

Influence of Natural and Man-Made Forests on Community Assembly and Species, Functional, and Soil Microbial Diversity

Abstract
:1. Introduction
2. Materials and Methods
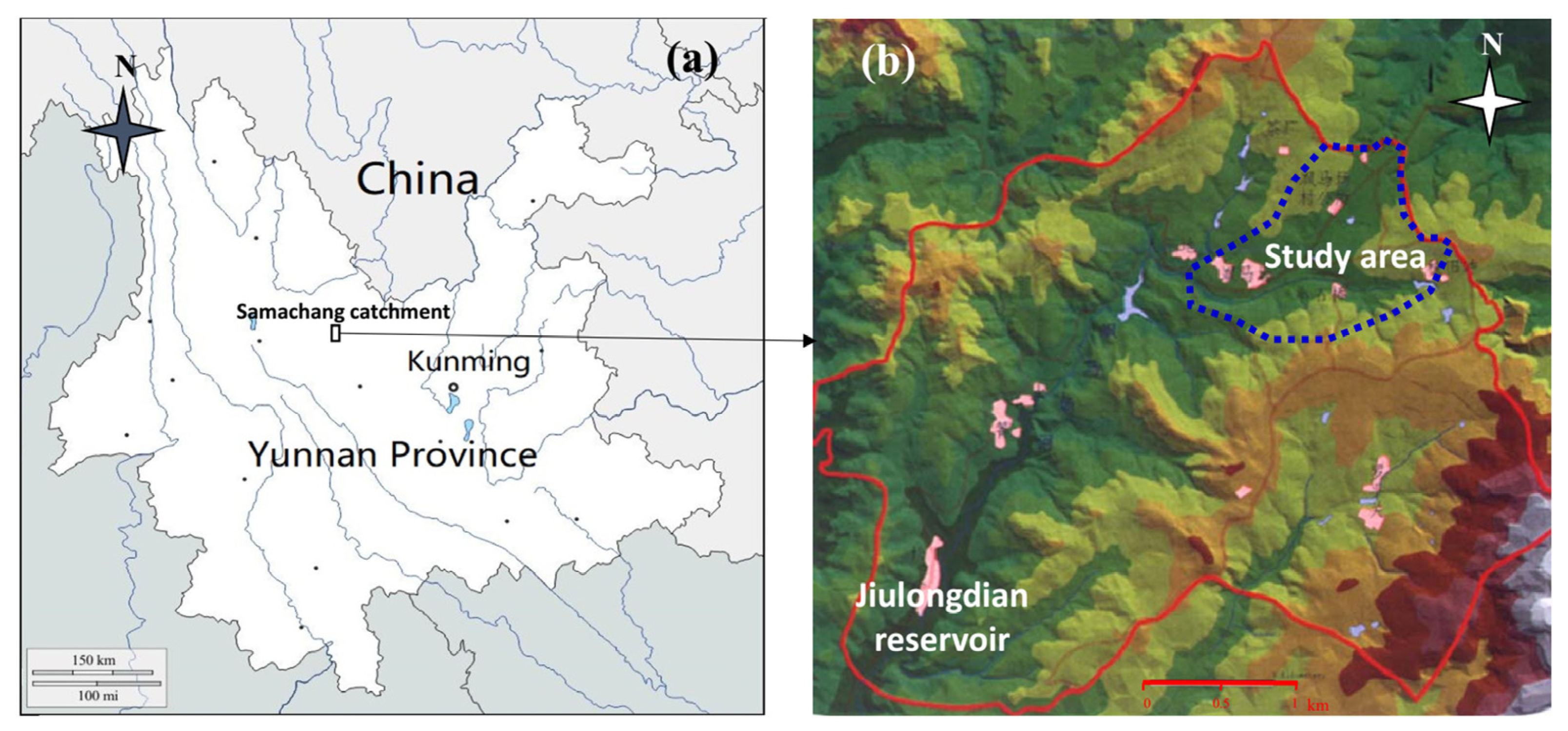
2.1. Study Site
2.2. Data Sampling Procedure
2.2.1. Plant Community Investigation and Species Diversity
2.2.2. Community Functional Characteristics
2.2.3. Soil Microbial Community
2.3. Data Analysis
2.3.1. Statistical Analyses of Co-Occurrence and Trait Dispersion
2.3.2. Analyses of the Relationship between Species Diversity and Functional Diversity
2.3.3. Relationships between Soil Microbial Diversity and Species/Functional Diversity
3. Results
3.1. Species Co-Occurrence and Trait Dispersion
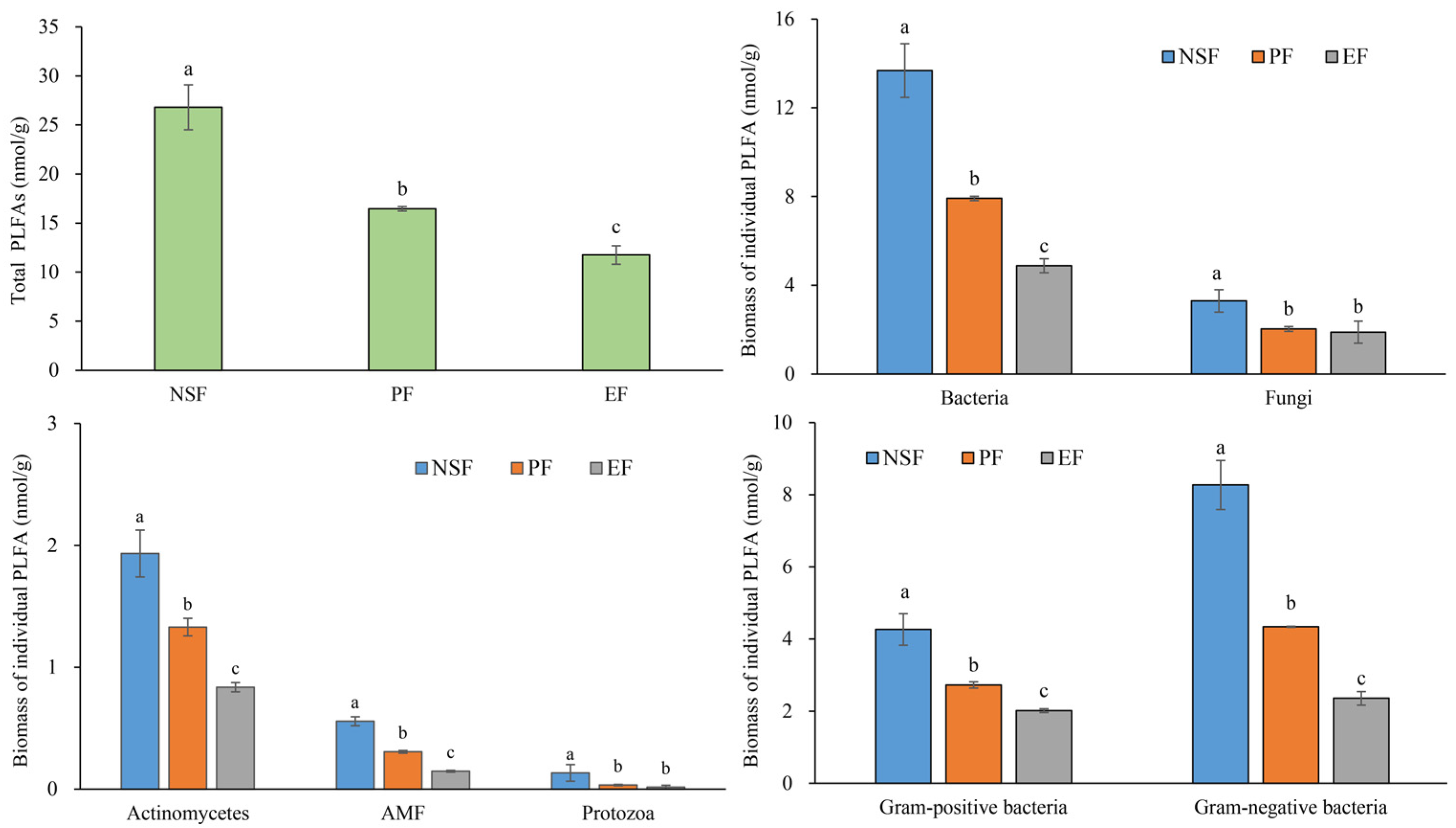
3.2. Species Richness, Functional Diversity, and Soil Microbial Diversity
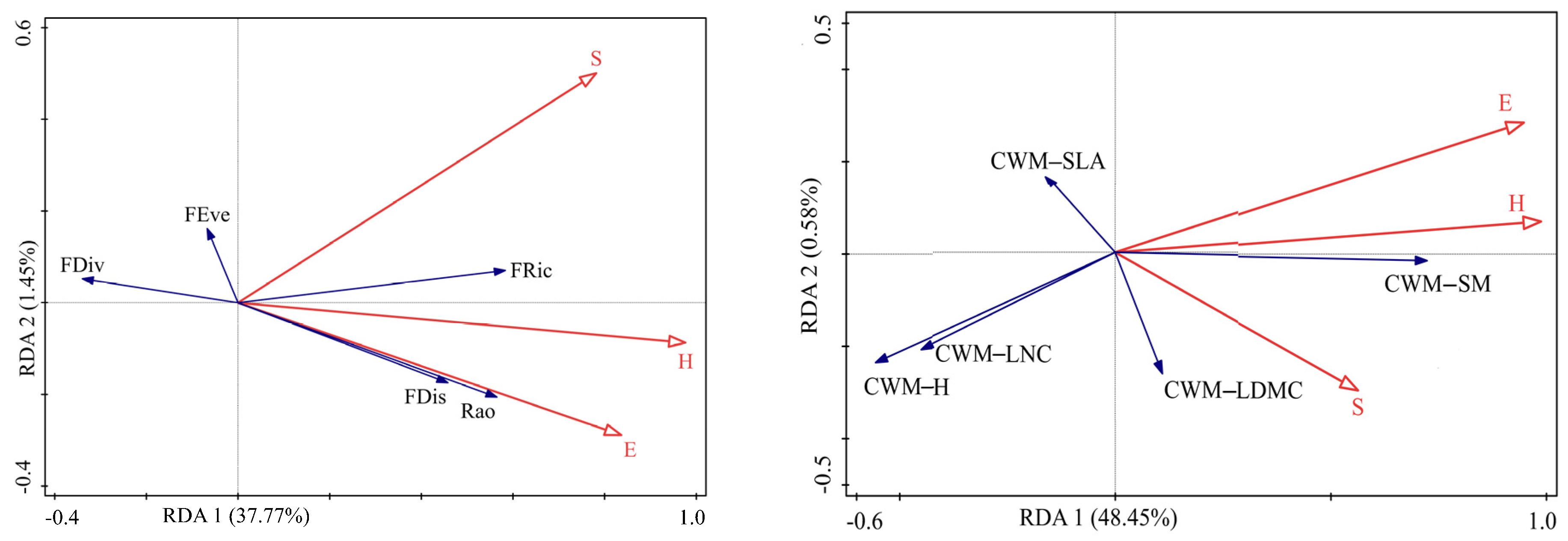
3.3. Relationships among Species Diversity, Functional Diversity, CWMs, and Soil Microbial Diversity
4. Discussion
4.1. Community Assembly at the Species and Trait Levels
4.2. Difference in the Diversity Index between Natural and Man-Made Forests
4.3. Relationship between Species Diversity and Functional Diversity
4.4. Relative Contributions of Species and Functional Diversity to Soil Microbial Diversity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Doran, J.W.; Zeiss, M.R. Soil Health and Sustainability: Managing the Biotic Component of Soil Quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global Threats to Human Water Security and River Biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef]
- Cerdà, A.; Lucas-Borja, M.E.; Franch-Pardo, I.; Úbeda, X.; Novara, A.; López-Vicente, M.; Popović, Z.; Pulido, M. The Role of Plant Species on Runoff and Soil Erosion in a Mediterranean Shrubland. Sci. Total Environ. 2021, 799, 149218. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Duan, C.; Chadwick, D.R.; Jones, D.L. Response of Soil Phosphorus Fractions and Fluxes to Different Vegetation Restoration Types in a Subtropical Mountain Ecosystem. CATENA 2020, 193, 104663. [Google Scholar] [CrossRef]
- Tang, C.Q.; Hou, X.; Gao, K.; Xia, T.; Duan, C.; Fu, D. Man-Made versus Natural Forests in Mid-Yunnan, Southwestern China. Mt. Res. Dev. 2007, 27, 242–249. [Google Scholar] [CrossRef]
- Cao, S.; Chen, L.; Yu, X. Impact of China’s Grain for Green Project on the Landscape of Vulnerable Arid and Semi-arid Agricultural Regions: A Case Study in Northern Shaanxi Province. J. Appl. Ecol. 2009, 46, 536–543. [Google Scholar] [CrossRef]
- Geldenhuys, C.J. Native Forest Regeneration in Pine and Eucalypt Plantations in Northern Province, South Africa. For. Ecol. Manag. 1997, 99, 101–115. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Huang, N.; Duan, C. Effects of the Invasive Herb Ageratina Adenophora on Understory Plant Communities and Tree Seedling Growth in Pinus yunnanensis Forests in Yunnan, China. J. For. Res. 2018, 23, 112–119. [Google Scholar] [CrossRef]
- Widyati, E.; Nuroniah, H.S.; Tata, H.L.; Mindawati, N.; Lisnawati, Y.; Abdulah, L.; Lelana, N.E.; Octavia, D.; Prameswari, D.; Rachmat, H.H. Soil Degradation Due to Conversion from Natural to Plantation Forests in Indonesia. Forests 2022, 13, 1913. [Google Scholar] [CrossRef]
- Dupré, C.; Ehrlén, J. Habitat Configuration, Species Traits and Plant Distributions. J. Ecol. 2002, 90, 796–805. [Google Scholar] [CrossRef]
- Weiher, E.; Clarke, G.D.P.; Keddy, P.A. Community Assembly Rules, Morphological Dispersion, and the Coexistence of Plant Species. Oikos 1998, 81, 309–322. [Google Scholar] [CrossRef]
- Zobel, M. The Relative of Species Pools in Determining Plant Species Richness: An Alternative Explanation of Species Coexistence? Trends Ecol. Evol. 1997, 12, 266–269. [Google Scholar] [CrossRef]
- MacArthur, R.; Levins, R. The limiting Similarity Convergence and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Schamp, B.S.; Chau, J.; Aarssen, L.W. Dispersion of Traits Related to Competitive Ability in an Old-field Plant Community. J. Ecol. 2008, 96, 204–212. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing Functional Diversity in the Field–Methodology Matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant Functional Markers Capture Ecosystem Properties during Secondary Succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Strahan, R.T.; Sánchez Meador, A.J.; Huffman, D.W.; Laughlin, D.C. Shifts in Community-level Traits and Functional Diversity in a Mixed Conifer Forest: A Legacy of Land-use Change. J. Appl. Ecol. 2016, 53, 1755–1765. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Z.; Castro, D.M.P.; Tan, X.; Jiang, X.; Meng, X.; Ge, Y.; Xie, Z. Effects of Different Types of Land-Use on Taxonomic and Functional Diversity of Benthic Macroinvertebrates in a Subtropical River Network. Environ. Sci. Pollut. Res. 2021, 28, 44339–44353. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Strahan, R.T.; Moore, M.M.; Fulé, P.Z.; Huffman, D.W.; Covington, W.W. The Hierarchy of Predictability in Ecological Restoration: Are Vegetation Structure and Functional Diversity More Predictable than Community Composition? J. Appl. Ecol. 2017, 54, 1058–1069. [Google Scholar] [CrossRef]
- Khlifa, R.; Paquette, A.; Messier, C.; Reich, P.B.; Munson, A.D. Do Temperate Tree Species Diversity and Identity Influence Soil Microbial Community Function and Composition? Ecol. Evol. 2017, 7, 7965–7974. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Bauhus, J.; Paré, D.; Côté, L. Effects of Tree Species, Stand Age and Soil Type on Soil Microbial Biomass and Its Activity in a Southern Boreal Forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- de Vries, F.T.; Manning, P.; Tallowin, J.R.B.; Mortimer, S.R.; Pilgrim, E.S.; Harrison, K.A.; Hobbs, P.J.; Quirk, H.; Shipley, B.; Cornelissen, J.H.C.; et al. Abiotic Drivers and Plant Traits Explain Landscape-Scale Patterns in Soil Microbial Communities. Ecol. Lett. 2012, 15, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.; Eichenberg, D.; Bruelheide, H.; Kröber, W.; Kühn, P.; Li, Y.; von Oheimb, G.; Purschke, O.; Scholten, T.; Buscot, F.; et al. Soil and Tree Species Traits Both Shape Soil Microbial Communities during Early Growth of Chinese Subtropical Forests. Soil Biol. Biochem. 2016, 96, 180–190. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Duan, C.; Smith, A.R.; Jones, D.L. Traits of Dominant Species and Soil Properties Co-Regulate Soil Microbial Communities across Land Restoration Types in a Subtropical Plateau Region of Southwest China. Ecol. Eng. 2020, 153, 105897. [Google Scholar] [CrossRef]
- Fu, D.; Duan, C.; Hou, X.; Xia, T.; Gao, K. Patterns and Relationships of Plant Traits, Community Structural Attributes, and Eco-Hydrological Functions during a Subtropical Secondary Succession in Central Yunnan (Southwest China). Arch. Biol. Sci. 2009, 61, 741–749. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Duan, C.; Guan, Q.; Huang, N. Changes in Functional Structure Characteristics Mediate Ecosystem Functions during Human-Induced Land-Cover Alteration: A Case Study in Southwest China. J. Soil Water Conserv. 2018, 73, 461–468. [Google Scholar] [CrossRef]
- Hou, X.; Duan, C.; Tang, C.Q.; Fu, D. Nutrient Relocation, Hydrological Functions, and Soil Chemistry in Plantations as Compared to Natural Forests in Central Yunnan, China. Ecol. Res. 2010, 25, 139–148. [Google Scholar] [CrossRef]
- Casanoves, F.; Pla, L.; Di Rienzo, J.A.; Díaz, S. FDiversity: A Software Package for the Integrated Analysis of Functional Diversity. Methods Ecol. Evol. 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A Handbook of Protocols for Standardised and Easy Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Vile, D.; Shipley, B.; Garnier, E. A Structural Equation Model to Integrate Changes in Functional Strategies during Old-Field Succession. Ecology 2006, 87, 504–517. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Phospholipid Fatty Acid Composition, Biomass, and Activity of Microbial Communities from Two Soil Types Experimentally Exposed to Different Heavy Metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J. EcoSim: Null Models Software for Ecology. Version 7. Acquired Intelligence Inc. and Kesey-Bear. 2009. Available online: http//garyentsminger.com/ecosim.htm (accessed on 31 January 2015).
- Swenson, T.W.E.; Weiher, E. Nellie: Null Models for the PC; University of Wisconsin Eau-Claire: Eau Claire, WI, USA, 2000. [Google Scholar]
- Stone, L.; Roberts, A. The Checkerboard Score and Species Distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef]
- Schluter, D. A Variance Test for Detecting Species Associations, with Some Example Applications. Ecology 1984, 65, 998–1005. [Google Scholar] [CrossRef]
- Gotelli, N.J. Null Model Analysis of Species Co-occurrence Patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly Rules, Null Models, and Trait Dispersion: New Questions from Old Patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef]
- Mayfield, M.M.; Boni, M.F.; Daily, G.C.; Ackerly, D. Species and Functional Diversity of Native and Human-dominated Plant Communities. Ecology 2005, 86, 2365–2372. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Bar-Massada, A.; Yang, Q.; Shen, G.; Wang, X. Tree Species Co-occurrence Patterns Change across Grains: Insights from a Subtropical Forest. Ecosphere 2018, 9, e02213. [Google Scholar] [CrossRef]
- Tian, K.; Chen, L.; Mi, X.; Ma, K.P.; Chen, J.H. The Effect of Habitat Filtering on Tree Seedling Distribution in a Subtropical Evergreen Broadleaf Forest in China. Chin. Sci. Bull. 2013, 58, 3561–3569. [Google Scholar]
- Zhang, B.; Lu, X.; Jiang, J.; DeAngelis, D.L.; Fu, Z.; Zhang, J. Similarity of Plant Functional Traits and Aggregation Pattern in a Subtropical Forest. Ecol. Evol. 2017, 7, 4086–4098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, Z.; Song, B.; Li, B.; Wang, X.; Ye, J. Fine-Scale Species Co-Occurrence Patterns in an Old-Growth Temperate Forest. For. Ecol. Manag. 2009, 257, 2115–2120. [Google Scholar] [CrossRef]
- Tang, C.Q.; Zhao, M.-H.; Li, X.-S.; Ohsawa, M.; Ou, X.-K. Secondary Succession of Plant Communities in a Subtropical Mountainous Region of SW China. Ecol. Res. 2010, 25, 149–161. [Google Scholar] [CrossRef]
- Ibsen, P.C.; Borowy, D.; Rochford, M.; Swan, C.M.; Jenerette, G.D. Influence of Climate and Management on Patterns of Taxonomic and Functional Diversity of Recreational Park Vegetation. Front. Ecol. Evol. 2020, 8, 501502. [Google Scholar] [CrossRef]
- Dai, X.; Chen, C.; Li, Z.; Wang, X. Taxonomic, Phylogenetic, and Functional Diversity of Ferns at Three Differently Disturbed Sites in Longnan County, China. Diversity 2020, 12, 135. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Chen, Y.; Zhao, J.; Wan, S.; Lin, Y.; Fu, S. Effects of Eucalyptus Litter and Roots on the Establishment of Native Tree Species in Eucalyptus Plantations in South China. For. Ecol. Manag. 2016, 375, 76–83. [Google Scholar] [CrossRef]
- Fu, D.; Wu, X.; Qiu, Q.; Duan, C.; Jones, D.L. Seasonal Variations in Soil Microbial Communities under Different Land Restoration Types in a Subtropical Mountains Region, Southwest China. Appl. Soil Ecol. 2020, 153, 103634. [Google Scholar] [CrossRef]
- Yannikos, N.; Leinweber, P.; Helgason, B.L.; Baum, C.; Walley, F.L.; Rees, K.C.J. Van Impact of Populus Trees on the Composition of Organic Matter and the Soil Microbial Community in Orthic Gray Luvisols in Saskatchewan (Canada). Soil Biol. Biochem. 2014, 70, 5–11. [Google Scholar] [CrossRef]
- Tedersoo, L.; Brundrett, M.C. Evolution of Ectomycorrhizal Symbiosis in Plants. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 407–467. ISBN 978-3-319-56363-3. [Google Scholar]
- Strickland, M.S.; Rousk, J. Considering Fungal:Bacterial Dominance in Soils–Methods, Controls, and Ecosystem Implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Fukami, T.; Martijn Bezemer, T.; Mortimer, S.R.; van der Putten, W.H. Species Divergence and Trait Convergence in Experimental Plant Community Assembly. Ecol. Lett. 2005, 8, 1283–1290. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Null Model | Observed Index | Mean of Simulated Indices | SES | p |
|---|---|---|---|---|
| C-score | ||||
| Pinus yunnanensis forest | 4.46 | 5.1 | −1.83 | 0.045 |
| Eucalyptus smithii forest | 5.07 | 5.25 | −0.51 | 0.722 |
| Natural secondary forest | 5.57 | 7.33 | −3.75 | 0.000 |
| V-ratio | ||||
| Pinus yunnanensis forest | 1.82 | 0.99 | 2.30 | 0.026 |
| Eucalyptus smithii forest | 1.22 | 1.00 | 0.59 | 0.272 |
| Natural secondary forest | 2.55 | 1.00 | 4.30 | 0.000 |
| Diversity Indices | NSF | PF | EF |
|---|---|---|---|
| Species diversity indices | |||
| Species richness (S) | 10.50 ± 2.27 a | 9.50 ± 2.46 a | 9.11 ± 2.32 a |
| Shannon-Wiener index (H) | 1.44 ± 0.27 a | 1.15 ± 0.29 b | 0.95 ± 0.20 b |
| Species evenness (E) | 0.62 ± 0.09 a | 0.51 ± 0.12 b | 0.43 ± 0.04 b |
| Functional diversity indices | |||
| Functional richness (FRic) § | 17.41 ± 1.29 a | 11.42 ± 2.68 b | 13.61 ± 3.59 b |
| Functional evenness (FEve) | 0.45 ± 0.11 a | 0.50 ± 0.11 a | 0.51 ± 0.12 a |
| Functional divergence (FDiv) | 0.44 ± 0.18 b | 0.47 ± 0.10 b | 0.79 ± 0.25 a |
| Functional dispersion (FDis) § | 5.97 ± 0.30 a | 3.97 ± 0.54 b | 4.50 ± 0.55 b |
| Rao entropy § | 12.52 ± 0.60 a | 8.82 ± 1.26 b | 9.98 ± 1.64 b |
| Community-weighted means (CWM) | |||
| CWM-LDMC | 410.4 ± 14.3 a | 329.2 ± 9.1 b | 419.4 ± 6.57 a |
| CWM-SLA | 7.96 ± 1.01 b | 16.10 ± 1.89 a | 7.34 ± 1.15 b |
| CWM-H | 6.74 ± 1.47 b | 5.51 ± 0.58 c | 18.28 ± 1.06 a |
| CWM-LNC | 9.14 ± 0.19 b | 8.78 ± 0.55 b | 13.64 ± 0.69 a |
| CWM-SM | 632.1 ± 86.9 a | 40.7 ± 27.2 b | 42.1 ± 61.5 b |
| Soil microbial composition diversity | |||
| Shannon-Wiener index (H) | 3.81 ± 0.75 a | 3.81 ± 0.716 a | 3.78 ± 0.71 a |
| Species richness (S) | 48.50 ± 1.71 a | 43.67 ± 1.70 b | 42.67 ± 1.80 b |
| Pielou evenness (J) | 0.98 ± 0.18 a | 1.01 ± 0.19 a | 1.01 ± 0.19 a |
| Traits | Forest Types | Model | Observed CWM § | Predicted CWM § | |
|---|---|---|---|---|---|
| a | R2 | ||||
| LDMC | PF | 0.018 | 0.148 | −0.57 | −0.55 c ± 0.044 |
| EF | 0.011 | 0.203 | −0.39 | −0.44 b ± 0.027 | |
| NSF | 0.025 * | 0.375 | −0.25 | −0.28 a ± 0.053 | |
| SLA | PF | 0.006 | 0.004 | −0.78 | −0.49 b ± 0.014 |
| EF | 0.030 ** | 0.819 | −0.40 | −0.45 b ± 0.068 | |
| NSF | 0.048 | 0.294 | −0.23 | −0.29 a ± 0.103 | |
| H | PF | 0.015 | 0.112 | −0.49 | −0.42 b ± 0.037 |
| EF | 0.029 ** | 0.906 | −0.36 | −0.39 b ± 0.063 | |
| NSF | 0.022 * | 0.401 | −0.31 | −0.32 a ± 0.046 | |
| LNC | PF | 0.005 | 0.018 | −0.60 | −0.53 ns ± 0.013 |
| EF | 0.087 ** | 0.908 | −0.45 | −0.60 ns ± 0.201 | |
| NSF | 0.037 * | 0.365 | −0.59 | −0.59 ns ± 0.079 | |
| SM | PF | 0.008 | 0.031 | −0.47 | −0.33 b ± 0.020 |
| EF | 0.031 ** | 0.903 | −0.36 | −0.39 c ± 0.070 | |
| NSF | 0.025 * | 0.401 | −0.18 | −0.19 a ± 0.053 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Ma, X.; Hu, L.; Shen, C.; Fu, D. Influence of Natural and Man-Made Forests on Community Assembly and Species, Functional, and Soil Microbial Diversity. Forests 2023, 14, 888. https://doi.org/10.3390/f14050888
Wu X, Ma X, Hu L, Shen C, Fu D. Influence of Natural and Man-Made Forests on Community Assembly and Species, Functional, and Soil Microbial Diversity. Forests. 2023; 14(5):888. https://doi.org/10.3390/f14050888
Chicago/Turabian StyleWu, Xiaoni, Xudong Ma, Lianyu Hu, Chunjie Shen, and Denggao Fu. 2023. "Influence of Natural and Man-Made Forests on Community Assembly and Species, Functional, and Soil Microbial Diversity" Forests 14, no. 5: 888. https://doi.org/10.3390/f14050888