
A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location
2.2. Sample Plot Setting and Investigation
2.3. Selection of Dominant Trees
2.4. Determination of Competing Trees
2.4.1. Sample Edge Correction
2.4.2. Determination of Competition Units for Dominant Trees
2.5. Soil Sample Collection
2.6. Data Processing
2.6.1. Construction of the Competition Index Model
2.6.2. Model Construction of Structural Equations
3. Results and Analysis
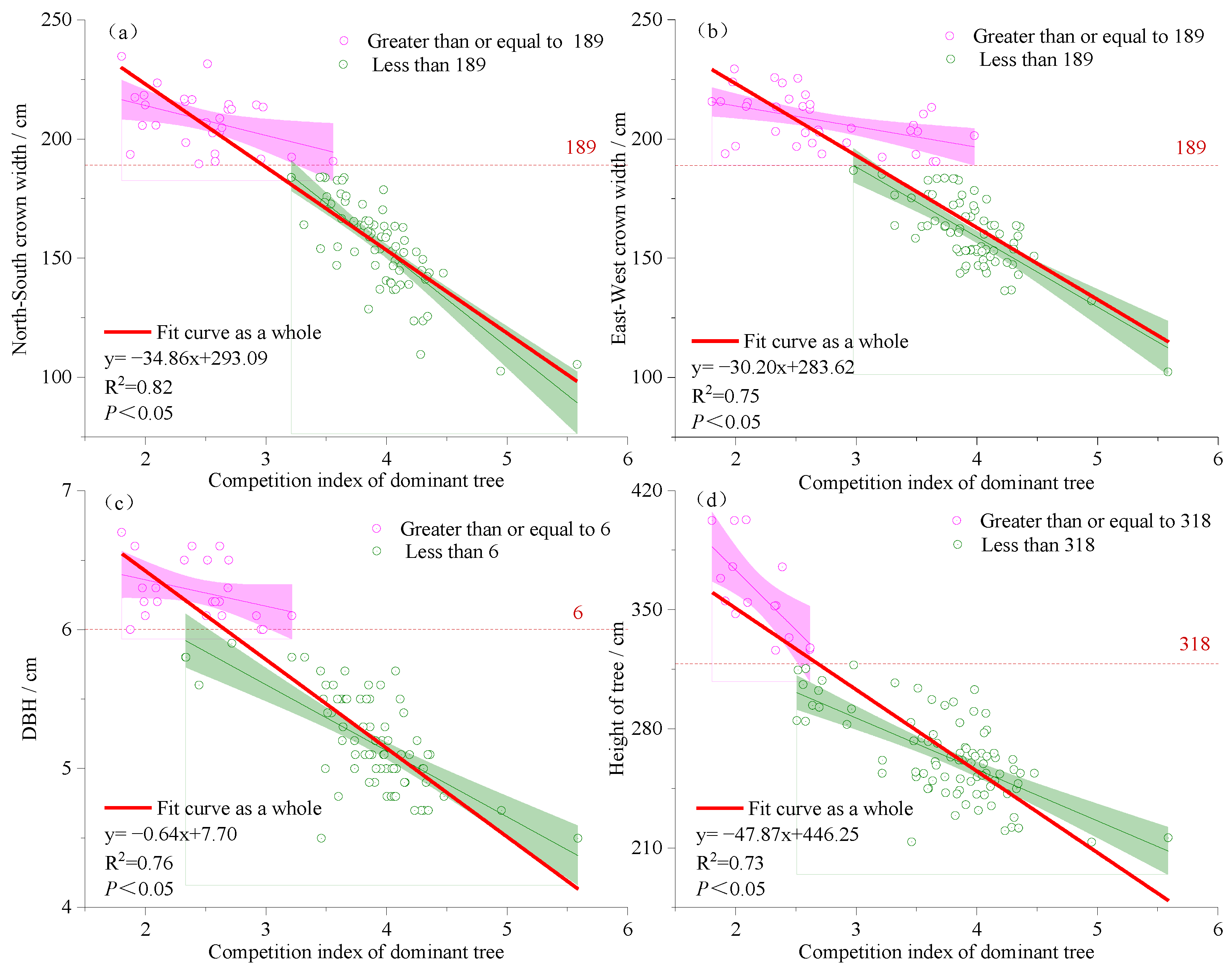
3.1. Fitting the Relationship between the Dominant Tree Competition Index and Stand Growth Factors
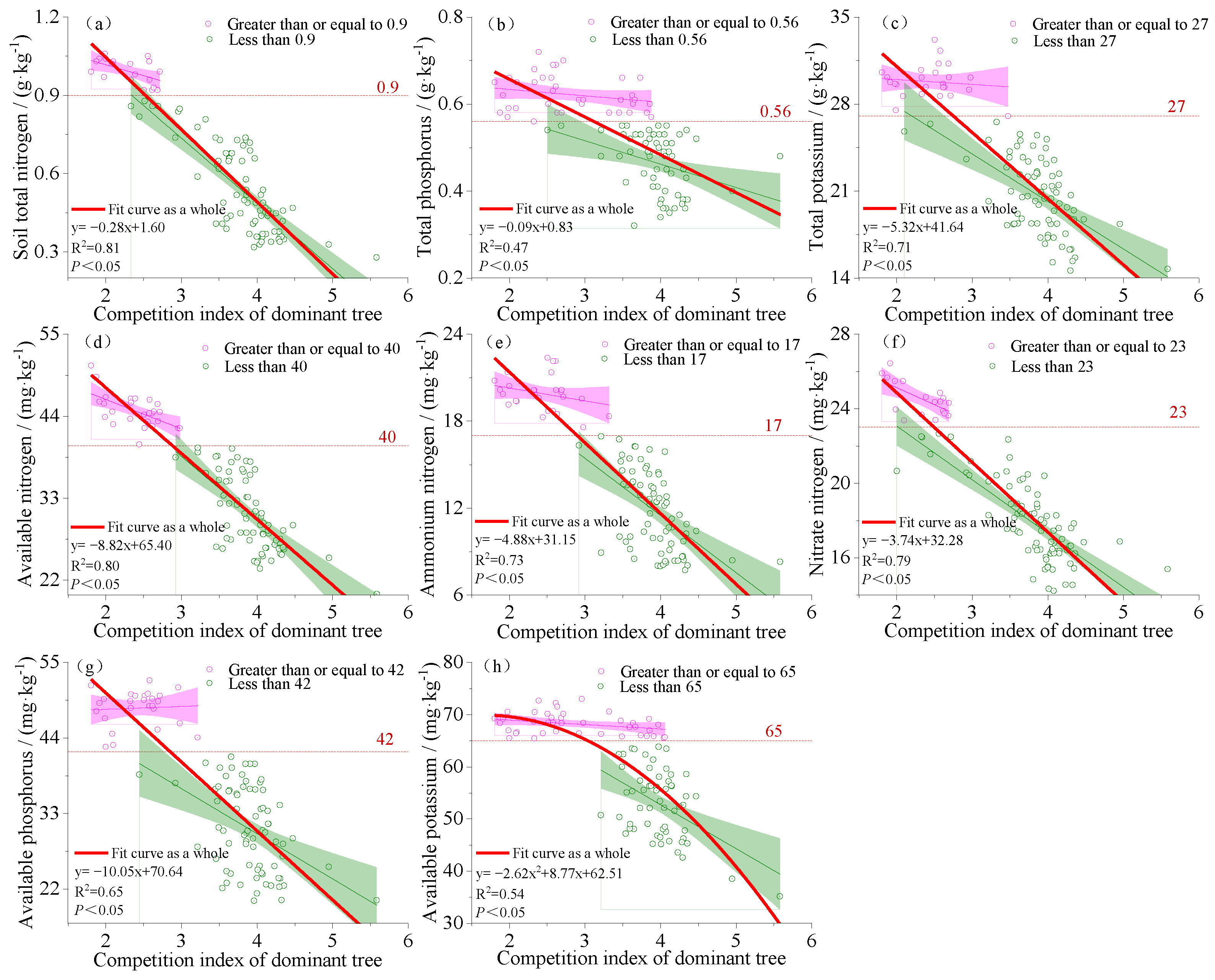
3.2. Fitting the Relationship between the Dominant Tree Competition Index and Soil Nutrient Factors
3.3. Fitting the Relationship between the Dominant Tree Competition Index and Soil Physical Factors
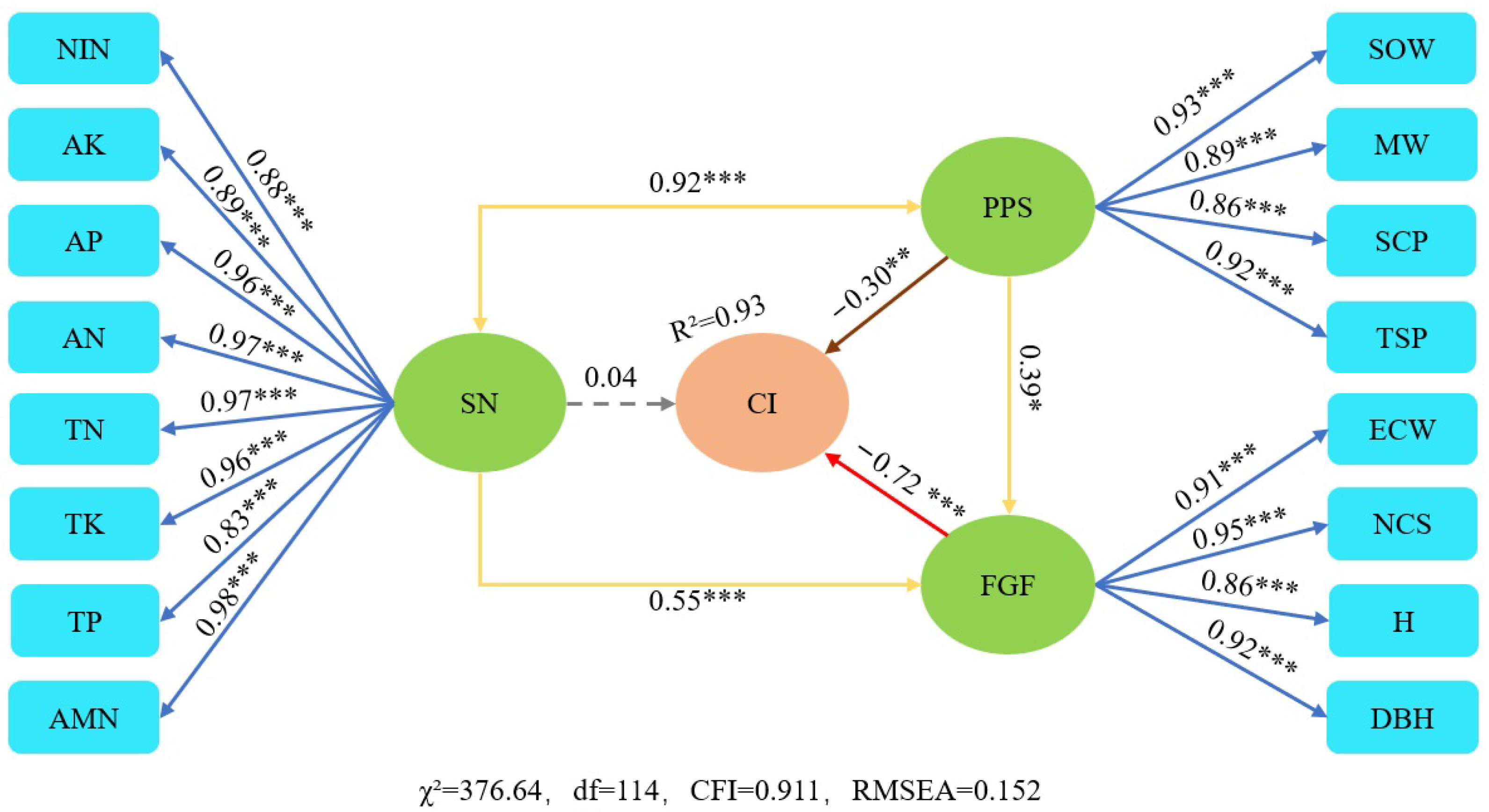
3.4. Structural Equation Model
4. Discussion
4.1. Relationship between Growth Factors of Dominant Trees and Competition Index
4.2. Relationship between Soil Physical and Chemical Properties and Competition Index
4.3. SEM Analysis
4.4. Future Research Directions and Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cortini, F.; Comeau, P.G. Evaluation of competitive effects of green alder, willow and other tall shrubs on white spruce and lodgepole pine in Northern Alberta. For. Ecol. Manag. 2008, 255, 82–91. [Google Scholar] [CrossRef]
- Chen, L.-X.; Ha, X.-M.; Duan, W.-B.; Han, D.H.; Li, C.Z.; Liu, Y.P.; Shen, H.L.; Zhang, P. Analysis on influencing factors of competitive index of dominant trees in Pinus koraiensis plantation. Acta Ecol. Sin. 2022, 42, 1777–1787. [Google Scholar] [CrossRef]
- Liu, W.S.; Li, X.; Chen, F.Y.; Zhu, M.T.; Mu, L.Q. Intraspecific and Interspecific Competition of Quercus mongolica Forest. Bull. Bot. Res. 2020, 40, 552–558. [Google Scholar]
- Dou, X.W.; Tang, M.P. Gravitational model-based competitive analysis of trees. Chin. J. Appl. Ecol. 2022, 33, 2695–2704. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Fraver, S.; Kastendick, D.N. Size-growth relationship, tree spatial patterns, and tree-tree competition influence tree growth and stand complexity in a 160-year red pine chronosequence. For. Ecol. Manag. 2018, 424, 85–94. [Google Scholar] [CrossRef]
- Yuan, X.Z.; Chen, Z.L.; Liu, H. Competitions between wetland dominant plants in water-level fluctuation zone. Chin. J. Ecol. 2011, 30, 1863–1867. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, H.; Li, Y.L.; Qiu, S. Tree competition index based on the structural equation model. J. Beijing For. Univ. 2015, 37, 28–37. [Google Scholar]
- Avery, T.E.; Burkhart, H.E. Forest Measurement, 3rd ed.; McGraw-Hill: New York, NY, USA, 1983. [Google Scholar]
- Du, X.F.; Tang, M.P.; Pan, J.Y.; Shen, Q.Y.; Yang, F. Study on competition index of different typical forest types on Lin′an region. Acta Ecol. Sin. 2020, 40, 4064–4072. [Google Scholar]
- Hegyi, F. A simulation model for managing jack pine stands. In Growth Models for Tree and Stand Simulation; Fries, J., Ed.; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Jiang, X.D. Research on the development and optimization of single wood models. Cent. South For. Inventory Plan. 1996, 15, 1–4. [Google Scholar]
- Shen, C.C.; Lei, X.D.; Wang, F.Y.; Ma, W.; Shen, J.B. Competitive States in Natural Middle-aged Forest of Mongolian Oak at Jincang Forest Farm. For. Res. 2012, 25, 339–345. [Google Scholar] [CrossRef]
- Li, S.H.; Bai, X.R. A summary of structural equation model. Coast. Enterp. Sci. Technol. 2009, 12, 40–42. [Google Scholar]
- Dai, L.; Ge, J.; Wang, L.; Zhang, Q.; Liang, T.; Bolan, N.; Lischeid, G.; Rinklebe, J. Influence of soil properties, topography, and land cover on soil organic carbon and total nitrogen concentration: A case study in Qinghai-Tibet plateau based on random forest regression and structural equation modeling. Sci. Total Environ. 2022, 821, 153440. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.M.; Zheng, J.W.; Wang, D.Z.; Zhang, Z.D. Structural equation modeling analysis of theresponse of herbaceous species richness to landscape factors in a forest-steppe zone. Acta Ecol. Sin. 2018, 38, 4649–4656. [Google Scholar] [CrossRef]
- Liu, D. Research on the Impact Factor Analysis of Forest Ecosystem Service Function Based on the Structural Equation Modeling. Master’s Thesis, Beijing Forestry University, Beijing, China, 2012. [Google Scholar]
- Sanaei, A.; Ali, A.; Yuan, Z.; Liu, S.; Lin, F.; Fang, S.; Ye, J.; Hao, Z.; Loreau, M.; Bai, E.; et al. Context-dependency of tree species diversity, trait composition and stand structural attributes regulate temperate forest multifunctionality. Sci. Total Environ. 2020, 757, 43724. [Google Scholar] [CrossRef]
- Zheng, C.Y.; Xu, Z.Q.; Ma, C.M. The Factors Influencing the Poplar Shelterbelt Degradation in the Bashang Plateau of Northwest Hebei Province. For. Resour. Manag. 2018, 47, 9–15. [Google Scholar] [CrossRef]
- Du, H.F. Reasons for the degradation of Poplar trees in Hebei dam and its reconstruction technology. Environ. Dev. 2019, 31, 195+197. [Google Scholar] [CrossRef]
- Radtke, P.J.; Burkhart, H.W. A comparison of methods for edgebias compensation. Can. J. For. Res. 1998, 28, 942–945. [Google Scholar] [CrossRef]
- Hrinca, I. An optimal control problem for the lotka-volterra system with delay. Nonlinear Anal. Theory Methods Appl. 1997, 28, 247–262. [Google Scholar] [CrossRef]
- Tang, M.P.; Chen, Y.G.; Shi, Y.J.; Zhou, G.M.; Zhao, M.S. Intraspecific and Interspecific competition analysis of community dominant plant populations based on Voronoi diagram. Acta Ecol. Sin. 2007, 27, 4707–4716. [Google Scholar]
- Getzin, S.; Wiegand, K.; Wiegand, T.; Yizhaq, H.; Von Hardenberg, J.; Meron, E. Adopting a spatially explicit perspective to study the mysterious fairy circles of Namibia. Ecography 2015, 38, 1–11. [Google Scholar] [CrossRef]
- Zheng, B.Z. Technical Guidelines for Soil Analysis; China Agriculture Press: Beijing, China, 2013; ISBN 7-109-17317-8. [Google Scholar]
- Shi, Y.F.; Shi, S.H.; Huang, X.M. The application of structural equation modeling in ecology based on R. Chin. J. Ecol. 2022, 41, 1015–1023. [Google Scholar] [CrossRef]
- Wang, M.Y. Effects of Interspecific Competition of Tree Species on Plant and Soil Carbon and Nitrogen Contents in Subtropical forests. Ph.D. Thesis, East China Normal University, Shanghai, China, 2021. [Google Scholar] [CrossRef]
- Mai Mu Ti Ming, Z.L.B. Study on Modeling of Competition Indices of Pinus tabulaeformis Individual Trees at Badaling Area. Master’s Thesis, Beijing Forestry University, Beijing, China, 2012. [Google Scholar]
- Yin, D.S.; Ge, W.Z.; Zhang, F.H.; Shen, H.L. Competition relationship of populations of natural secondary Acer mono forest. Bull. Bot. Res. 2012, 32, 105–109. [Google Scholar]
- Zhang, Q.; Hong, W.; Wu, C.Z.; Hu, X.S.; Lin, Y.M. Studies on intraspecific and interspecific competition in natural communities of Tsuga longibracteata. Guihaia 2005, 25, 14–17. [Google Scholar]
- Zhang, L.J.; Hu, Y.B.; Zhao, Z.H.; Sun, C.Z. Spatial structure diversity of platycladus orientalis plantation in Beijing Jiulong Mountain. Chin. J. Ecol. 2015, 34, 60–69. [Google Scholar] [CrossRef]
- Li, B.; Chen, J.K.; Andrew, R.W. A Literature Review on Plant Competition. Chin. Bull. Bot. 1998, 15, 18–29. [Google Scholar]
- Li, C. Screening of Competition Index for Pinus tabulaeformis Plantations and the Impact of Factors on the Diversity of Understory Species. Master’s Thesis, Chinese Academy of Forestry, Beijing, China, 2019. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Liu, P.; Wei, Z.P. The Relationship between DBH Growth and Competition Factors for Pinus tabulaeformis Plantation. J. Shenyang Agric. Univ. 2016, 47, 299–306. [Google Scholar]
- Wang, Q.; Pan, P.; OuYang, X.Z. Intraspecific and interspecific competition intensity in mixed plantation with different proportion of Pinus massoniana and Schima superba. Chin. J. Ecol. 2021, 40, 49–57. [Google Scholar] [CrossRef]
- Chai, Z.Z.; Wang, D.X.; Zhang, L.N.; Zhang, Y.; Huang, Q.P.; Wu, H. Intraspecific and interspecific competition of Pinus tablaeformis natural secondary forests in western section of Qinling Mountains. J. Northwest AF Univ. 2012, 40, 57–63. [Google Scholar] [CrossRef]
- Tu, H.R.; Li, J.F.; Liu, R.H.; Liang, S.C.; Lan, Z.N.; Zhang, X.Y.; Kang, X.D.; Jiang, Y. Spatial distribution patterns and association of Loropetalum chinense population in karst hills of Guilin, Southwest China. Chin. J. Appl. Ecol. 2019, 30, 2621–2630. [Google Scholar] [CrossRef]
- Li, H.D.; Shen, W.S.; Fang, Y.; Yan, S.G.; Zhang, H.; Zhao, W. Point pattern analysis of several psammophyte populations in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet, China. Chin. J. Plant Ecol. 2011, 35, 834–843. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Review: Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil Sci. 2002, 165, 296–382. [Google Scholar] [CrossRef]
- Yang, X. Coupling Relationships between Stand Structure and Soil and Water Conservation Functionin Based on Structural Equation Model. Master’s Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar] [CrossRef]
- Noorbakhsh, S.; Schoenau, J.; Si, B.C.; Zeleke, T. Soil Properties, Yield, and Landscape Relationships in South-Central Saskatchewan Canada. J. Plant Nutr. 2008, 31, 539–556. [Google Scholar] [CrossRef]
- Pei, Z.X.; Wu, Y.; Wang, Q.; Zhong, Z.L.; Ren, M.L.; Wei, C.H.; Lu, J.L.; Wang, W.J. Correlations between Soil Porosity-Related Parameters and Other Soil Parameters in Songnen Plain, Northeastern China. Res. Soil Water Conserv. 2016, 23, 134–138. [Google Scholar] [CrossRef]
- Deng, J.F.; Ding, G.D.; Wei, Y.W.; Zhao, G.P.; Zhang, R.H.; Deng, G.; Zhou, Y.B.; Yin, Y. Soil physiochemical properties and the relations with Mongolian pine growth in Southern Mu Us desert. J. Arid. Land Resour. Environ. 2017, 31, 160–166. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Liu, L.X.; Han, L.N.; Wang, D.X. Coupling of different plant functional group, soil, and litter nutrients in a natural secondary mixed forest in the Qinling Mountains, China. Environ. Sci. Pollut. Res. 2021, 28, 66272–66286. [Google Scholar] [CrossRef]
- Long, J.; Tang, M. The relationship between spatial structure and topographic factors in the evergreen broad-leaved forest of Tianmushan. J. Zhejiang AF Univ. 2021, 38, 47–57. [Google Scholar]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. The quest for a mechanistic understanding of resource competition in plant communities: The role of experiments. Funct. Ecol. 1991, 5, 241–253. [Google Scholar] [CrossRef]
- Bush, J.K.; Van Auken, O.W. Competition between Schizachyrium scoparium and Buchloe dactyloides: The role of soil nutrients. J. Arid. Environ. 2010, 74, 49–53. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Average Value |
|---|---|
| Average DBH (cm) | 5.45 |
| Average height (cm) | 277.65 |
| East-West Crown width (cm) | 177.26 |
| North-South Crown width (cm) | 170.33 |
| Causality of Variables | Direct Effect | Indirect Effect | Total Effect |
|---|---|---|---|
| Ci←SN | 0.04 | −0.70 | −0.66 |
| Ci←PPS | −0.30 | −0.61 | −0.91 |
| Ci←FGF | −0.72 | 0 | −0.72 |
| PPS←SN | 0.92 | 0 | 0.92 |
| SN←PPS | 0.92 | 0 | 0.92 |
| FGF←SN | 0.55 | 0.36 | 0.91 |
| FGF←PPS | 0.39 | 0.51 | 0.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Jia, Z.; Li, Q.; He, L.; Gao, Y.; Wang, L.; Han, D. A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia. Forests 2023, 14, 1191. https://doi.org/10.3390/f14061191
Zhang J, Jia Z, Li Q, He L, Gao Y, Wang L, Han D. A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia. Forests. 2023; 14(6):1191. https://doi.org/10.3390/f14061191
Chicago/Turabian StyleZhang, Jiapeng, Zhiqing Jia, Qingxue Li, Lingxianzi He, Ya Gao, Long Wang, and Dong Han. 2023. "A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia" Forests 14, no. 6: 1191. https://doi.org/10.3390/f14061191
APA StyleZhang, J., Jia, Z., Li, Q., He, L., Gao, Y., Wang, L., & Han, D. (2023). A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia. Forests, 14(6), 1191. https://doi.org/10.3390/f14061191