, S. Variations of rhizosphere and bulk soil microbial community in successive planting of Chinese fir (Cunninghamia lanceolata). Front. Plant Sci. 2022, 13, 954777. [Google Scholar] [CrossRef]
- Pan, B.; Xing, B.; Tao, S.; Liu, W.; Lin, X.; Xiao, Y.; Dai, H.; Zhang, X.; Zhang, Y.; Yuan, H. Effect of physical forms of soil organic matter on phenanthrene sorption. Chemosphere 2007, 68, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Wang, M.K.; Hu, G.; Weng, B. Seasonal Change in Microbial Diversity and Its Relationship with Soil Chemical Properties in an Orchard. PLoS ONE 2019, 14, e0215556. [Google Scholar] [CrossRef] [PubMed]
- Namaghi, H.H.; Karami, G.H.; Saadat, S. A study on chemical properties of groundwater and soil in ophiolitic rocks in Firuzabad, east of Shahrood, Iran: With emphasis to heavy metal contamination. Environ. Monit. Assess. 2011, 174, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Ankenbrand, M.J.; Keller, A.; Wolf, M.; Schultz, J.; Forster, F. ITS2 Database V: Twice as Much. Mol. Biol. Evol. 2015, 32, 3030–3032. [Google Scholar] [CrossRef]
- Qadir, M.; Hussain, A.; Shah, M.; Hamayun, M.; Iqbal, A.; Nadia. Enhancement of chromate phytoremediation and soil reclamation potential of Brassica campestris L. by Aspergillus niger. Environ. Sci. Pollut. Res. Int. 2022, 30, 9471–9482. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-3. 2017. Available online: http://cran.r-project.org (accessed on 1 January 2016).
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Aylin Kasa, N.; Sel, S.; Ozkan, B.C.; Bakirdere, S. Determination of palladium in soil samples by slotted quartz tube-flame atomic absorption spectrophotometry after vortex-assisted ligandless preconcentration with magnetic nanoparticle-based dispersive solid-phase microextraction. Environ. Monit. Assess. 2019, 191, 692. [Google Scholar] [CrossRef] [PubMed]
- Elmi, A.A.; Astatkie, T.; Madramootoo, C.; Gordon, R.; Burton, D. Assessment of denitrification gaseous end-products in the soil profile under two water table management practices using repeated measures analysis. J. Environ. Qual. 2005, 34, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Aihemaiti, A.; Gao, Y.; Liu, L.; Yang, G.; Han, S.; Jiang, J. Effects of liquid digestate on the valence state of vanadium in plant and soil and microbial community response. Environ. Pollut. 2020, 265, 114916. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; Blair, D.; Keith, H.; Lindenmayer, D. Modelling water yields in response to logging and Representative Climate Futures. Sci. Total Environ. 2019, 688, 890–902. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Z.; Niu, S.; Bai, Z. The reclaimed coal mine ecosystem diverges from the surrounding ecosystem and reaches a new self-sustaining state after 20-23 years of succession in the Loess Plateau area, China. Sci. Total Environ. 2020, 727, 138739. [Google Scholar] [CrossRef]
- Yang, X.J.; Deng, Y.Y.; Duan, L.D. Research on Restoration of Soil Moisture Regulation Function in the Harvested Chinese Fir Plantation after Reforestation. For. Res. 2016, 29, 261–267, (Chinese with English abstract). [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef]
- Luo, X.; Hou, E.; Zang, X.; Zhang, L.; Yi, Y.; Wen, D. Effects of elevated atmospheric CO2 and nitrogen deposition on leaf litter and soil carbon degrading enzyme activities in a Cd-contaminated environment: A mesocosm study. Sci. Total Environ. 2019, 671, 157–164. [Google Scholar] [CrossRef]
- Kuscu, I.S.K. Changing of soil properties and urease-catalase enzyme activity depending on plant type and shading. Environ. Monit. Assess. 2019, 191, 178. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Yao, J.; Cai, M.; Qian, Y.; Guo, Y.; Richnow, H.H.; Blake, R.E.; Doni, S.; Ceccanti, B. Effects of petroleum contamination on soil microbial numbers, metabolic activity and urease activity. Chemosphere 2012, 87, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- An, S.S.; Huang, Y.M.; Zheng, F.L. Urease Activity in the Loess Hilly Grassland Soil and its Relationship to Soil Property. Acta Agrestia Sin. 2005, 13, 233–237, (Chinese with English Abstract). [Google Scholar]
- Tsai, S.H.; Chen, Y.T.; Yang, Y.L.; Lee, B.Y.; Huang, C.J.; Chen, C.Y. The Potential Biocontrol Agent Paenibacillus polymyxa TP3 Produces Fusaricidin-Type Compounds Involved in the Antagonism Against Gray Mold Pathogen Botrytis cinerea. Phytopathology 2002, 112, 775–783. [Google Scholar] [CrossRef]
- Wang, C.; Huang, Y.; Yang, X.; Xue, W.; Zhang, X.; Zhang, Y.; Pang, J.; Liu, Y.; Liu, Z. Burkholderia sp. Y4 inhibits cadmium accumulation in rice by increasing essential nutrient uptake and preferentially absorbing cadmium. Chemosphere 2020, 252, 126603. [Google Scholar] [CrossRef]
- Hengge, N.N.; Mallinson, S.J.B.; Pason, P.; Lunin, V.V.; Alahuhta, M.; Chung, D.; Himmel, M.E.; Westpheling, J.; Bomble, Y.J. Characterization of the Biomass Degrading Enzyme GuxA from Acidothermus cellulolyticus. Int. J. Mol. Sci. 2022, 23, 6070. [Google Scholar] [CrossRef]
- Luo, L.; Wang, L.; Deng, L.; Mei, X.; Liu, Y.; Huang, H.; Du, F.; Zhu, S.; Yang, M. Enrichment of Burkholderia in the Rhizosphere by Autotoxic Ginsenosides to Alleviate Negative Plant-Soil Feedback. Microbiol. Spectr. 2021, 9, e0140021. [Google Scholar] [CrossRef]
- Kong, C.H.; Chen, L.C.; Xu, X.H.; Wang, P.; Wang, S.L. Allelochemicals and activities in a replanted Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) tree ecosystem. J. Agric. Food Chem. 2008, 56, 11734–11739. [Google Scholar] [CrossRef]
- Alsowayeh, N.; Albutti, A. Designing a novel chimeric multi-epitope vaccine against Burkholderia pseudomallei, a causative agent of melioidosis. Front. Med. 2022, 9, 945938. [Google Scholar] [CrossRef]
- Haq, F.U.; Imran, M.; Saleem, S.; Waheed, Y. Antibacterial Activity of Different Extracts of Ascomata of Morchella conica and M. esculenta (Ascomycota) against Salmonella Species. Int. J. Med. Mushrooms 2022, 24, 85–95. [Google Scholar] [CrossRef]
- Xu, Z.; Ma, L.; Zhao, B.; Li, Y.; Chen, Y.; Deng, Y.; Wang, Y. Humification process enhancement through relative abundance promotion of Talaromyces and Coprinopsis by inoculated Phanerochaete chrysosporium during the secondary fermentation of composting. Environ. Sci. Pollut. Res. Int. 2022, 30, 9060–9065. [Google Scholar] [CrossRef]
- Shi, J.; Yang, S.; Yu, L.; Fu, B.; Liu, H. The pelletization characteristics of the filamentous fungi Talaromyces flavus S1 in sludge and the improvement for sludge dewatering. Acta Sci. Circumstantiae 2017, 37, 3672–3678. [Google Scholar]
- Sun, X.-R.; Xu, M.-Y.; Kong, W.-L.; Wu, F.; Zhang, Y.; Xie, X.-L.; Li, D.-W.; Wu, X.-Q. Fine Identification and Classification of a Novel Beneficial Talaromyces Fungal Species from Masson Pine Rhizosphere Soil. J. Fungi 2022, 8, 155. [Google Scholar] [CrossRef]
- Liu, M.; Gan, B.; Li, Q.; Xiao, W.; Song, X. Effects of Nitrogen and Phosphorus Addition on Soil Extracellular Enzyme Activity and Stoichiometry in Chinese Fir (Cunninghamia lanceolata) Forests. Front. Plant Sci. 2022, 13, 834184. [Google Scholar] [CrossRef]
- Nord, C.; Levenfors, J.J.; Bjerketorp, J.; Guss, B.; Oberg, B.; Brogerg, A. Tetramic acid based alkaloids from Aspergillus amoenus Roberg strain UP197 - antibiotic properties and new pyranterreones. Nat. Prod. Res. 2022, 36, 967–973. [Google Scholar] [CrossRef]
- Li, Y.; Shao, J.; Fu, Y.; Chen, Y.; Wang, H.; Xu, Z.; Feng, H.; Xun, W.; Liu, Y.; Zhang, N.; et al. The volatile cedrene from Trichoderma guizhouense modulates Arabidopsis root development through auxin transport and signalling. Plant Cell Environ. 2022, 45, 969–984. [Google Scholar] [CrossRef]
- Agbessenou, A.; Akutse, K.S.; Yusuf, A.A.; Khamis, F.M. The Endophyte Trichoderma asperellum M2RT4 Induces the Systemic Release of Methyl Salicylate and (Z)-jasmone in Tomato Plant Affecting Host Location and Herbivory of Tuta absoluta. Front. Plant Sci. 2022, 13, 860309. [Google Scholar] [CrossRef]
- Tian, Y.H.; Peng, H.Y.; Wang, D.H.; Li, X.F.; He, B.L.; Gao, K.X. Biocontrol potential of Talaromyces purpurogenus and its regulation on soil microbial community. Chin. J. Appl. Ecol. 2020, 31, 3255–3266, (Chinese with English abstract). [Google Scholar] [CrossRef]
- Janicki, T.; Krupinski, M.; Dlugonski, J. Degradation and toxicity reduction of the endocrine disruptors nonylphenol, 4-tert-octylphenol and 4-cumylphenol by the non-ligninolytic fungus Umbelopsis isabellina. Bioresour. Technol. 2016, 200, 223–229. [Google Scholar] [CrossRef]
- Nykiel-Szymanska, J.; Stolarek, P.; Bernat, P. Elimination and detoxification of 2,4-D by Umbelopsis isabellina with the involvement of cytochrome P450. Environ. Sci. Pollut. Res. Int. 2018, 25, 2738–2743. [Google Scholar] [CrossRef]
- Wang, L.; Qin, D.; Zhang, K.; Huang, Q.; Liu, S.; Han, M.J.; Dong, J.Y. Metabolites from the co-culture of nigranoic acid and Umbelopsis dimorpha SWUKD3.1410, an endophytic fungus from Kadsura angustifolia. Nat. Prod. Res. 2017, 31, 1414–1421. [Google Scholar] [CrossRef]
- Qin, D.; Wang, L.; Han, M.; Wang, J.; Song, H.; Yan, X.; Duan, X.; Dong, J. Effects of an Endophytic Fungus Umbelopsis dimorpha on the Secondary Metabolites of Host-Plant Kadsura angustifolia. Front. Microbiol. 2018, 9, 2845. [Google Scholar] [CrossRef]
- Aliyu, H.; Gorte, O.; Neumann, A.; Ochsenreither, K. Global Transcriptome Profile of the Oleaginous Yeast Saitozyma podzolica DSM 27192 Cultivated in Glucose and Xylose. J. Fungi 2021, 7, 758. [Google Scholar] [CrossRef]
- Cui, J.; Zhu, Z.K.; Xu, X.L.; Liu, S.; Ge, T. Carbon and nitrogen recycling from microbial necromass to cope with C:N stoichiometric imbalance by priming. Soil Biol. Biochem. 2020, 142, 107720. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Liu, H.Y.; Zhang, Y.Q.; Yu, L.Y. Soil pH is a Key Determinant of Soil Fungal Community Composition in the Ny-Ålesund Region, Svalbard (High Arctic). Front. Mocrobiol. 2016, 7, 227. [Google Scholar] [CrossRef]
- Fu, H.; Ding, D.; Sui, Y.; Zhang, H.; Hu, N.; Li, F.; Dai, Z.; Li, G.; Ye, Y.; Wang, Y. Transport of uranium(VI) in red soil in South China: Influence of initial pH and carbonate concentration. Environ. Sci. Pollut. Res. Int. 2019, 26, 37125–37136. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Z.; Xue, Y. Study on the new dynamics and driving factors of soil pH in the red soil, hilly region of South China. Environ. Monit. Assess. 2021, 193, 304. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, G.; Wu, L.; Qu, X.; Dai, L.; Ye, Y.; Xu, S.; Wu, C.; Chen, Y. Correlation between Changes in Soil Properties and Microbial Diversity Driven by Different Management in Artificial Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) Plantations. Forests 2023, 14, 877. https://doi.org/10.3390/f14050877
Cao G, Wu L, Qu X, Dai L, Ye Y, Xu S, Wu C, Chen Y. Correlation between Changes in Soil Properties and Microbial Diversity Driven by Different Management in Artificial Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) Plantations. Forests. 2023; 14(5):877. https://doi.org/10.3390/f14050877
Chicago/Turabian StyleCao, Guangqiu, Lihua Wu, Xiaoyu Qu, Linli Dai, Yiquan Ye, Shanshan Xu, Chao Wu, and Yu Chen. 2023. "Correlation between Changes in Soil Properties and Microbial Diversity Driven by Different Management in Artificial Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.) Plantations" Forests 14, no. 5: 877. https://doi.org/10.3390/f14050877