
Cinnamyl Alcohol Dehydrogenase Gene Regulates Bursaphelenchus xylophilus Reproduction and Development

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation and Culture of B. xylophilus
2.3. Construction of cDNA Library of B. xylophilus
2.4. Cloning of BxCAD-1 Gene
2.5. Biological Information Analysis of BxCAD-1 Gene
2.6. Construction of Engineered Bacteria
2.7. Expression and Purification of Recombinant BxCAD-1
2.8. Determination of Physical and Chemical Properties of BxCAD-1
2.9. RNA Interference for BxCAD-1 Gene
2.9.1. Synthesis of dsRNA
2.9.2. Efficiency Assessment of RNAi
2.9.3. Effect of RNAi on Movement
2.9.4. Effect of RNAi on Feeding and Reproduction
2.9.5. Effect of RNAi on Spawning Rate and Hatching Rate
2.9.6. Effect of RNAi on Lifespan of PWN
2.9.7. Effect of RNAi on Infectivity
2.9.8. Effect of RNAi on Development
2.9.9. Effect of RNAi on Sensitivity to Ethanol
2.9.10. Effect of RNAi on the Reactive Oxygen Species and Activities of Antioxidant Enzymes
2.10. Fluorescence In Situ Hybridization of BxCAD-1 Gene
2.11. Analysis of KEGG Pathway of BxCAD-1 Gene
2.12. Data Analysis
3. Results
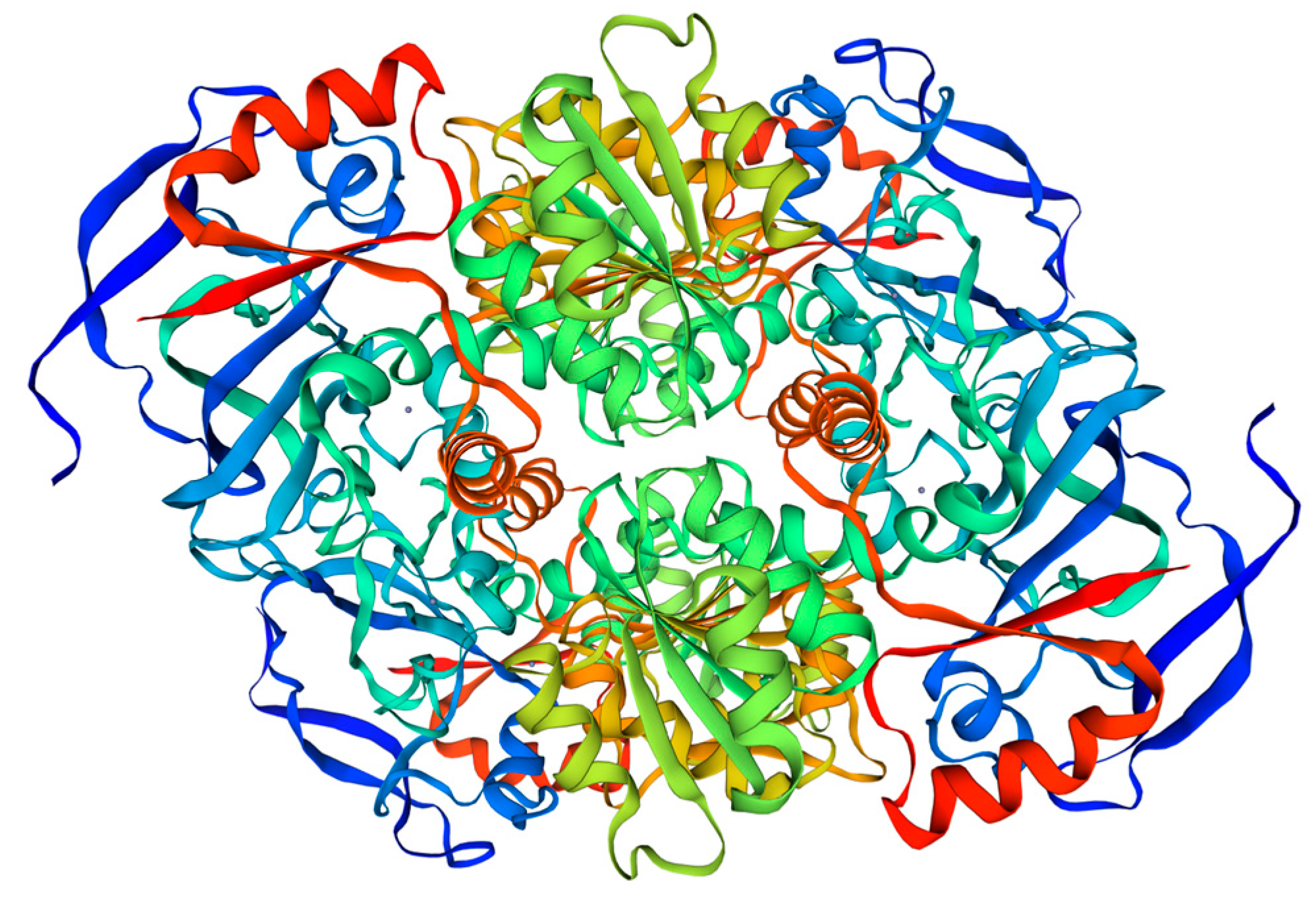
3.1. Sequence Analysis of BxCAD-1 Gene
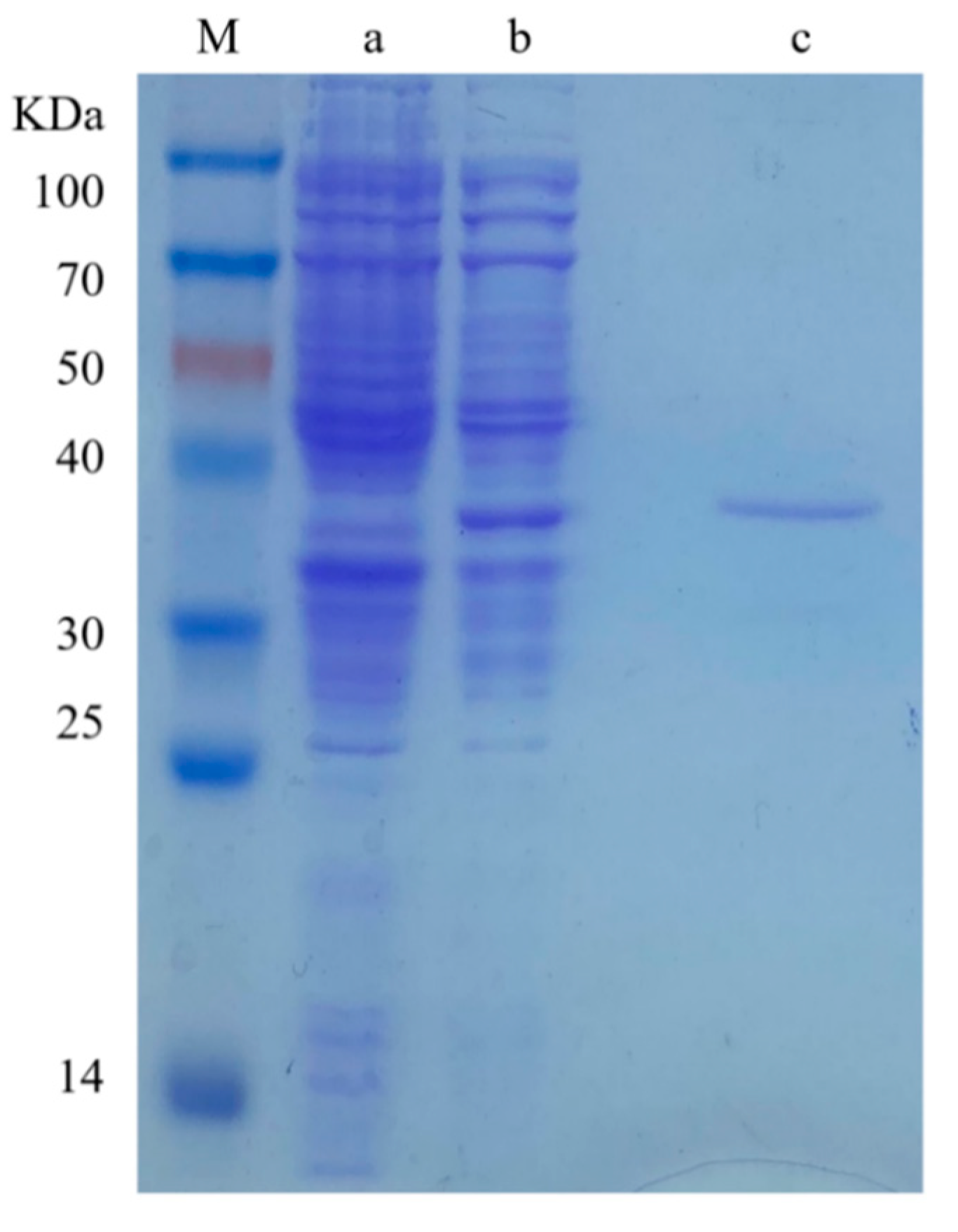
3.2. Induced Expression and Purification of Recombinant BxCAD-1
3.3. Determination of Cinnamyl Alcohol Dehydrogenase Activity
3.4. RNAi of BxCAD-1
3.4.1. Efficiency Assessment of RNAi
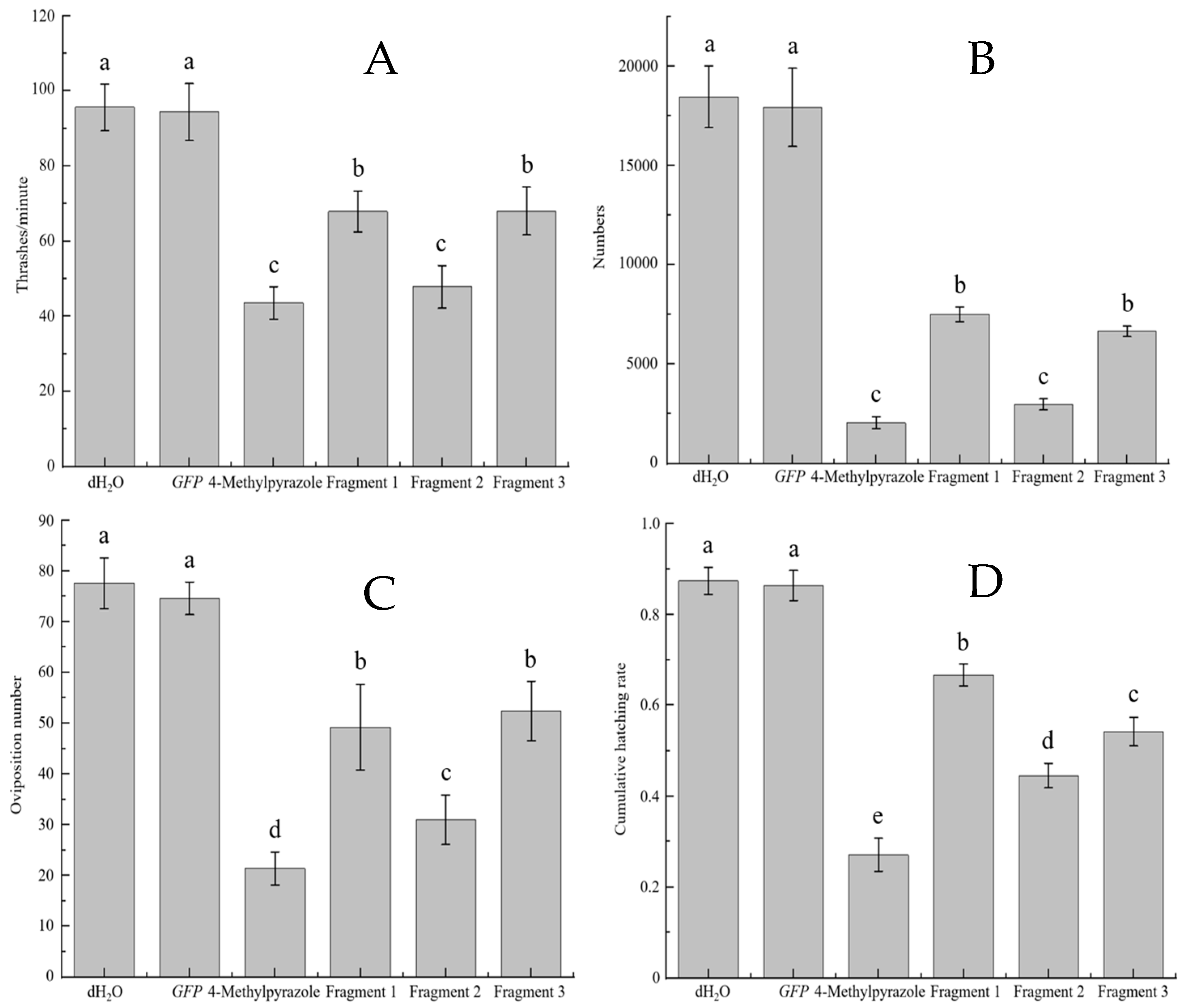
3.4.2. B. xylophilus Movement and Reproduction
3.4.3. Spawning and Hatching Rate of B. xylophilus
3.4.4. Feeding and Infectivity Ability of B. xylophilus
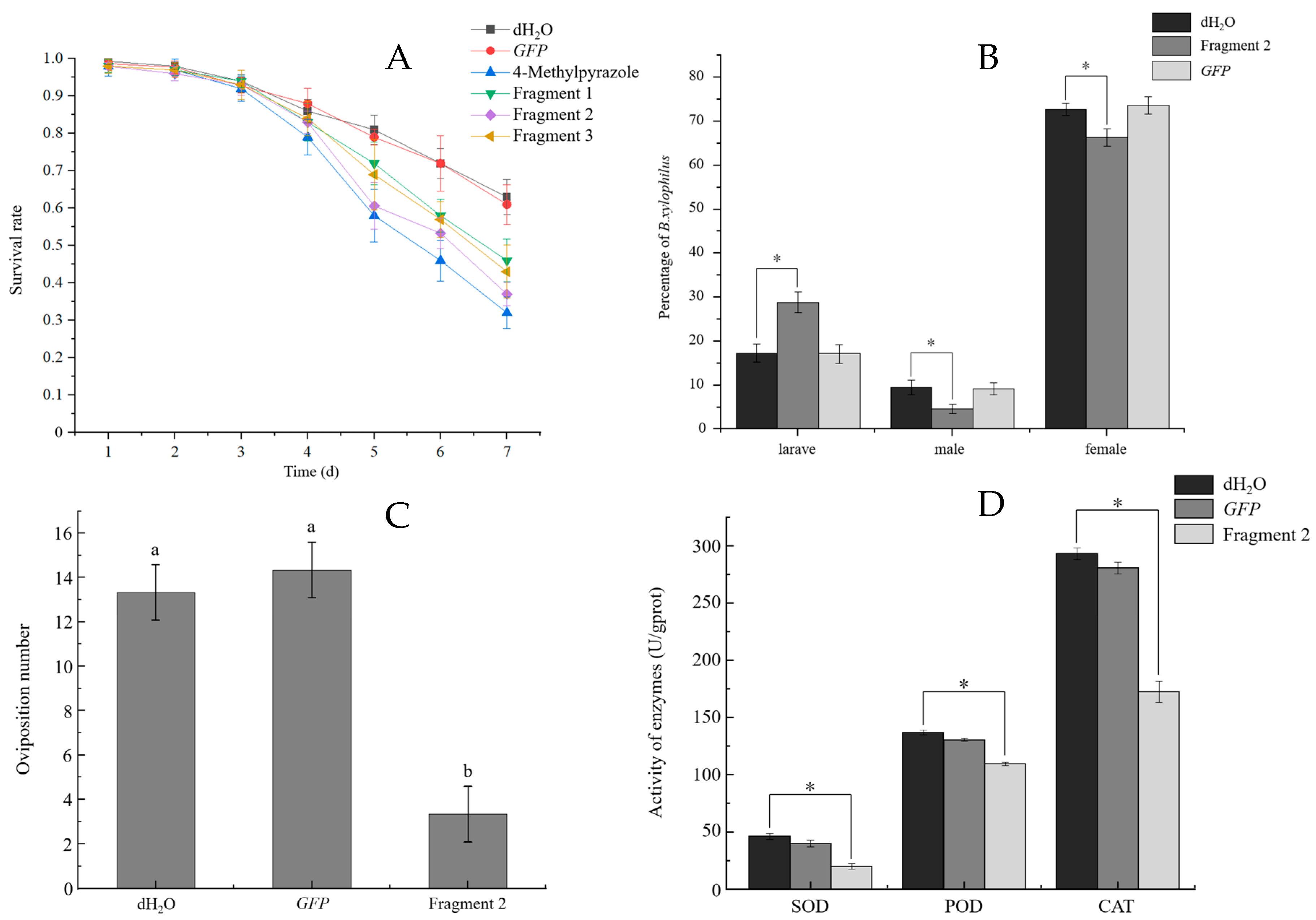
3.4.5. Lifespan and Development of B. xylophilus
3.4.6. Sensitivity to Ethanol and Oviposition of B. xylophilus
3.4.7. Reactive Oxygen Species and Activities of Antioxidant Enzymes
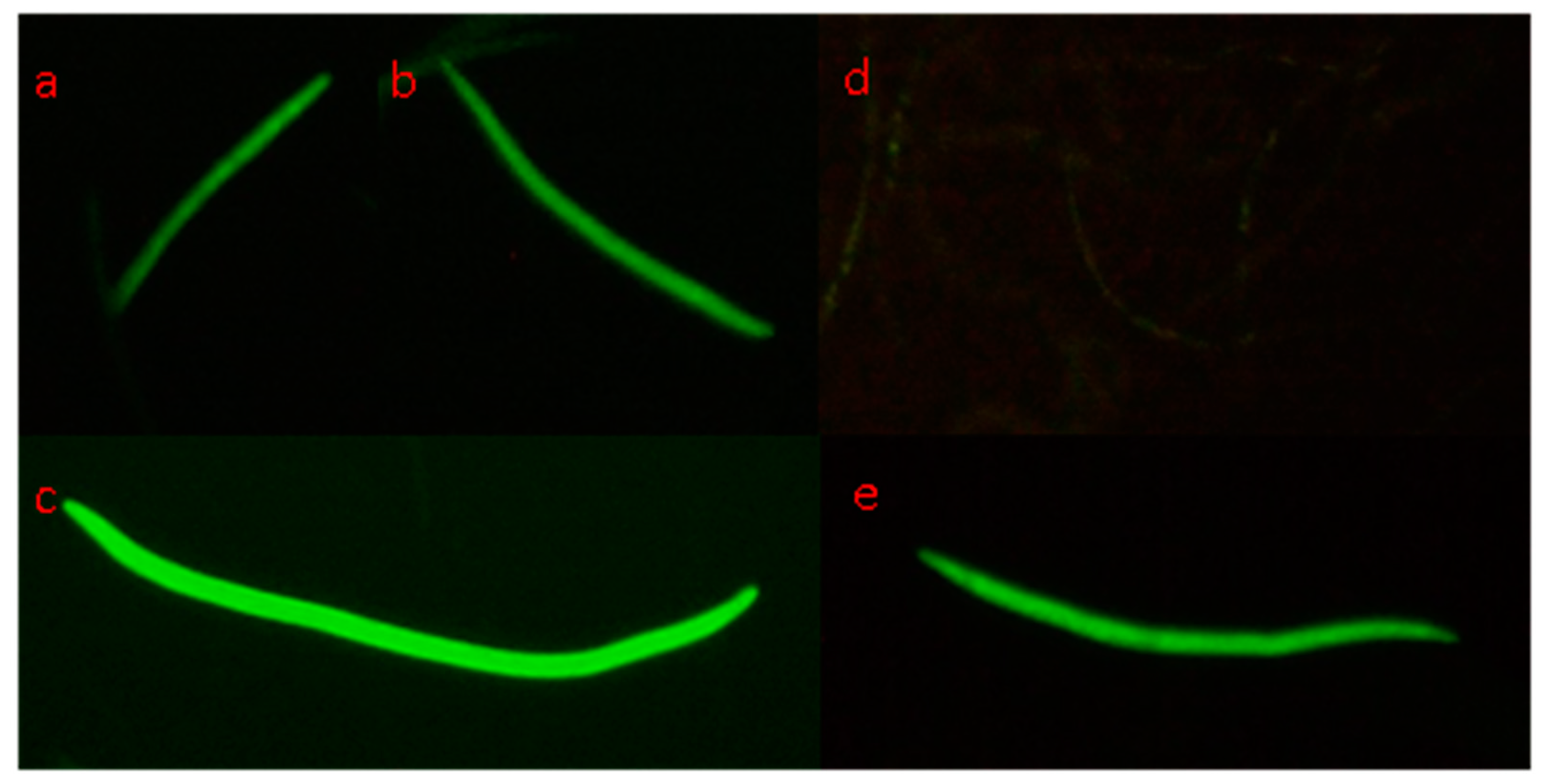
3.5. In Situ Hybridization
3.6. Analysis of KEGG Pathway of BxCAD-1 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. (Eds.) Pine Wilt Disease; Springer International Publishers: Tokyo, Japan, 2008; pp. 5–13. [Google Scholar]
- Wang, Y.; Chen, F.M.; Wang, L.C.; Zhou, L.F.; Song, J. Study of the departure of pine wood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) from Monochamus alternatus (Coleoptera: Cerambycidae). J. Asia-Pac. Entomol. 2020, 23, 981–987. [Google Scholar] [CrossRef]
- Fukuda, K.; Hogetsu, T.; Suzuki, K. Cavitation and cytological changes in xylem of pine seedlings inoculated with virulent and avirulent isolates of Bursaphelenchus xylophilus and B. mucronatus. JJFS 1992, 74, 289–299. [Google Scholar]
- Ishida, K.; Hogetsu, T. Role of resin canals in the early stages of pine wilt disease caused by the pine wood nematode. Can. J. Bot. 1997, 75, 346–351. [Google Scholar] [CrossRef]
- Hara, N.; Futai, K. Histological changes in xylem parenchyma cells and the effects on tracheids of Japanese black pine inoculated with pine wood nematode, Bursaphelenchus xylophilus. J. Jpn. For. Soc. 2001, 83, 285–289. [Google Scholar]
- Ikegami, M.; Jenkins, T.A.R. Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model—Pine Wilt disease as a model case. For. Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- Yang, B.J.; He, Z.Y.; Wang, C.F. Survey of pine wood nematode disease abroad. For. Pest Dis. 1999, 5, 40–42. [Google Scholar]
- Mantas, V.; Fonseca, L.; Baltazar, E.; Canhoto, J.; Abrantes, I. Detection of tree decline (Pinus pinaster aiton) in european forests using sentinel-2 data. Remote. Sens. 2022, 14, 2028. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Wu, W.B.; Zhang, Z.B.; Zheng, L.J.; Han, C.Y.; Wang, X.M.; Xu, J.; Wang, X.R. Research progress on the early monitoring of pine wilt disease using hyperspectral techniques. Sensors 2020, 20, 3729. [Google Scholar] [CrossRef]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, H.Y.; Bai, G.; Xiong, H.L.; Yang, W.B. Immunological Analysis of Bursaphelenchus xylophilus Pathogenous Cellulase. Sci. Silvae Sin. 2007, 43, 64–67. [Google Scholar]
- Kikuchi, T.; Jonesb, J.T.; Aikawa, T.; Kosaka, H.; Ogura, N. A family of glycosyl hydrolase family 45 cellulases from the pine wood nematode Bursaphelenchus xylophilus. FEBS Lett. 2004, 572, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, J.M.S.; Fonseca, L.; Abrantes, I. α-l-fucosidases from Bursaphelenchus xylophilus secretome—Molecular characterization and their possible role in breaking down plant cell walls. Forests 2020, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Shinya, R.; Kirino, H.; Morisaka, H.; Takeuchi-Kaneko, Y.; Futai, K.; Ueda, M. Comparative secretome and functional analyses reveal glycoside hydrolase family 30 and cysteine peptidase as virulence determinants in the pinewood nematode Bursaphelenchus xylophilus. Front. Plant Sci. 2021, 12, 640459. [Google Scholar] [CrossRef]
- Shinya, R.; Morisaka, H.; Kikuchi, T.; Takeuchi, Y.; Ueda, M.; Futai, K. Secretome analysis of the pine wood nematode Bursaphelenchus xylophilus reveals the tangled roots of parasitism and its potential for molecular mimicry. PLoS ONE 2013, 8, e67377. [Google Scholar] [CrossRef]
- Yazaki, K.; Takanashi, T.; Kanzaki, N.; Komatsu, M.; Levia, D.F.; Kabeya, D.; Tobita, H.; Kitao, M.; Ishida, A. Pine wilt disease causes cavitation around the resin canals and irrecoverable xylem conduit dysfunction. J. Exp. Bot. 2018, 69, 589–602. [Google Scholar] [CrossRef]
- Li, Y.X.; Ying, W.; Liu, Z.Y.; Wang, X.; Lu, Q.; Jia, X.Z.; Zhang, X.Y. Functional analysis of the venom allergen-like protein gene from pine wood nematode Bursaphelenchus xylophilus using a baculovirus expression system. Physiol. Mol. Plant. Pathol. 2015, 93, 58–66. [Google Scholar] [CrossRef]
- Hara, N.; Takeuchi, Y.; Futai, K. Cytological changes in ray parenchyma cells of seedlings of three pine species infected with the pine wilt disease. J. Nematol. 2006, 36, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Umebayashi, T.; Fukuda, K.; Haishi, T.; Sotooka, R.; Zuhair, S.; Otsuki, K. The developmental process of xylem embolisms in pine wilt disease monitored by multipoint imaging using compact magnetic resonance imaging. Plant Physiol. 2011, 156, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.G.; Li, L.; Ju, Y.W.; Zhang, J.C.; Zhao, B.G. Pathological changes in ultrastructure of the Pinus thunbergii due to flagellin toxin. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2018, 42, 137–144. [Google Scholar]
- Zhu, L.H.; Ye, J.R.; Negi, S.; Xu, X.L.; Wang, Z.L.; Ji, J.Y. Pathogenicity of aseptic Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e38095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Q.; Xiang, Y.; Wu, X.Q.; Li, M.J. Bacterial communities and virulence associated with pine wood nematode Bursaphelenchus xylophilus from different Pinus spp. Int. J. Mol. Sci. 2019, 20, 3342. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.M.; Hong, Y.D.; Zhao, B.G. A study on pathogenicity of bacteria carried by pine wood nematodes. J. Phytopathol. 2010, 151, 683–689. [Google Scholar] [CrossRef]
- Ebel, J.; Grisebach, H. Reduction of cinnamic acids to cinnamyl alcohols with an enzyme preparation from cell suspension cultures of soybean (Glycine max). FEBS Lett. 1973, 30, 141–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, R.; Zhang, X.Q.; Li, Y.H.; Xie, X.M. Cloning and in silico analysis of a cinnamyl alcohol dehydrogenase gene in Pennisetum purpureum. J. Genet. 2014, 93, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Kasirajan, L.; Valiyaparambth, R.; Kubandiran, A.; Velu, J. Isolation, cloning and expression analysis of cinnamyl alcohol dehydrogenase (CAD) involved in phenylpropanoid pathway of Erianthus arundinaceus, a wild relative of sugarcane. 3 Biotech 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, D.M.; Chen, J.L.; Pu, G.B.; Ji, Y.P.; Lei, C.Y.; Du, Z.G.; Liu, B.Y.; Ye, H.C.; Wang, H. Biochemical characterization and identification of a cinnamyl alcohol dehydrogenase from Artemisia annua. Plant. Sci. 2012, 193–194, 85–95. [Google Scholar] [CrossRef]
- Park, H.L.; Kim, T.L.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Biochemical characterization of the rice cinnamyl alcohol dehydrogenase gene family. Molecules 2018, 23, 2659. [Google Scholar] [CrossRef] [Green Version]
- Sibout, R.; Eudes, A.; Mouille, G.; Pollet, B.; Lapierre, C.; Jouanin, L.; Séguin, A. Cinnamyl alcohol dehydrogenase-C and -D are the primary genes involved in lignin biosynthesis in the floral stem of Arabidopsis. Plant Cell 2005, 17, 2059–2076. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chang, J.M.; Wang, S.; Lu, J.; Liu, Y.; Si, H.Q.; Sun, G.; Ma, C.X. Cloning, expression analysis and molecular marker development of cinnamyl alcohol dehydrogenase gene in common wheat. Protoplasma 2021, 17, 881–889. [Google Scholar] [CrossRef]
- Chao, N.; Liu, S.X.; Liu, B.M.; Li, N.; Jiang, X.N.; Gai, Y. Molecular cloning and functional analysis of nine cinnamyl alcohol dehydrogenase family members in Populus tomentosa. Planta 2014, 240, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Larroy, C.; Pare´s, X.; Biosca, J.A. Characterization of a Saccharomyces cerevisiae NADP(H)-dependent alcohol dehydrogenase (ADHVⅡ), a member of the cinnamyl alcohol dehydrogenase family. Eur. J. Biochem. 2002, 269, 5738–5745. [Google Scholar] [CrossRef] [PubMed]
- Larroy, C.; Fernández, M.R.; González, E.; Parés, X.; Biosca, J.A. Properties and functional significance of Saccharomyces cerevisiae ADHVI. Chem. Biol. Interact. 2003, 143, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.H.; Lin, Y.; Fu, Y.P.; Xie, J.T.; Jiang, D.H.; Cheng, J.S. A cinnamyl alcohol dehydrogenase required for sclerotial development in Sclerotinia sclerotiorum. Phytopathol. Res. 2020, 2, 13. [Google Scholar] [CrossRef]
- Wilkin, J.M.; Soetaert, K.; Ste´landre, M.; Buyssens, P.; Castillo, G.; Demoulin, V.; Bottu, G.; Laneelle, M.A.; Daffe, M.; Bruyn, J.D. Overexpression, purification and characterization of Mycobacterium bovis BCG alcohol dehydrogenase. Eur. J. Biochem. 1999, 262, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Mee, B.; Kelleher, D.; Frias, J.; Malone, R.; Tipton, K.F.; Henehan, G.T.M.; Windle, H.J. Characterization of cinnamyl alcohol dehydrogenase of Helicobacter pylori. An aldehyde dismutating enzyme. FEBS J. 2010, 272, 1255–1264. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.Q.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Chi-Ham, C.L.; Clark, K.L.; Bennett, A.B. The intellectual property landscape for gene suppression technologies in plants. Nat. Biotechnol. 2010, 28, 32–36. [Google Scholar] [CrossRef]
- Niu, J.H.; Jian, H.; Xu, J.M.; Guo, Y.D.; Liu, Q. RNAi technology extends its reach: Engineering plant resistance against harmful eukaryotes. Afr. J. Biotechnol. 2015, 9, 7573–7582. [Google Scholar]
- Wang, M.; Wang, D.; Zhang, X.; Wang, X.; Liu, W.; Hou, X.; Huang, X.Y.; Xie, B.Y.; Cheng, X.Y. Double-stranded RNA-mediated interference of dumpy genes in Bursaphelenchus xylophilus by feeding on filamentous fungal transformants. Int. J. Parasitol. 2016, 46, 351–360. [Google Scholar] [CrossRef]
- Xue, Q.; Wu, X.Q.; Zhang, W.J.; Deng, L.N.; Wu, M.M. Cathepsin L-like cysteine proteinase genes are associated with the development and pathogenicity of pine wood nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.; Yang, L.; Ye, J.; Wang, W.; Zhao, T.; Hu, H.; Zhou, G. Silencing of cyp-33C9 gene affects the reproduction and pathogenicity of the pine wood nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 4520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Ye, J.R. Influence of Bxpel1 gene silencing by dsRNA interference on the development and pathogenicity of the pine wood nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2016, 17, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.J.; Wu, X.Q.; Li, H.Y.; Zhao, Q.; Wang, Y.C.; Ye, J.R. An effector, BxSapB1, induces cell death and contributes to virulence in the pine wood nematode Bursaphelenchus xylophilus. Mol. Plant Microbe Interact. 2019, 32, 452–463. [Google Scholar]
- Di, L.; Balesano, A.; Jordan, S.; Shi, S.M. The role of alcohol dehydrogenase in drug metabolism: Beyond ethanol oxidation. AAPS J. 2021, 23, 20. [Google Scholar] [CrossRef]
- Zhao, T.Y.; Han, Z.X.Y.; Zhang, J.M.; Ding, Y.; Chen, J.J.; Qiao, H.L.; Gao, N. Effect of ADHI on hepatic stellate cell activation and liver fibrosis in mice. Biochem. Biophys. Res. Commun. 2023, 651, 98–106. [Google Scholar]
- Villalobos-García, D.; Ayhllon-Osorio, C.A.; Hernández-Muñoz, R. The fructose-dependent acceleration of ethanol metabolism. Biochem. Pharmacol. 2021, 188, 114498. [Google Scholar] [CrossRef]
- Wang, L.S. Function Analysis of adh-1 of B. xylophilus. Ph.D. Thesis, Qingdao University, Qingdao, China, 2019. [Google Scholar]
- Guo, Q.Q.; Du, G.C.; Qi, H.T.; Zhang, Y.N.; Yue, T.Q.; Wang, J.C.; Li, R.G. A nematicidal tannin from Punica granatum L. rind and its physiological effect on pine wood nematode (Bursaphelenchus xylophilus). Pestic. Biochem. Physiol. 2017, 135, 64–68. [Google Scholar] [CrossRef]
- Wang, M.; Du, G.C.; Fang, J.N.; Wang, L.S.; Guo, Q.Q.; Zhang, T.T.; Li, R.G. UGT440A1 is associated with motility, reproduction, and pathogenicity of the plant-parasitic nematode Bursaphelenchus xylophilus. Front. Plant Sci. 2022, 13, 862594. [Google Scholar]
- Liu, G.H.; Feng, K.; Guo, D.S.; Li, R.G. Overexpression and activities of 1-Cys peroxiredoxin from Pseudomonas fluorescens GcM5-1A carried by pine wood nematode. Folia Microbiol. 2015, 60, 443–450. [Google Scholar]
- Wang, L.S.; Zhang, T.T.; Pan, Z.S.; Lin, L.L.; Dong, G.Q.; Wang, M.; Li, R.G. The alcohol dehydrogenase with a broad range of substrate specificity regulates vitality and reproduction of the plant-parasitic nematode Bursaphelenchus xylophilus. Parasitology 2018, 146, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Ma, R.Q.; Zhu, N.; Guo, K.; Guo, Y.; Bai, L.; Yu, H.; Hu, J.; Zhang, X. Bxy-fuca encoding alpha-L-fucosidase plays crucial roles in development and reproduction of the pathogenic pinewood nematode, Bursaphelenchus xylophilus. Pest. Manag. Sci. 2020, 76, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ma, L.; Wang, F.; Wang, B.; Hao, X.; Xu, J.; Ma, Y. Low temperature extends the lifespan of Bursaphelenchus xylophilus through the cGMP pathway. Int. J. Mol. Sci. 2017, 18, 2320. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.X. Isolation and Identification of Nematicidal Bacteria from Rootzone Soils of Pinus thunbergii and Characterization of Derived Active Components against Bursaphelenchus xylophilus. Master’s Thesis, Qingdao University, Qingdao, China, 2022. [Google Scholar]
- Shinya, R.; Takeuchi, Y.; Futai, K. A technique for separating the developmental stages of the propagative form of the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2009, 11, 305–307. [Google Scholar]
- Li, Z.Z.; Shen, H.J.; Jiang, Q.H.; Ji, B.Z. A study on the activities of endogenous enzymes of protective system in some insects. Acta Entomol. Sin. 1994, 37, 399–403. [Google Scholar]
- Kim, C.; Kim, J.; Kim, S.; Cook, D.E.; Evans, K.S.; Andersen, E.C.; Lee, J. Long-read sequencing reveals intra-species tolerance of substantial structural variations and new subtelomere formation in C. elegans. Genome Res. 2019, 29, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Du, G.; Guo, Q.; Dong, G.; Wang, M.; Zhang, T.; Li, R. Transcriptome sequencing and analysis of genes related to disease resistance in Pinus thunbergii. Forests 2023, 14, 650. [Google Scholar] [CrossRef]
- Wang, X.Y.; Wu, X.Q.; Wen, T.Y.; Feng, Y.Q.; Zhang, Y. Terpene production varies in Pinus thunbergii parl. with different levels of resistance, with potential effects on pinewood nematode behavior. Forests 2022, 13, 1140. [Google Scholar] [CrossRef]
- Hu, L.J.; Wu, X.Q.; Wen, T.Y.; Qiu, Y.J.; Rui, L.; Zhang, Y.; Ye, J.R. A Bursaphelenchus xylophilus effector, BxSCD3, suppresses plant defense and contributes to virulence. Int. J. Mol. Sci. 2022, 23, 6417. [Google Scholar] [CrossRef]
- Li, Y.; Hu, L.J.; Wu, X.Q.; Ye, J.R. A Bursaphelenchus xylophilus effector, Bx-FAR-1, suppresses plant defense and affects nematode infection of pine trees. Eur. J. Plant. Pathol. 2020, 157, 637–650. [Google Scholar] [CrossRef]
- Yan, D.H.; Yang, B.J. The enzymes in the secretions of pine wood nematode (Bursaphelenchus xylophilus). For. Res. 1997, 10, 265–269. [Google Scholar]
- Shinya, R.; Morisaka, H.; Takeuchi, Y.; Ueda, M.; Futai, K. Comparison of the surface coat proteins of the pine wood nematode appeared during host pine infection and in vitro culture by a proteomic approach. Phytopathology 2010, 100, 1289–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Oda, K. The occurrence of attractiveness for Monochamus alternatus Hope (Coleoptera: Cerambycidae) in nematode-infected pine trees. J. Jap. For. Soc. 1980, 62, 432–434. [Google Scholar]
- Shuto, Y.; Watanabe, H. Stimulating effect of ethanol on oviposition of the pine wood nematode, Bursaphelenchus xylophilus. Agr. Biol. Chem. 1988, 52, 2927–2928. [Google Scholar]
- Wang, M.; Wang, L.S.; Fang, J.N.; Du, G.C.; Zhang, T.T.; Li, R.G. Transcriptomic profiling of Bursaphelenchus xylophilus reveals differentially expressed genes in response to ethanol. Mol. Biochem. Parasitol. 2022, 248, 111460. [Google Scholar] [CrossRef]
- Oku, H. Role of phytotoxins in pine wilt diseases. J. Nematol. 1988, 20, 245–251. [Google Scholar]
- Gao, M.G.; Li, Y.X.; Zhang, W.; Wei, P.F.; Wang, X.; Feng, Y.Q.; Zhang, X.Y. Bx-daf-22 contributes to mate attraction in the gonochoristic nematode Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 4316. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Products | Manufacturers |
|---|---|
| Escherichia coli DH5α, BL21 (DE3), vector pET-15b | Invitrogen Co. (Carlsbad, CA, USA) |
| RNA Easy Fast Tissue/Cell Kit | Tiangen Biotech (Beijing, China) |
| PrimeScriptTM II First Strand cDNA Synthesis Kit | Takara Biomedical Technology (Beijing, China) |
| pEASY®-T1 Cloning Kit | TransGen Biotech Co., Ltd. (Beijing, China) |
| T4 DNA ligase, restriction endonuclease NdeI, XhoI | New England Biolabs (Beijing, China) |
| Extraction Mini Kit, HiScript III RT SuperMix for qPCR, ChanQ Universal SYBR qPCR Master Mix | Vazyme Biotech Co., Ltd. (Nanjing, Jiangsu, China) |
| MEGAscriptTM RNAi Kit | Thermo Fisher Scientific (Waltham, MA, USA) |
| Reactive oxygen species Assay Kit | Nanjing Jiancheng Bioengineering Institute (Nanjing, Jiangsu, China) |
| RNA Free paraformaldehyde fixative, FISH in situ hybridization kit | Shanghai Gefan Biotechnology Co., Ltd. (Shanghai, China) |
| Primer | Sequence (5′→3′) |
|---|---|
| CAD-F | ATGGTGGTATATTTCAGACATTTTG |
| CAD-R | TTAGTTCCACATGTCCAGAAC |
| CAD-1-F | ATGGTGGTATATTTCAGACATTTTG |
| CAD-1-R | TACAAGCGGCATATTGTTGG |
| CAD-2-F | TCCGATCAAGAGCCCACC |
| CAD-2-R | CGTAGGCATCAACGAACC |
| CAD-3-F | TATGCCAAAGCTATGGGAATG |
| CAD-3-R | TCAAGGGTTGCATCTCCAC |
| CAD-1T7-F | TAATACGACTCACTATAGGGATGGTGGTATATTTCAGACATTTTG |
| CAD-1T7-R | TAATACGACTCACTATAGGGTACAAGCGGCATATTGTTGG |
| CAD-2T7-F | TAATACGACTCACTATAGGGTCCGATCAAGAGCCCACC |
| CAD-2T7-R | TAATACGACTCACTATAGGGCGTAGGCATCAACGAACC |
| CAD-3T7-F | TAATACGACTCACTATAGGGTATGCCAAAGCTATGGGAATG |
| CAD-3T7-R | TAATACGACTCACTATAGGGTCAAGGGTTGCATCTCCAC |
| GFP-F | TAATACGACTCACTATAGGGCAAAGATGACGGGAACTAC |
| GFP-R | TAATACGACTCACTATAGGGGATAATGGTCTGCTAGTTG |
| Q1R | CAGACATTTTGATATGTCCGACG |
| Q1F | GGCAGACACCTGAATACAAAATC |
| Q2R | GTACTGCAAACGGGGCTATG |
| Q2F | CCTTCAACGCCTTATAAACCG |
| Q3R | CCTACGGCAAGACTGATGTGG |
| Q3F | CTTCAATGTGACCGTGGCATC |
| CTR | CTGCTGAGCGTGAAATCGT |
| CTF | GTTGTAGGTGGTCTCGTGGA |
| Name | Formula | Remarks |
|---|---|---|
| Formula (1) | Ack: absorbance of sample AE: absorbance of control V: total volume of enzyme solution of the sample w: total protein content of sample v: volume of enzyme solution during measurement | |
| Formula (2) | ΔA470: difference in absorbance V: total volume of enzyme solution of the sample w: total protein content of sample v: volume of enzyme solution during measurement t: reaction time | |
| Formula (3) | ΔA240: difference in absorbance V: total volume of enzyme solution of the sample w: total protein content of sample v: volume of enzyme solution during measurement t: reaction time |
| Group | Fluorescence Intensity (OD) |
|---|---|
| distilled H2O | 22,935.33 ± 523.15 |
| GFP | 28,191.33 ± 619.84 |
| fragment 2 | 56,021.67 ± 1262.13 |
| negative control | 1732.00 ± 10.20 |
| positive control | 32,641.33 ± 479.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, G.; Guo, Q.; Deng, W.; Zhang, Y.; Tai, H.; Wang, C.; Du, G.; Li, R. Cinnamyl Alcohol Dehydrogenase Gene Regulates Bursaphelenchus xylophilus Reproduction and Development. Forests 2023, 14, 1379. https://doi.org/10.3390/f14071379
Dong G, Guo Q, Deng W, Zhang Y, Tai H, Wang C, Du G, Li R. Cinnamyl Alcohol Dehydrogenase Gene Regulates Bursaphelenchus xylophilus Reproduction and Development. Forests. 2023; 14(7):1379. https://doi.org/10.3390/f14071379
Chicago/Turabian StyleDong, Guosong, Qunqun Guo, Wenjun Deng, Yu Zhang, Hongzheng Tai, Chao Wang, Guicai Du, and Ronggui Li. 2023. "Cinnamyl Alcohol Dehydrogenase Gene Regulates Bursaphelenchus xylophilus Reproduction and Development" Forests 14, no. 7: 1379. https://doi.org/10.3390/f14071379