
Genome-Wide Analysis, Identification, and Characterization of the PFK Gene Family Members of Populus deltoides
by
 , , , and
, , , and
Tae-Lim Kim
1,†,
Michael Immanuel Jesse Denison
2,†,
Hyemin Lim
1,*,
Hoyong Chung
2 and
Changyoung Oh
1 1
Department of Forest Bioresources, National Institute of Forest Science, Suwon 16631, Republic of Korea
2
3BIGS Company Limited, Hwaseong 18469, Republic of Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2023, 14(6), 1104; https://doi.org/10.3390/f14061104
Submission received: 11 May 2023
/
Revised: 24 May 2023
/
Accepted: 24 May 2023
/
Published: 26 May 2023
(This article belongs to the Special Issue Genome-Wide Identification and Expression Analysis for the Genetic Improvement of Forest Plants)
Abstract
:Plants produce two phosphorylated enzymes: pyrophosphate-dependent fructose-6-phosphate phosphotransferase (PFP) and ATP-dependent phosphofructokinase (PFK). However, the characterization of phosphofructokinase (PFK) family members and their functions remains unexplored in Eastern Cottonwood (Populus deltoides). This study comprehensively analyzed 17 phosphofructokinase genes in the P. deltoides genome, including their gene structure, conserved domains, evolutionary relationships, and expression patterns. Genome-wide analysis of the poplar genome identified 11 genes belonging to PdPFK and 6 genes belonging to PdPFP. Phylogenetic, gene structure, and motif analyses revealed two main categories of PFK genes: PFK with eleven genes and PFP with six genes. Quantitative PCR (qPCR) analysis demonstrated that all 17 phosphofructokinase genes were induced in various tissues and developmental stages of P. deltoides. PFPs were strongly expressed throughout P. deltoides’ growth and development, notably in the roots, according to the transcript analysis. This genome-wide analysis provides insights into PFK and PFP genes and establishes a foundation for future investigations into the role of genes in P. deltoids.
1. Introduction
Glycolysis, which involves the conversion of glucose to pyruvic acid, is a crucial respiratory pathway in plants. Glycolysis provides ATP, reducing agents, and precursors for plant growth and development and helps cells adapt to cold, drought, and anoxia [1,2,3,4]. The enzymes PFP and PFK, which phosphorylate fructose-6-phosphate, are found in plants. Fructose-6-phosphate phosphotransferase (PFP, EC 1.7.1.90) catalyzes the phosphorylation of fructose-6-phosphate (F-6-P) into fructose-1,6-bisphosphate (F-1,6-BP) in the glycolytic pathway. It also facilitates the dephosphorylation of F-1,6-BP back to F-6-P in the gluconeogenic direction during primary carbohydrate metabolism in the cytosol [5]. In contrast with ATP-dependent phosphofructokinase (PFK, EC 2.7.1.11), which catalyzes the irreversible phosphorylation of F-6-P to F-1,6-BP, PFP is an enzyme that catalyzes the reversible phosphorylation of F-6-P to F-1,6-BP [4]. In contrast with plants that possess both PFP and PFK [6], bacteria and protozoans have only one type, while yeasts and animals exclusively rely on PFK [1,2,4].
Phosphofructokinase gene families have been extensively studied, and several genes have been identified in various plants. For instance, Arabidopsis has 11 PFK genes [7], Saccharum has 10 genes [8], Oryza sativa has 15 genes [9], and Pyrus bretschneideri has 14 genes [10]. PFK plays a central role in plant growth and development, serving multiple functions. The PFK family comprises more than ten members. Interestingly, the activity of PFP is comparable to or sometimes even surpasses that of PFK, and it is present in diverse plant tissues [11]. The activity and composition of PFP enzymes, including subunits, are influenced by environmental factors, developmental stages, tissue specificity, species variation, and growth conditions [12,13,14]. In most plant tissues, PFP exists as a heterotetramer composed of two catalytic subunits and two regulatory subunits, typically β2 and α2β2 [15,16,17]. The β subunit of PFP acts as a catalyst, while the α subunit controls the catalytic activity of fructose-2,6-bisphosphate (Fru-2,6-BP) [11,18,19]. However, plants have been found to exhibit copy number differences in the genes that encode these subunits [20]. It has been reported that under a variety of circumstances, including anoxia, wounds, phosphate shortage, excessive salt, and dehydration, PFP is engaged in stress responses [2,4,21,22]. However, the specific function and relevance of PFP remain unclear.
Poplar is a representative model tree species that grows rapidly, exhibits strong adaptability, and is widely used for greening, afforestation, and production because of its excellent properties. P. deltoides, which is native to the southeastern United States and southern Canada, is a member of the section Aigeiros [23]. In South Korea, the Korea National Commission on Poplars and Other Fast-Growing Trees (KCFGT) reported the establishment of poplar plantations in the reclaimed tidelands of Saemangeum for restoration and timber production between 2013 and 2018. This involved a total of 156.5 hectares and included the planting of P. euramericana, P. deltoides, and P. alba × P. tremula var. glandulosa (source: https://www.kcfgt.org/, accessed on 11 May 2023). Currently, P. deltoides is widely distributed in the forests of Korea. Genomic information on P. deltoides has been previously published [24]. As a species of significant ecological and economic value, it is characterized by rapid growth, resistance to disease, a moderate genome size, and easy experimental manipulation [24].
In this study, we carried out a systematic and thorough analysis of the poplar PFP and PFK families, including chromosomal location analysis, subcellular localization, family member identification and categorization, and phylogenetic analysis, using the published genome as a reference. These investigations serve as a solid foundation for understanding the functions of PFP and PFK in poplar and exploring the underlying regulatory mechanisms.
2. Materials and Methods
2.1. Tree Materials
The experimental material used in this study was the ‘Ay48’ clone of 12-year-old P. deltoides. These trees were planted at the National Institute of Forest Science in Korea, with the coordinates 37°15′04″ N, 136°57′59″ E. Plants aged 1 year were propagated from 15 cm long dormant cuttings and grown in pots filled with topsoil in a greenhouse. The greenhouse maintained a temperature range of 18–25 °C. Fresh tissue samples were collected from the poplar trees, immediately frozen in liquid nitrogen, and stored at −80 °C until further extraction of RNA and protein.
2.2. Identification of Phosphofructokinase Family Genes in P. deltoides
The phosphofructokinase gene family was identified based on the presence of a PFK domain. To identify the members of the phosphofructokinase gene family in Populus genomes, whole genome and proteome data on P. deltoides (var. WV94) genome assembly v2.1 were downloaded from Phytozome version V13 (https://phytozome-next.jgi.doe.gov/info/PdeltoidesWV94_v2_1, accessed on 24 April 2023). The hidden Markov model (HMM) profile of the conserved functional domain of phosphofructokinase (PFK) (PF00365) was obtained from InterPro 93.0 (https://www.ebi.ac.uk/interpro/, accessed on 24 April 2023). The HMM profile was then used to screen Populus proteomes using the hmmsearch software in the HMMER package v3.0 to identify potential members of the PFK gene family. After removing redundant and incomplete sequences, the conserved domain architectures of the recovered sequences were confirmed using the Pfam database in the Conserved Domain Interface (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi, accessed on 24 April 2023). Sequences exhibiting the typical functional domain of a phosphofructokinase were considered potential members of the Populus phosphofructokinase family.
2.3. Chromosomal Location and Collinearity Analyses of PdPFK Genes
The PFK-domain-containing PdPFPs and PdPFKs were mapped onto the chromosomes based on the poplar genome information from the Phytozome database and visualized using TB tools (ToolBox for Biologists) (Version 1.121) [25]. In addition, the genome data and gene annotation files from Phytozome version V13 (https://phytozome-next.jgi.doe.gov/info/PdeltoidesWV94_v2_1, accessed on 24 April 2023) were downloaded to investigate the syntenic relationship between PFK genes on poplar chromosomes. MCScanX (Multiple Collinearity Scan toolkit X version) [26] was used to detect gene synteny and collinearity using standard Pa parameters. A circular plot was constructed using TBTools (Version 1.121).
2.4. Conserved Motifs and Gene Structure Analysis
Gene features, such as the composition and positions of exons, introns, and conserved elements of PdPFPs and PdPFKs, were visualized by aligning coding sequence regions (CDSs) to genomic DNA sequences in the Gene Structure Display Server (GSDS) (http://gsds.gao-lab.org/Gsd.sabout.Php, accessed on 18 April 2023). Motifs were predicted using the online tool Multiple Expectation Maximization for Motif Elicitation (MEME) (http://memesuite.org/tools/meme, accessed on 18 April 2023). Based on the protein sequences of all PFK members, the online tool ExPASy (https://www.expasy.org/tools/, accessed on 18 April 2023) was used to determine the molecular weight (MW) and theoretical isoelectric point (pI).
2.5. Phylogenetic Analysis and Classification of the Phosphofructokinase Genes
Complete amino acid sequences of PFK from Arabidopsis thaliana (TAIR10.1), Solanum tuberosum (SolTub_3.0), Ricinus communis (ASM1957865v1), and Oryza sativa (IRGSP-1.0) were retrieved from the NCBI database. ClustalW (https://www.genome.jp/tools-bin/clustalw, accessed on 13 April 2023) was used to perform multiple sequence alignments between the PFK proteins of all five species, including P. deltoides. Phylogenetic trees were constructed based on protein alignment using iTOL software. The final phylogenetic tree was obtained using iTOL (https://itol.embl.de/, accessed on 13 April 2023) [27], and the output was established for visualization using Adobe Photoshop (Version 2020).
2.6. RNA Extraction and Quantitative PCR (qPCR)
To extract total RNA from poplar tissues, the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany) was utilized. Three biological replicates were used for RNA extraction. Subsequently, single-stranded cDNA was synthesized from the RNA samples using the cDNA EcoDryTM Premix (TaKaRa, Shiga, Japan). IQtm SYBR Green Supermix and a CFX96 Touch Real-Time PCR Detection System from Bio-Rad, Hercules, CA, USA were used to conduct qPCR. The qPCR reaction consisted of an initial denaturation step at 95 °C for 3 min, followed by 36 cycles of amplification at 95 °C for 15 s and 60 °C for 60 s. Technical replicates were performed three times using samples derived from three independent biological replicates. The relative transcript abundance was analyzed using the 2−ΔΔCt method [28]. UBQ7 and ACTIN were used as reference genes to normalize the qPCR results [29]. In Supplementary Table S1, the gene-specific primers that were employed are provided.
2.7. Measurement of Phosphofructokinase Content
To determine the phosphofructokinase (PFK) content in homogenized leaf, stem, and root samples, a PFK ELISA kit (MyBioSource, San Diego, CA, USA) was used. We followed the manufacturer’s directions for every procedure. After the completion of the substrate reaction, absorbance readings were taken at 450 nm using an automated plate reader (SpectraMax M2, Molecular Devices, San Jose, CA, USA).
2.8. Statistical Analysis
One-way ANOVA was used for the analyses, and Tukey’s honest significant difference test (HSD) was used for multiple comparisons. For statistical significance, p values of less than 0.05 were considered significant. The results are presented as means with standard deviation (SD).
3. Results
3.1. Identification and Physicochemical Properties of Phosphofructokinase Gene Family in P. deltoides
A total of 17 PFK gene family members were downloaded from the InterPro database using PFK-domain-hidden Markov models (PF00365) (as of 28 March 2023) and used as queries to search in the local P. deltoides database using the hmmer program [30]. The identified PFK proteins were checked for the presence of redundant proteins using the NCBI Conserved Database (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb /bwrpsb.cgi, accessed on 18 April 2023). Seventeen PFK family members were found to have conserved domains in all protein sequences, leading to the final identification of PFKs (Table 1). The lengths of 17 PFK proteins, which is a crucial physicochemical characteristic, ranged from 86 amino acids (Podel.01G082600.1.p) to 617 amino acids (Podel.05G274300.1.p), with a mean length of 435 amino acids. The proteins were 9.74 (Podel.01G082600.1.p) to 67.24 (Podel.05G274300.1.p) KDa in relative molecular weight. The isoelectric points in theory ranged from 5.71 to 9.07. More than seven isoelectric points were present in five PFK genes. These proteins were alkaline, whereas the other proteins were acidic.
3.2. Prediction of the Subcellular Localization of Phosphofructokinase Family Members of P. deltoides
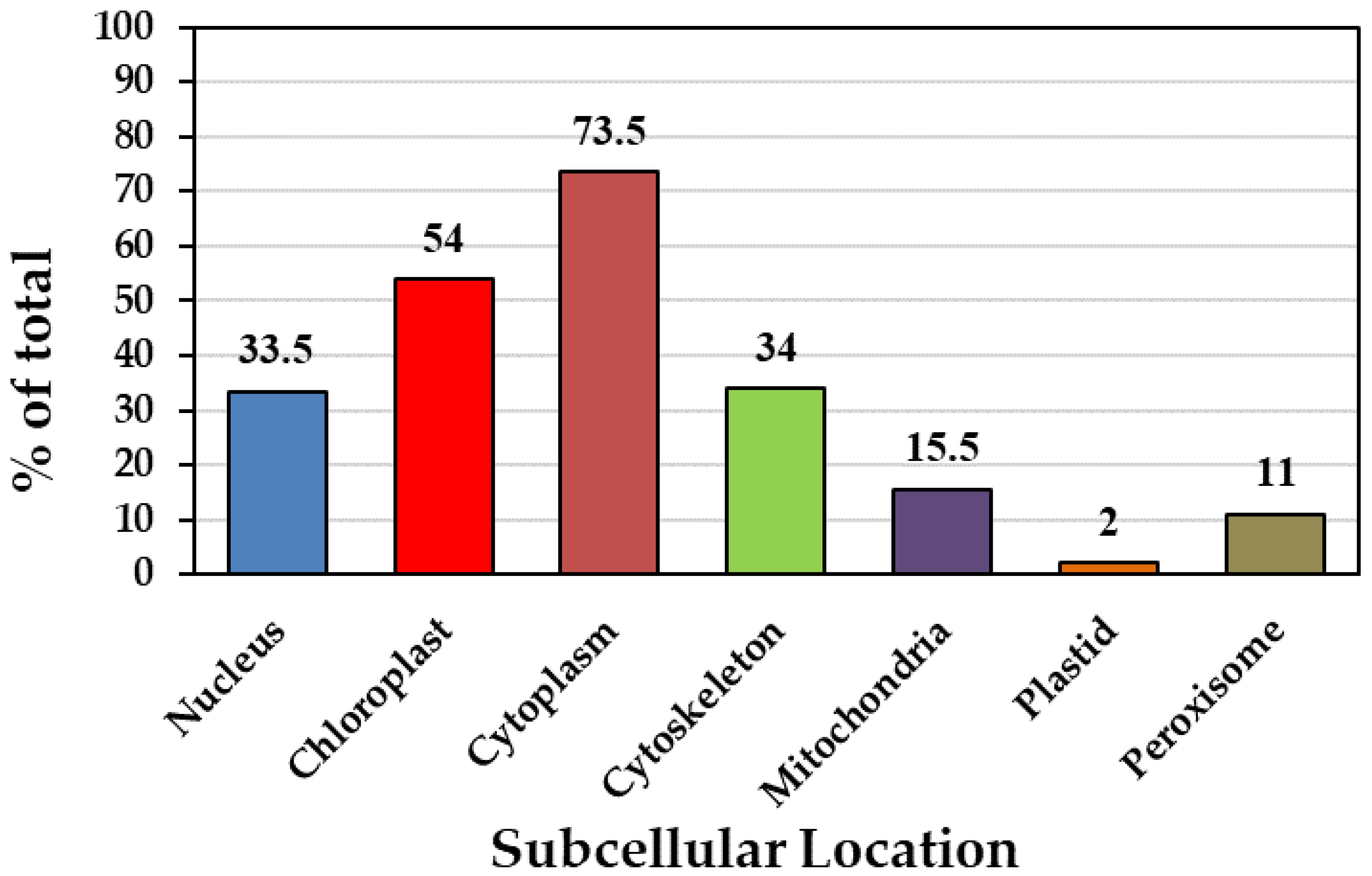
In silico analysis revealed that Podel.01G054800.1.p is localized in the nucleus, with four proteins in the chloroplasts, seven in the cytoplasm, three in the cytoskeleton, one (Podel.T151900.1.p) in the mitochondria, and one (Podel.T151900.1.p) in peroxisomes. According to the projected subcellular distribution, the cytoplasm hosted 73% of the PFK proteins, chloroplasts hosted 54%, the cytoskeleton hosted 34%, and the nucleus hosted 33.5% (Figure 1, Table 1).
3.3. Chromosomal Location and Collinearity Analysis of PdPFP and PdPFK Gene Families
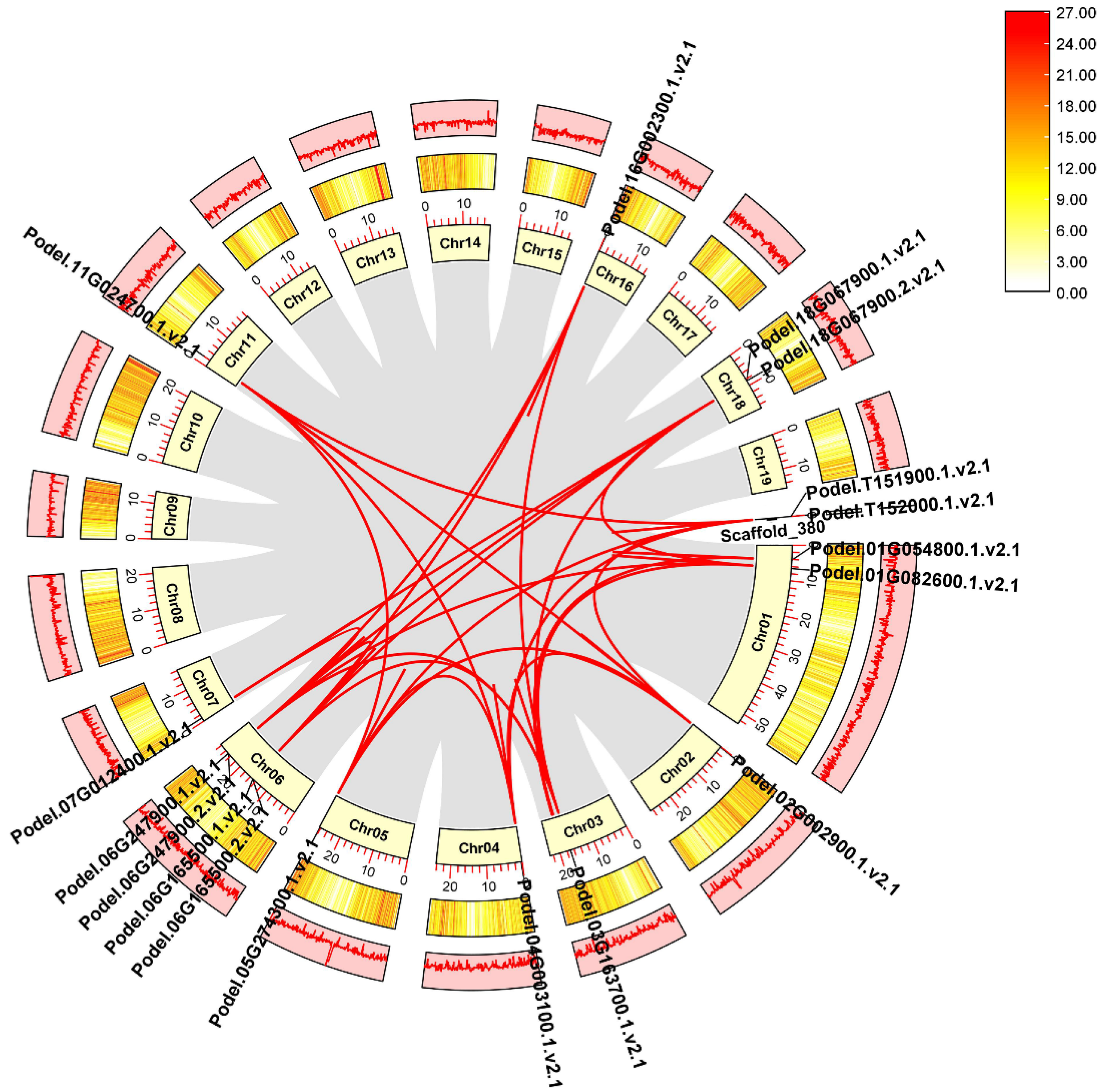
We mapped phosphofructokinase proteins on P. deltoides chromosomes and found that PdPFK genes were mapped on chromosomes Chr01, Chr03, Chr04, Chr05, Chr06, Chr07, and Chr16, whereas six PdPFPs were located on Chr02, Chr11, Chr18, and scaffold 380 (Figure 2). Most of the PdPFKs (four genes) were mapped on chromosome Chr06 of P. deltoides. Chromosome Chr18 and the unplaced Scaffold_380 harbored several PdPFPs (two each). Chr01, Chr03-Chr07, and Chr16 were occupied by PdPFKs. In general, chromosome length did not positively correlate with gene distribution (Figure 2).
Gene duplication events in P. deltoides chromosomes lead to the expansion of gene families. These events were identified using the homology-based BLAST approach in the MCScanX toolkit. At the genome level of P. deltoides, 21,626 gene pairs and 1922 groupings of tandem duplication genes were discovered. The identified 17 PdPFK genes were unevenly distributed across the 19 chromosomes of P. deltoides (Figure 3). Using TBtools, the synteny gene pairs and tandem duplication gene pairs of the PdPFK gene family were chosen and visualized. (Figure 3). Using the MCscanX and Advanced Circos software from TBTools, we conducted synteny analysis to comprehend the duplication events of all PdPFK genes. On 10 of the 19 chromosomes of P. deltoides, there were a total of 85 pairs of duplicated genes that were scattered irregularly. Additionally, genes scattered on chromosomes 1 and 2 were syntenic with genes on chromosomes 2, 4, 5, and 11 as well as the unplaced scaffold 380. Genes distributed on chromosome 1 were also syntenic with genes on chromosomes 3, 6, and 18 (Supplementary File S1). Overall, the results showed that syntenic genes from chromosomes 1, 6, and 18 were the most shared genes. These results indicate that a duplication event was crucial for the evolution of PdPFK genes in P. deltoides.
3.4. Conserved Motifs and Gene Structure Analysis of the PdPFP and PdPFK Gene Families
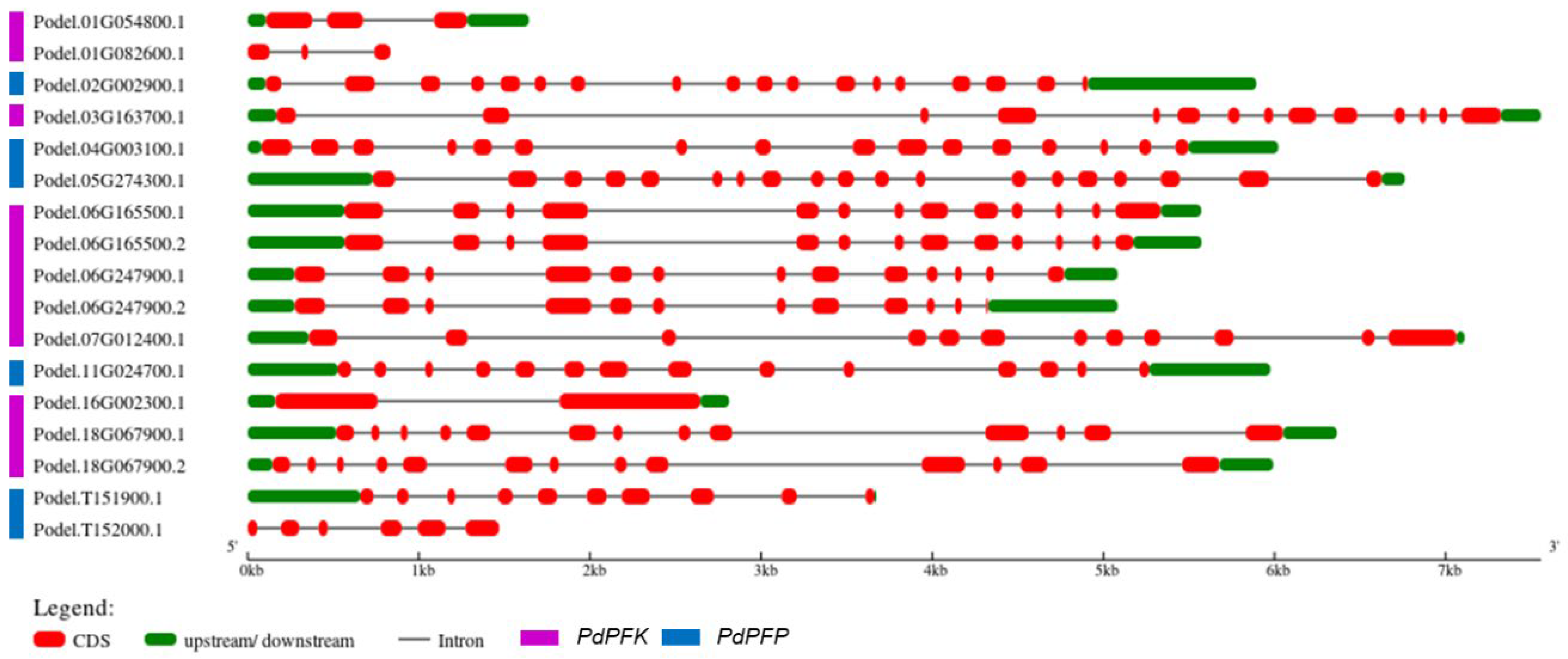
By analyzing the structural exon–intron distribution in P. deltoides, the gene architecture of PdPFPs and PdPFKs was revealed (Figure 4). The gene structure reflects the evolutionary relationship between PdPFK gene families. Comparing the length of the genes, PdPFPs were mostly longer than other genes, with an average length of greater than 5 kb. In comparison, PdPFPs had higher exon numbers than PdPFKs, especially Podel.05G274300.1, Podel.02G002900, and Podel.04G003100.1, which contained 19, 18, and 16 exons, respectively. The analysis of the motifs in PFK proteins was carried out using the MEME suite (Figure 5, Table 2). Of the 17 proteins subjected to motif analysis, only 14 showed motif blocks and 3 did not. A total of ten motifs were analyzed, of which nine were observed in seven proteins. Two pro-proteins lacked motif 8, whereas five proteins only had four motifs. The number of conserved motifs was larger in PdPFKs than in PdPFPs. A common motif in PdPFPs was motif 1.
3.5. Phylogenetic Analysis of the PdPFP and PdPFK Gene Families
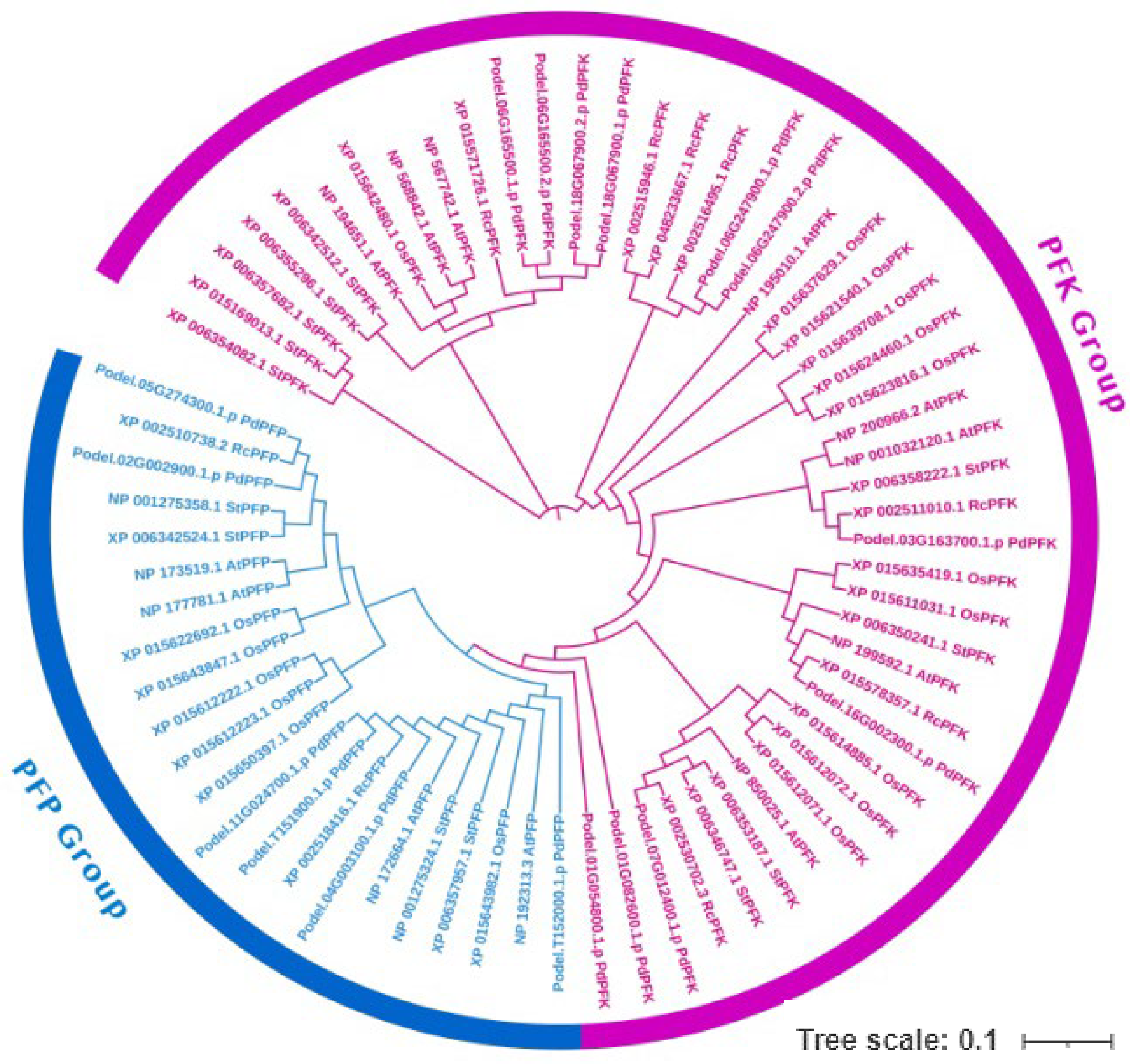
The evolutionary analysis and patterns of the PFKs were compared with four species: A. thaliana, R. communis, S. tuberosum, and O. sativa (Figure 6). The phylogenetic tree clustered into two groups. ClustalW was used to align the proteins of all five species, including P. deltoides. A total of six PdPFPs were strongly clustered with four AtPFPs, two RcPFPs, and four StPFPs, while 11 PdPFKs were clustered with eight AtPFKs, seven RcPFKs, and 11 StPFKs. The number of PdPFPs was higher in P. deltoides than in the other species. The phylogenetic tree was constructed using iTOLv6, EMBL.
3.6. Analysis of PdPFP and PdPFK Expression Patterns
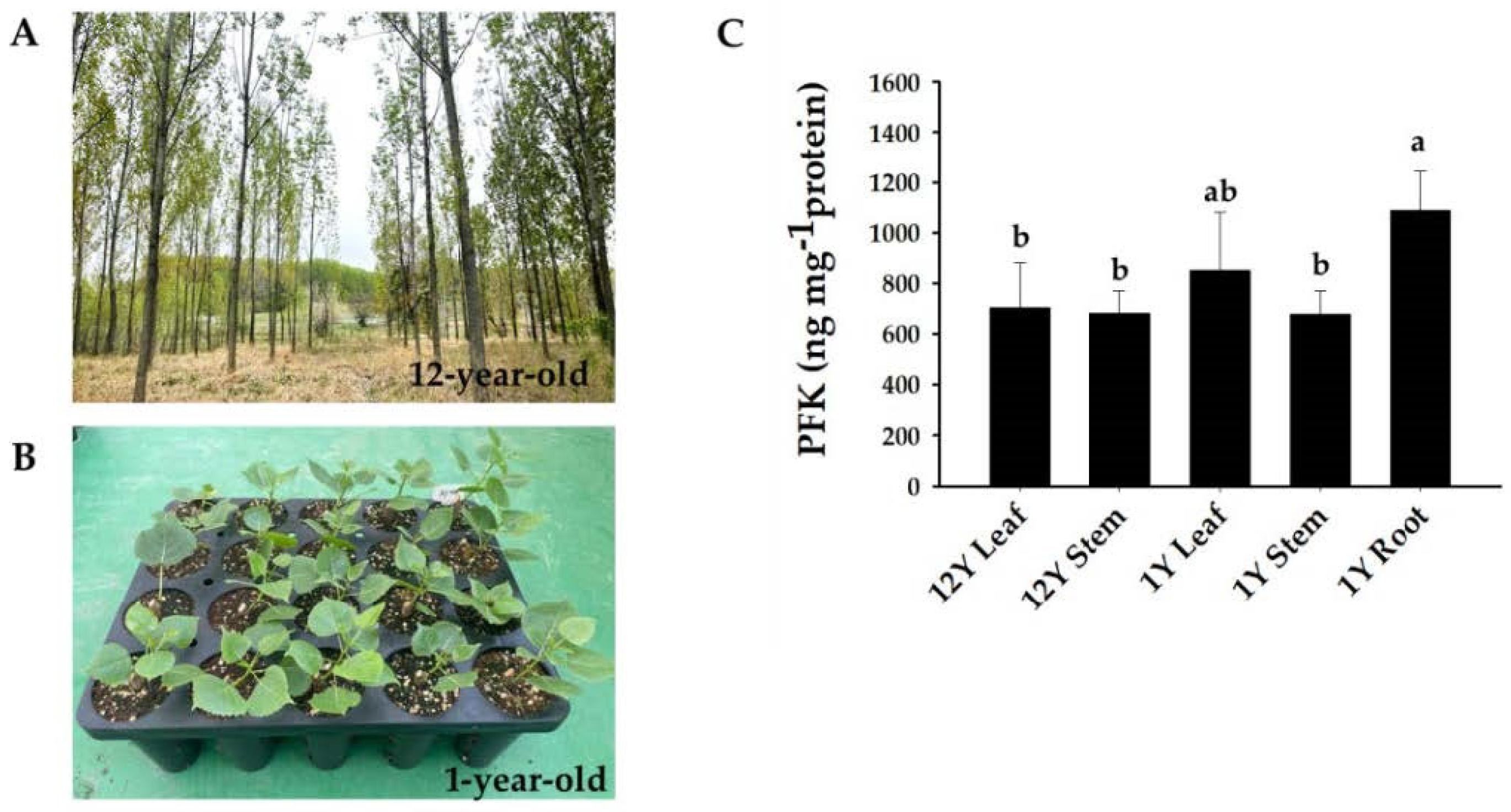
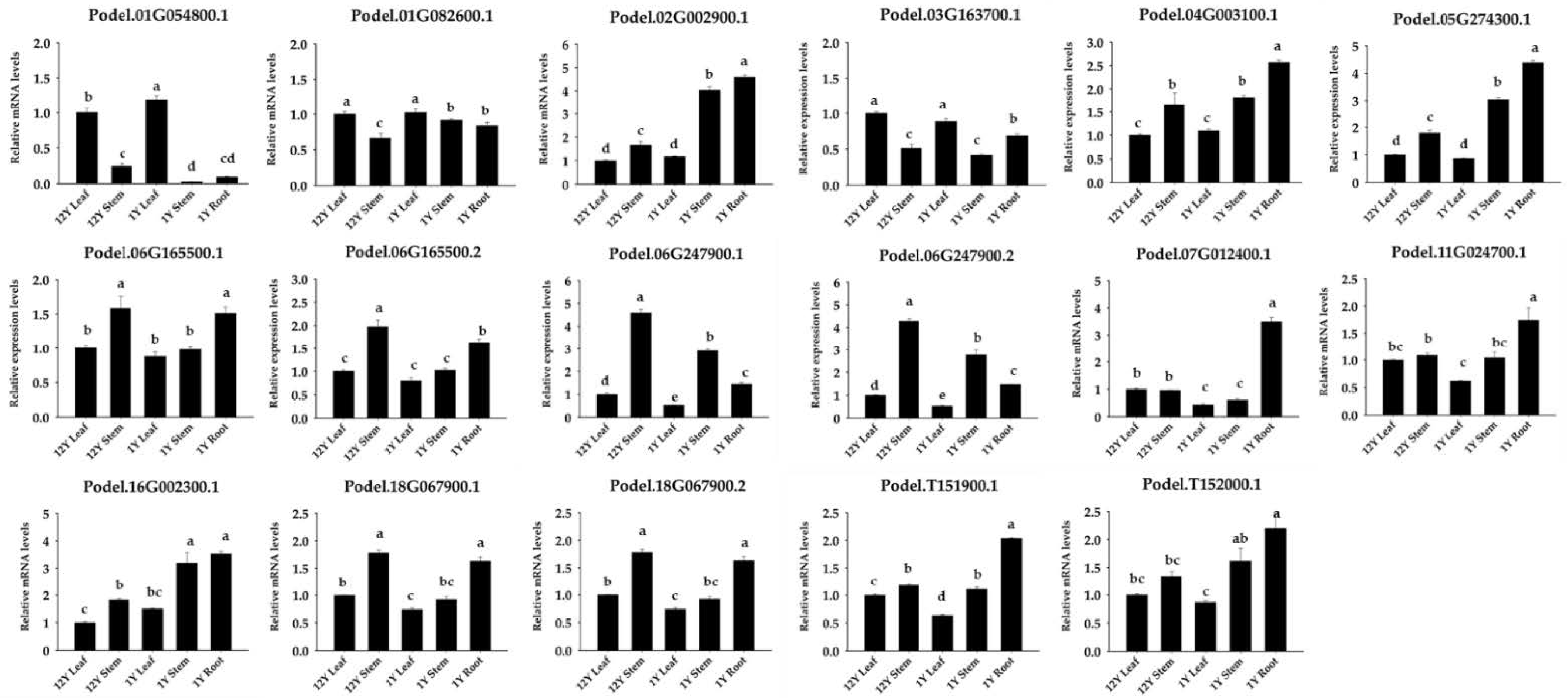
To investigate the potential role of phosphofructokinase, the PFK content was measured in P. deltoides. PdPFK showed high content in all tissues, particularly in roots (Figure 7). To further investigate the expression of PFP and PFK genes in different tissues and ages, transcriptional analysis was conducted in P. deltoides. qPCR was utilized to assess gene expression and provide insights into the functions of PdPFP and PdPFK genes in different tissues and developmental stages of poplar. Phosphofructokinase is known to play a role in diverse processes, including growth and the response to biotic and abiotic stresses [10]. Consistent with the protein assay findings, the expression of all 17 phosphofructokinase genes was detected in various tissues and ages of P. deltoides (Figure 8). The PdPFP genes Podel.02G002900.1, Podel.04G003100.1, Podel.05G274300.1, Podel.11G024700.1, Podel.T151900.1, and Podel.T152000.1 showed similar patterns in different tissues and ages, being more similar to PFK content patterns (Figure 8). This suggests that PdPFP is essential in the glycolytic pathway of poplar.
This study confirmed the tissue-specific expression patterns of phosphofructokinase genes. Podel.01G054800.1, Podel.01G082600.1, and Podel.03G163700.1 were expressed specifically in leaves, while Podel.06G247900.1 and Podel.06G247900.2 were expressed specifically in steams. In addition, Podel.05G274300.1, Podel.07G012400.1, Podel.11G024700.1, and Podel.T151900.1 were expressed specifically in roots. Podel.02G002900.1, Podel.05G274300.1, Podel.16G002300.1, and Podel.T152000.1 showed similar patterns of strong expression in one-year-old stems and roots (Figure 8). These genes may be used for the production of compounds that are beneficial for specific plant organs.
4. Discussion
Phosphofructokinase is essential in plant growth, metabolism, and stress responses; however, further research is required to confirm its exact mechanism and function. In particular, it has yet to be reported in poplar, which is a tree model plant. As far as we are aware, this study is the first to identify phosphofructokinase in P. deltoides through genome analysis.
We identified 17 PFK gene families in P. deltoides, with most PFK genes (11 genes) and only 6 genes belonging to the PFP subfamily. A similar division of the gene family into PFK and PFP groups was found in the other plants studied. The PFK gene family in A. thaliana consists of eleven members, with four members associated with AtPFP and seven members associated with AtPFK. Five of the fifteen PFK genes found in rice belonged to OsPFP and ten to OsPFK. Ten of the fourteen members of the PFK family in white pears (P. bretschneideri) are PbPFKs, while the other four are PbPFPs [31]. In cotton, 56 PFKs and 24 PFPs have been detected in 4 Gossypium species [32].
Based on our analysis of the subcellular localization of PFK genes, we found that of the six PdPFPs, three PdPFPs, and most PdPFKs, were localized in the cytoplasm. Four PdPFKs and one PdPFP were localized in the chloroplasts. It was hypothesized that five MePFKs are localized in the chloroplasts, two in the cytoplasm, and four in the cytoplasm of Manihot esculenta. To provide evidence for this assertion, MePFK03 and MePFPA1, which are presumed to be localized in the chloroplast and cytoplasm, respectively, were selected for the production of GFP fusion proteins. The localization of PFK and PFP in the cytoplasm and chloroplasts was confirmed using GFP images. Therefore, most PFKs and PFPs are expected to be in the cytoplasm and chloroplasts [32].
Under normal growth conditions, the cellular sugar status is essentially maintained but severely affected by numerous environmental stresses. One such negative environmental stress is drought, which reduces the cell’s osmotic potential and hampers the photosynthetic fixation of carbon and the transportation of sugars. A specific category of proteins, referred to as sugar transporters, plays a crucial role in facilitating the transportation of cellular sugars. The transporter proteins play vital roles in regulating the influx and efflux of diverse sugars and their metabolic intermediates, thereby facilitating plant growth and development. Abiotic stresses, particularly drought stress, lead to the reprogramming of sugar distribution within cellular and subcellular compartments [33]. PFP is tightly controlled; however, its purpose remains unknown. There is no evidence that PFP controls glycolysis in transgenic potato plants, where PFP activity is reduced by more than 90%. However, PFP is implicated in the capacity of plants to adapt to adverse conditions, such as phosphate or oxygen deficiency [7].
In this study, we confirmed that PdPFK and PdPFP show tissue- or development-specific expression. We specifically looked into the function of PdPFPs during P. deltoides’ developmental stages. Examination of the expression of PFKs and PFPs clearly showed that compared with PdPFKs, the expression of PdPFP was higher in developing P. deltoides. We found that the expression of PdPFPs, such as Podel.02G002900.1, Podel.04G003100.1, Podel.05G274300.1, Podel.11G024700.1, Podel.T151900.1, and Podel.T152000.1, was higher in roots than in other tissues. Previous studies have documented that PdPFPs are upregulated in conjunction with heat shock proteins (HSPs) when maize endosperm is subjected to heat, cold, and unfolded protein response stresses [34]. In Arabidopsis, the expression of PFPα1 was found to be higher than PFPα2 in leaves, flowers, and roots, while the PFPβ2 gene exhibited predominant expression in leaves [35]. Moreover, AtPFPα2 had a trichome-specific expression promoter [36]. The expression levels of PdPFP and PdPFK in the stems differed according to age and developmental stage. In particular, the expression levels of Podel.02G002900.1 and Podel.06G165500.2 were more than twice as high according to age.
5. Conclusions
This study provided a bioinformatic analysis to identify 17 phosphofructokinase genes, which were further classified into 2 subfamilies based on phylogenetic analysis. The PFK subfamily comprised 11 genes, while the PFP subfamily consisted of 6 genes. The subfamilies exhibited relatively conserved characteristics in terms of motif composition and exon/intron structures. The conserved motif 1, composed of 50 amino acids, was the center, and 8 motifs of PFK and 4 motifs of PFP constituted the protein. Phosphofructokinase was unevenly distributed in the chromosomes and showed high synteny, especially on chromosomes 1, 6, and 18. Additionally, the expression patterns of PFKs in various tissues of P. deltoides were analyzed, revealing high gene expression and protein concentrations in the roots. This study establishes a foundation for investigating the PdPFK and PdPFP gene families in poplar and provides valuable insights into their molecular mechanisms.
Supplementary Materials
The following supporting information can be downloaded from https://www.mdpi.com/article/10.3390/f14061104/s1: Table S1: Sequences of primers used for qPCR in P. deltoides; File S1: Collinearity of P. deltoides.
Author Contributions
Conceptualization, T.-L.K. and H.L; methodology, H.L and H.C.; software, M.I.J.D. and H.C.; validation, T.-L.K. and M.I.J.D.; formal analysis, T.-L.K. and M.I.J.D.; investigation, T.-L.K.; resources, H.L. and C.O.; data curation, T.-L.K. and M.I.J.D.; writing—original draft preparation, T.-L.K. and M.I.J.D.; writing—review and editing, H.L.; visualization, T.-L.K. and M.I.J.D.; supervision, H.L.; project administration, H.L.; funding acquisition, H.L. and C.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Institute of Forest Science of the Republic of Korea, grant number FG0402-2022-01-2023.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Plaxton, W.C. The organization and regulation of plant glycolysis. Annu. Rev. Plant Biol. 1996, 47, 185–214. [Google Scholar] [CrossRef] [PubMed]
- Mertens, E.; Larondelle, Y.; Hers, H.-G. Induction of pyrophosphate: Fructose 6-phosphate 1-phosphotransferase by anoxia in rice seedlings. Plant Physiol. 1990, 93, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Claassen, P.A.; Budde, M.A.; de Ruyter, H.J.; van Calker, M.H.; van Es, A. Potential role of pyrophosphate: Fructose 6-phosphate phosphotransferase in carbohydrate metabolism of cold stored tubers of Solanum tuberosum cv Bintje. Plant Physiol. 1991, 95, 1243–1249. [Google Scholar] [CrossRef]
- Lim, H.; Cho, M.-H.; Bhoo, S.H.; Hahn, T.-R. Pyrophosphate: Fructose-6-phosphate 1-phosphotransferase is involved in the tolerance of Arabidopsis seedlings to salt and osmotic stresses. In Vitro Cell Dev. Biol.-Plant 2014, 50, 84–91. [Google Scholar] [CrossRef]
- Nielsen, M.F.; Caumo, A.; Chandramouli, V.; Schumann, W.C.; Cobelli, C.; Landau, B.R.; Vilstrup, H.; Rizza, R.A.; Schmitz, O. Impaired basal glucose effectiveness but unaltered fasting glucose release and gluconeogenesis during short-term hypercorti-solemia in healthy subjects. Am. J. Physiol.-Endocrinol. Metab. 2004, 286, E102–E110. [Google Scholar] [CrossRef]
- Dennis, D.T.; Greyson, M.F. Fructose 6-phosphate metabolism in plants. Physiol. Plant 1987, 69, 395–404. [Google Scholar] [CrossRef]
- Mustroph, A.; Sonnewald, U.; Biemelt, S. Characterisation of the ATP-dependent phosphofructokinase gene family from Ara-bidopsis thaliana. FEBS Lett. 2007, 581, 2401–2410. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, J.; Chen, Y.; Pan, H.; Ming, R. Identification and genes expression analysis of ATP-dependent phosphofructo-kinase family members among three Saccharum species. Funct. Plant Biol. 2012, 40, 369–378. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. The catalytic direction of pyrophosphate: Fructose 6-phosphate 1-phosphotransferase in rice coleoptiles in anoxia. Physiol. Plant. 2002, 116, 345–350. [Google Scholar] [CrossRef]
- Lü, H.; Li, J.; Huang, Y.; Zhang, M.; Zhang, S.; Wu, J. Genome-wide identification, expression and functional analysis of the phosphofructokinase gene family in Chinese white pear (Pyrus bretschneideri). Gene 2019, 702, 133–142. [Google Scholar] [CrossRef]
- Stitt, M. Fructose-2, 6-bisphosphate as a regulatory molecule in plants. Annu. Rev. Plant Biol. 1990, 41, 153–185. [Google Scholar] [CrossRef]
- Hajirezaei, M.; Sonnewald, U.; Viola, R.; Carlisle, S.; Dennis, D.; Stitt, M. Transgenic potato plants with strongly decreased expression of pyrophosphate: Fructose-6-phosphate phosphotransferase show no visible phenotype and only minor changes in metabolic fluxes in their tubers. Planta 1993, 192, 16–30. [Google Scholar] [CrossRef]
- Spilatro, S.R.; Anderson, J.M. Carbohydrate metabolism and activity of pyrophosphate: Fructose-6-phosphate phosphotrans-ferase in photosynthetic soybean (Glycine max, Merr.) suspension cells. Plant Physiol. 1988, 88, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-P.; Sung, S.-J.S.; Black, C.C. Sucrose metabolism in lima bean seeds. Plant Physiol. 1989, 89, 1106–1116. [Google Scholar] [CrossRef]
- Kruger, N.J.; Dennis, D.T. Molecular properties of pyrophosphate: Fructose-6-phosphate phosphotransferase from potato tuber. Arch. Biochem. Biophys. 1987, 256, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Shi, J.-N. Evidence that fructose 1, 6-bisphosphate specifically protects the α-subunit of pyrophosphate-dependent 6-phosphofructo-1-phosphotransferase against proteolytic degradation. FEBS Lett. 1999, 459, 448–452. [Google Scholar] [CrossRef]
- Guo, X.; Ronhovde, K.; Yuan, L.; Yao, B.; Soundararajan, M.P.; Elthon, T.; Zhang, C.; Holding, D.R. Pyrophosphate-dependent fructose-6-phosphate 1-phosphotransferase induction and attenuation of Hsp gene expression during endosperm modification in quality protein maize. Plant Physiol. 2012, 158, 917–929. [Google Scholar] [CrossRef]
- Podestá, F.E.; Plaxton, W.C. Fluorescence study of ligand binding to potato tuber pyrophosphate-dependent phosphofructo-kinase: Evidence for competitive binding between fructose-1, 6-bisphosphate and fructose-2, 6-bisphosphate. Arch. Biochem. Biophys. 2003, 414, 101–107. [Google Scholar] [CrossRef]
- Theodorou, M.; Cornel, F.; Duff, S.; Plaxton, W. Phosphate starvation-inducible synthesis of the alpha-subunit of the pyro-phosphate-dependent phosphofructokinase in black mustard suspension cells. J. Biol. Chem. 1992, 267, 21901–21905. [Google Scholar] [CrossRef]
- Wong, J.H.; Kiss, F.; Wu, M.-X.; Buchanan, B.B. Pyrophosphate fructose-6-P 1-phosphotransferase from tomato fruit: Evidence for change during ripening. Plant Physiol. 1990, 94, 499–506. [Google Scholar] [CrossRef]
- Huang, S.; Colmer, T.D.; Millar, A.H. Does anoxia tolerance involve altering the energy currency towards PPi? Trends Plant Sci. 2008, 13, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Van Schaftingen, E.; Hers, H.-G. Fructose 2, 6-bisphosphate in relation with the resumption of metabolic activity in slices of Jerusalem artichoke tubers. FEBS Lett. 1983, 164, 195–200. [Google Scholar] [CrossRef]
- Fahrenkrog, A.M.; Neves, L.G.; Resende, M.F., Jr.; Dervinis, C.; Davenport, R.; Barbazuk, W.B.; Kirst, M. Population genomics of the eastern cottonwood (Populus deltoides). Ecol. Evol. 2017, 7, 9426–9440. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Wu, H.; Zhang, J.; Pan, Z.; Zhao, W.; Li, Z.; Tong, C. Genome assembly of Salicaceae Populus deltoides (Eastern Cottonwood) I-69 based on nanopore sequencing and Hi-C technologies. J. Hered. 2021, 112, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for inter-active analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for de-tection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pettengill, E.A.; Parmentier-Line, C.; Coleman, G.D. Evaluation of qPCR reference genes in two genotypes of Populus for use in photoperiod and low-temperature studies. BMC Res. Notes 2012, 5, 366. [Google Scholar]
- Eddy, S.R. Bioinformatics Review. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, P.; Shen, X.; Xia, Z.; Zhou, X.; Chen, X.; Lu, C. Genome-wide survey of the phosphofructokinase family in cassava and functional characterization in response to oxygen-deficient stress. BMC Plant Biol. 2021, 21, 375. [Google Scholar] [CrossRef] [PubMed]
- Mehari, T.G.; Xu, Y.; Umer, M.J.; Hui, F.; Cai, X.; Zhou, Z.; Hou, Y.; Wang, K.; Wang, B.; Liu, F. Genome-Wide Identification and Expression Analysis Elucidates the Potential Role of PFK Gene Family in Drought Stress Tolerance and Sugar Metabolism in Cotton. Front. Genet. 2022, 13, 922024. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Manna, M.; Thakur, T.; Gautam, V.; Salvi, P. Imperative role of sugar signaling and transport during drought stress responses in plants. Physiol. Plant. 2021, 171, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Basson, C.; Groenewald, J.-H.; Kossmann, J.; Cronjé, C.; Bauer, R. Upregulation of pyrophosphate: Fructose 6-phosphate 1-phosphotransferase (PFP) activity in strawberry. Transgenic Res. 2011, 20, 925–931. [Google Scholar] [CrossRef]
- Lim, H.; Cho, M.-H.; Jeon, J.-S.; Bhoo, S.H.; Kwon, Y.-K.; Hahn, T.-R. Altered expression of pyrophosphate: Fruc-tose-6-phosphate 1-phosphotransferase affects the growth of transgenic Arabidopsis plants. Mol. Cells 2009, 27, 641–649. [Google Scholar] [CrossRef]
- Lim, H.-M.; Cho, J.-I.; Lee, S.; Cho, M.-H.; Bhoo, S.H.; An, G.; Hahn, T.-R.; Jeon, J.-S. Identification of a 20-bp regulatory element of the Arabidopsis pyrophosphate: Fructose-6-phosphate 1-phosphotransferase α2 gene that is essential for expression. Plant Cell Rep. 2007, 26, 683–692. [Google Scholar] [CrossRef]
Figure 1.
Prediction of subcellular localization.
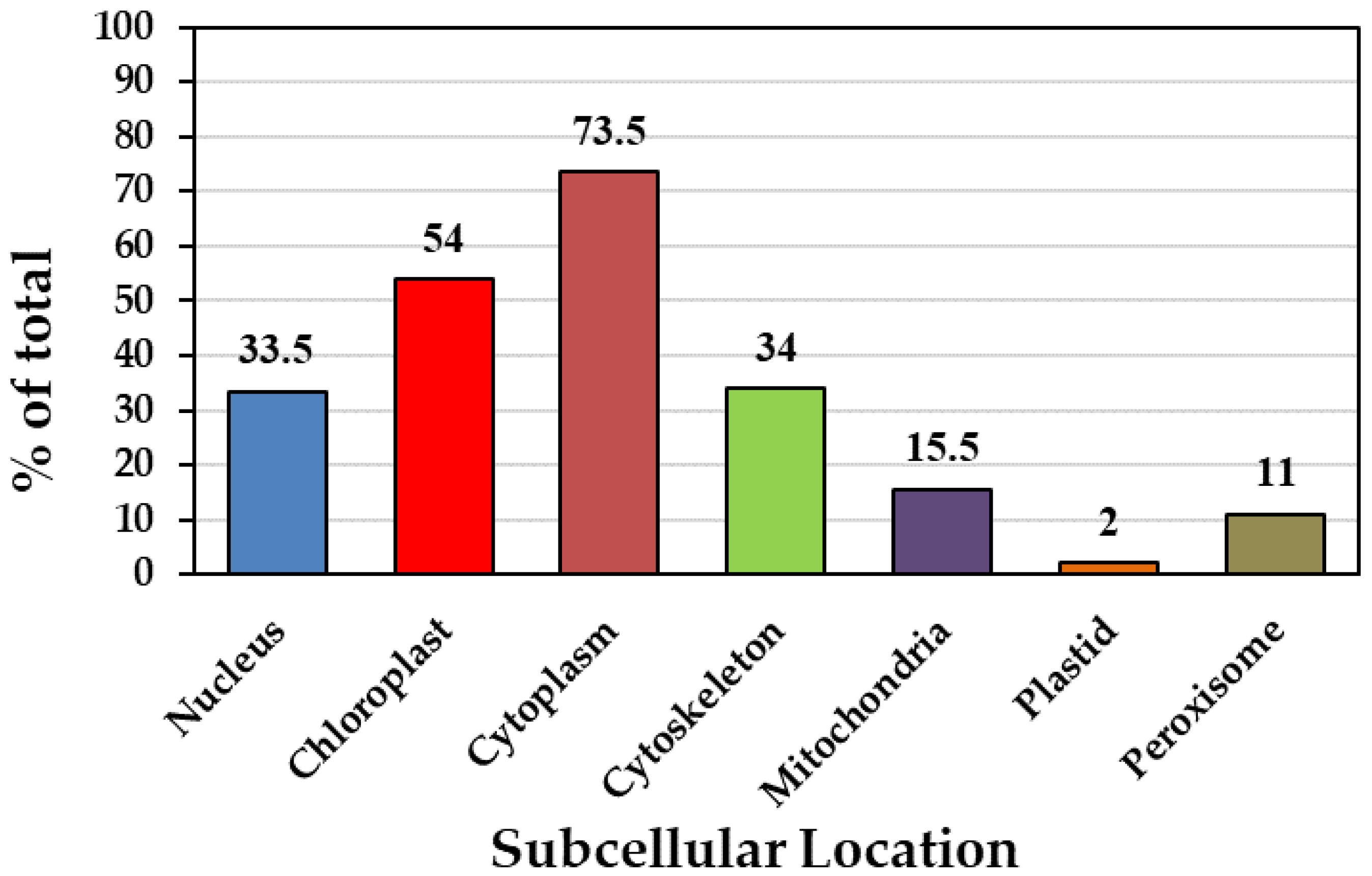
Figure 2.
Chromosome location of phosphofructokinase gene family in P. deltoides. The length of chromosomes can be estimated with the scale on the side. The chromosome map was constructed using TBTools.
Figure 2.
Chromosome location of phosphofructokinase gene family in P. deltoides. The length of chromosomes can be estimated with the scale on the side. The chromosome map was constructed using TBTools.

Figure 3.
Collinearity analysis between phosphofructokinase genes in P. deltoides. The red lines indicate the collinearity of PFK genes in the poplar genome. The inner pale yellow layer represents the poplar chromosomes, where Chr stands for chromosomes, and the chromosome number can be found next to it. The length of each poplar chromosome is expressed in units of Mb with the labels of collinear genes. The middle layer represents the distribution of genes on individual poplar species chromosomes. The third, outermost layer represents the percentage of GC on each chromosome. The collinearity was identified using MCScanX and the circular plot was drawn using TBTools.
Figure 3.
Collinearity analysis between phosphofructokinase genes in P. deltoides. The red lines indicate the collinearity of PFK genes in the poplar genome. The inner pale yellow layer represents the poplar chromosomes, where Chr stands for chromosomes, and the chromosome number can be found next to it. The length of each poplar chromosome is expressed in units of Mb with the labels of collinear genes. The middle layer represents the distribution of genes on individual poplar species chromosomes. The third, outermost layer represents the percentage of GC on each chromosome. The collinearity was identified using MCScanX and the circular plot was drawn using TBTools.
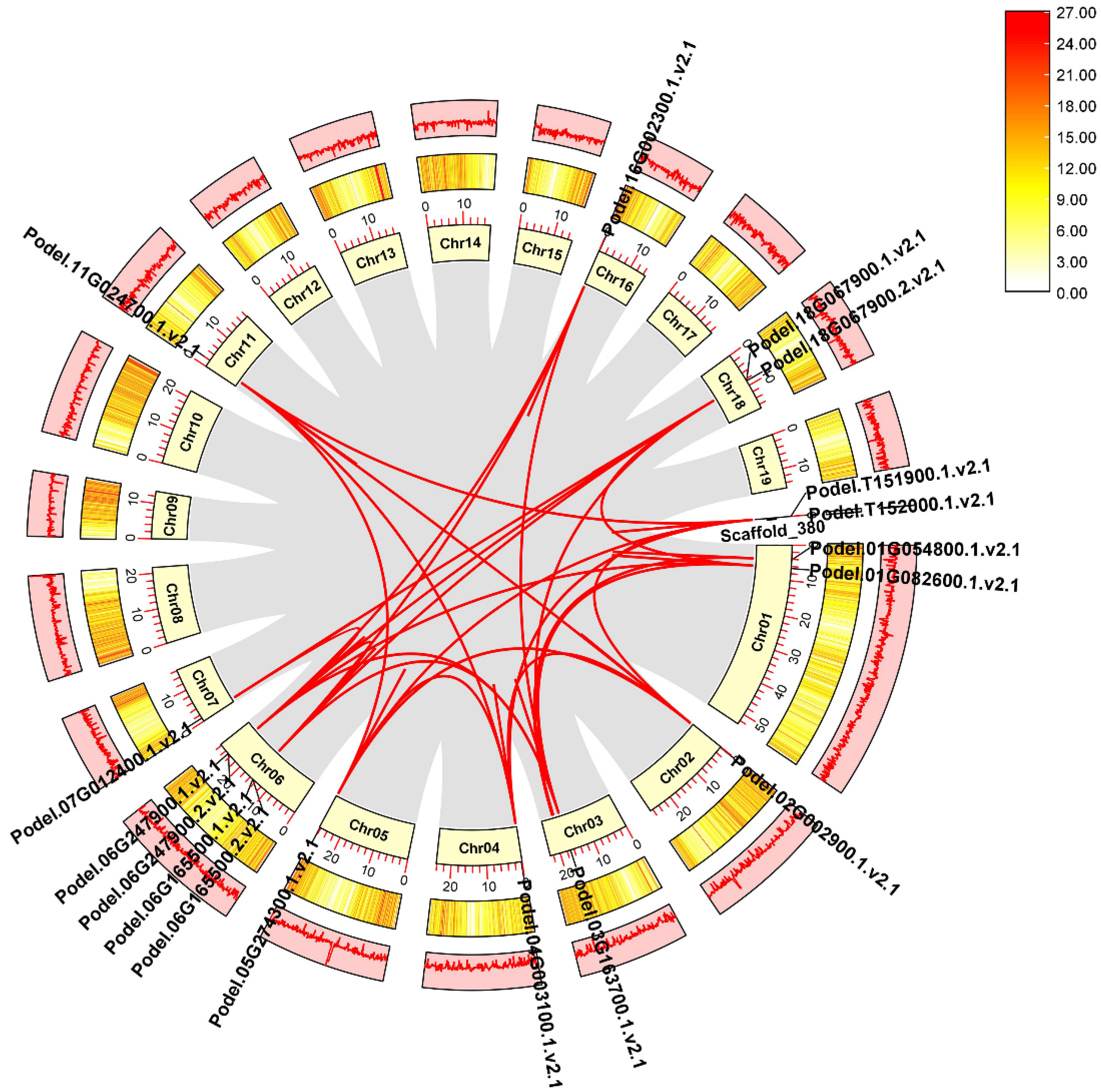
Figure 4.
Gene structures of phosphofructokinase gene family in P. deltoides. The exon–intron structure was constructed using the Gene Structure Display Server (GSDS 2.0) (http://gsds.gao-lab.org/, accessed on 18 April 2023).
Figure 4.
Gene structures of phosphofructokinase gene family in P. deltoides. The exon–intron structure was constructed using the Gene Structure Display Server (GSDS 2.0) (http://gsds.gao-lab.org/, accessed on 18 April 2023).

Figure 5.
Analysis of conserved phosphofructokinase gene family motifs in P. deltoides. Motifs were identified using the MEME suite available online (https://meme-suite.org/meme/tools/meme, accessed on 19 April 2023). There are 10 motifs in total, which are shown in different colors. The number of motifs in PdPFKs is higher than in PdPFPs.
Figure 5.
Analysis of conserved phosphofructokinase gene family motifs in P. deltoides. Motifs were identified using the MEME suite available online (https://meme-suite.org/meme/tools/meme, accessed on 19 April 2023). There are 10 motifs in total, which are shown in different colors. The number of motifs in PdPFKs is higher than in PdPFPs.

Figure 6.
Phylogenetic analysis of phosphofructokinase genes of poplar and four other species (At—Arabidopsis thaliana, Rc—Ricinus communis, St—Solanum tuberosum, and Os—Oryza sativa). The two members of the PFK gene family are shown in two different colors. Pink and blue color represent the PFK and PFP members respectively. The phylogenetic tree was constructed using the iTOL server (v6), and the visualization was improved using Adobe Photoshop 2020. The tree scale bar represents 0.1 substitutions per amino acid.
Figure 6.
Phylogenetic analysis of phosphofructokinase genes of poplar and four other species (At—Arabidopsis thaliana, Rc—Ricinus communis, St—Solanum tuberosum, and Os—Oryza sativa). The two members of the PFK gene family are shown in two different colors. Pink and blue color represent the PFK and PFP members respectively. The phylogenetic tree was constructed using the iTOL server (v6), and the visualization was improved using Adobe Photoshop 2020. The tree scale bar represents 0.1 substitutions per amino acid.
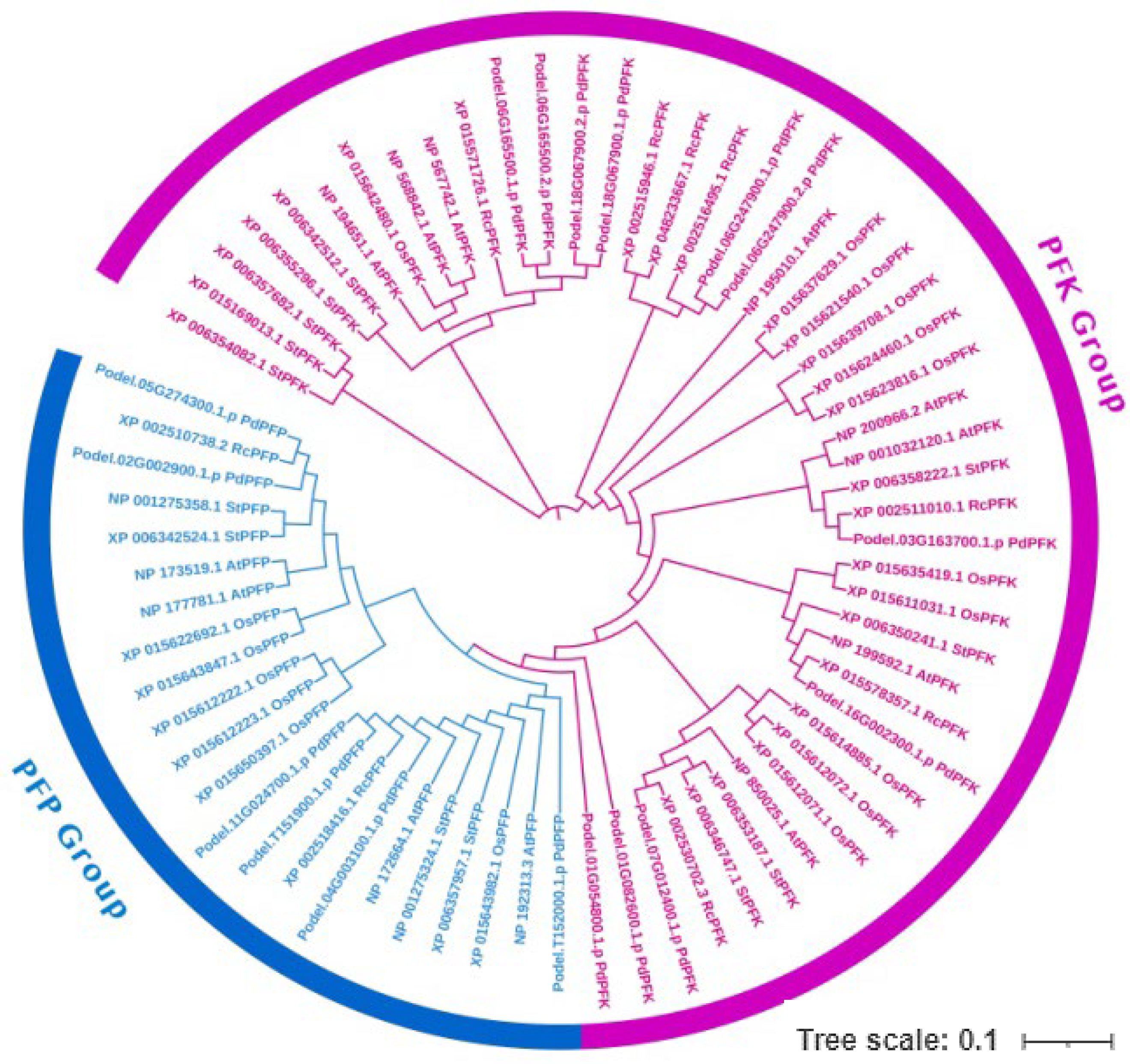
Figure 7.
(A) Growth phenotypes of P. deltoides trees 12 years after planting. (B) Growth phenotypes of P. deltoides one year after planting. (C) Analysis of the phosphofructokinase (PFK) contents in different tissues (leaf, stem, and root) and ages (12-year-old and 1-year-old trees) of P. deltoides. The data are represented as means ± SD of triplicate experiments. Significant differences are denoted with different lowercase letters (ANOVA with Tukey’s HSD, p < 0.05).
Figure 7.
(A) Growth phenotypes of P. deltoides trees 12 years after planting. (B) Growth phenotypes of P. deltoides one year after planting. (C) Analysis of the phosphofructokinase (PFK) contents in different tissues (leaf, stem, and root) and ages (12-year-old and 1-year-old trees) of P. deltoides. The data are represented as means ± SD of triplicate experiments. Significant differences are denoted with different lowercase letters (ANOVA with Tukey’s HSD, p < 0.05).
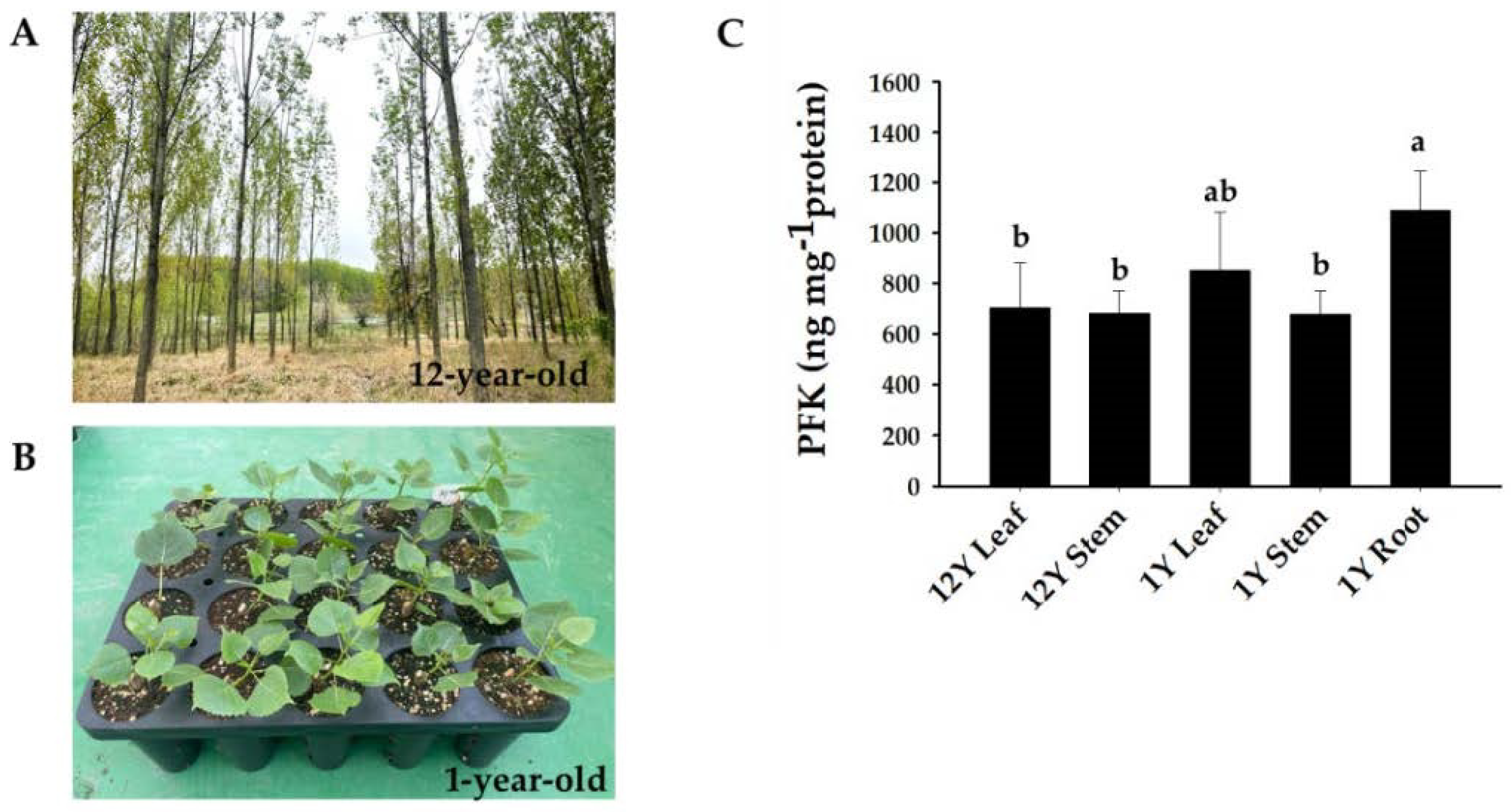
Figure 8.
Expression analysis of 17 phosphofructokinase genes in different tissues (leaf, stem, and root tissues) and ages (12-year-old and 1-year-old trees) of P. deltoides. The statistically significant differences between expression levels of phosphofructokinase genes were tested using Tukey’s test (p < 0.05). Error bars indicate the standard deviation calculated from three biological replicates. Different lowercase letters indicate significant differences (p < 0.05).
Figure 8.
Expression analysis of 17 phosphofructokinase genes in different tissues (leaf, stem, and root tissues) and ages (12-year-old and 1-year-old trees) of P. deltoides. The statistically significant differences between expression levels of phosphofructokinase genes were tested using Tukey’s test (p < 0.05). Error bars indicate the standard deviation calculated from three biological replicates. Different lowercase letters indicate significant differences (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physicochemical properties of phosphofructokinase family members in P. deltoides.
| Gene Name | Transcript ID | CDS (bp) | Protein Length (aa) | Molecular Weight (KDa) | Theoretical PI | Gravy | Predicted Localization |
|---|---|---|---|---|---|---|---|
| Podel.01G054800.1.p | Podel.01G054800.1 | 672 | 223 | 25.08 | 8.59 | −0.437 | Nucleus |
| Podel.01G082600.1.p | Podel.01G082600.1 | 261 | 86 | 9.74 | 8.21 | 0.029 | Chloroplast |
| Podel.02G002900.1.p | Podel.02G002900.1 | 1590 | 529 | 57.94 | 6.78 | −0.103 | Cytoplasm |
| Podel.03G163700.1.p | Podel.03G163700.1 | 1515 | 504 | 55.83 | 7.65 | −0.127 | Cytoplasm |
| Podel.04G003100.1.p | Podel.04G003100.1 | 1686 | 516 | 61.28 | 6.00 | −0.131 | Peroxisome |
| Podel.05G274300.1.p | Podel.05G274300.1 | 1854 | 617 | 67.24 | 6.39 | −0.115 | Cytoplasm |
| Podel.06G165500.1.p | Podel.06G165500.1 | 1659 | 552 | 60.69 | 6.16 | −0.217 | Chloroplast |
| Podel.06G165500.2.p | Podel.06G165500.2 | 1497 | 498 | 54.92 | 5.73 | −2.242 | Cytoplasm |
| Podel.06G247900.1.p | Podel.06G247900.1 | 1443 | 480 | 52.72 | 8.74 | −0.222 | Cytoplasm |
| Podel.06G247900.2.p | Podel.06G247900.2 | 1296 | 431 | 47.45 | 9.07 | −0.201 | Chloroplast |
| Podel.07G012400.1.p | Podel.07G012400.1 | 1599 | 532 | 58.34 | 6.83 | −0.112 | Cytoplasm |
| Podel.11G024700.1.p | Podel.11G024700.1 | 1284 | 427 | 47.20 | 6.24 | −0.22 | Cytoplasm |
| Podel.16G002300.1.p | Podel.16G002300.1 | 1419 | 472 | 52.08 | 5.82 | −0.161 | Cytoskeleton |
| Podel.18G067900.1.p | Podel.18G067900.1 | 1479 | 492 | 54.57 | 5.73 | −0.294 | Cytoskeleton |
| Podel.18G067900.2.p | Podel.18G067900.2 | 1479 | 492 | 54.57 | 5.73 | −0.294 | Cytoskeleton |
| Podel.T151900.1.p | Podel.T151900.1 | 948 | 315 | 34.77 | 6.56 | −0.121 | Mitochondria |
| Podel.T152000.1.p | Podel.T152000.1 | 699 | 232 | 24.50 | 5.71 | −0.189 | Chloroplast |
Molecular weight—molecular weight of the amino acid sequence; PI—isoelectric point of the PdPFKs; grand average of hydropathicity—hydrophilic mean; predicted localization—subcellular localization was predicted by analyzing the sequences.
Table 2.
Conserved motif information on phosphofructokinase family members in P. deltoides.
| Motif | Symbol | Length | Amino Acid Sequence Information |
|---|---|---|---|
| motif 1 |  | 50 | INAAHVEAESVENGIGLVKLMGRYSGFIAMEATLASRDVDCCLIPESPFY |
| motif 2 |  | 50 | YIDPTYMIRAVPPNASDNVYCTLLAQSAVHGAMAGYTGFTSGVVNGRQPY |
| motif 3 |  | 43 | HFRRAGPRQKVYFEPDEVRACIVTCGGLCPGLNTVIREIVYSL |
| motif 4 |  | 50 | LTPKVVNDIHKRGGTILGTSRGGHDTSKIVDSIQDRGINQVYIIGGDGTQ |
| motif 5 |  | 41 | EIRRRGLKVAVAGIPKTIDNDIPVIDRSFGFDTAVEEAQRA |
| motif 6 |  | 41 | LEGKGGLFEYIEKRLKENGHMVIVIAEGAGQELLSESMQSD |
| motif 7 |  | 50 | DVPHLTDYIPBLPTYSNPLQDNPAYSVVKQYFVHVDDSVPQKIVVHKDSP |
| motif 8 |  | 41 | IPFYRINEKQNKVVITDRMWARLLSSTNQPSFISNKEVIED |
| motif 9 |  | 29 | ASGNKLLQDVGLWISQGIKDYFSRQKKMA |
| motif 10 |  | 50 | NQSLQLFEFLPPAIQEQLMLERDPHGNVQVAKIETEKMLIQMVETELEKR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, T.-L.; Denison, M.I.J.; Lim, H.; Chung, H.; Oh, C. Genome-Wide Analysis, Identification, and Characterization of the PFK Gene Family Members of Populus deltoides. Forests 2023, 14, 1104. https://doi.org/10.3390/f14061104
AMA Style
Kim T-L, Denison MIJ, Lim H, Chung H, Oh C. Genome-Wide Analysis, Identification, and Characterization of the PFK Gene Family Members of Populus deltoides. Forests. 2023; 14(6):1104. https://doi.org/10.3390/f14061104
Chicago/Turabian StyleKim, Tae-Lim, Michael Immanuel Jesse Denison, Hyemin Lim, Hoyong Chung, and Changyoung Oh. 2023. "Genome-Wide Analysis, Identification, and Characterization of the PFK Gene Family Members of Populus deltoides" Forests 14, no. 6: 1104. https://doi.org/10.3390/f14061104
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.