
Seasonal Variation and Soil Texture-Related Thinning Effects on Soil Microbial and Enzymatic Properties in a Semi-Arid Pine Forest



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Forestry Treatments
2.2. Microclimate in the Experimental Plots
2.3. Soil Sampling
2.4. Analysis of General Soil Properties
2.5. Soil Respiration
2.6. Microbial Biomass Carbon
2.7. Enzime Activity Measurements
2.8. Statistical Analysis
3. Results
3.1. Soil Characteristics in the Experimental Blocks
3.1.1. Physico-Chemical Properties of Soils
3.1.2. Soil Water-Holding Capacity
3.2. Seasonal Variation of Soil Labile Organic Carbon Fractions and Basal Soil Respiration
3.3. Seasonal Variation of the Soil Enzyme Parameters for Each Treatment
3.4. Principal Component Analysis (PCA) Correlations between Microbial and Enzymatic Soil Properties and Climate
3.5. Relationships between Microbial and Enzymatic Variables and Soil Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 2 | Factor/Interaction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment (T) | Season (S) | T × S | Residuals | ||||||||
| Block | F | df | p-Value 1 | F | df | p-Value 1 | F | df | p-Value 1 | df | |
| WHC (non-transformed) | I | 3.43 | 2 | 0.0455 * | 2.14 | 4 | 0.0999 ns | 0.63 | 8 | 0.7430 ns | 30 |
| II | 6.82 | 2 | 0.0031 ** | 3.81 | 5 | 0.0072 ** | 0.55 | 10 | 0.8445 ns | 36 | |
| III | 44.01 | 2 | <0.0001 *** | 5.52 | 5 | 0.0007 *** | 1.34 | 10 | 0.2474 ns | 36 | |
| SOC (LOG10-transformed) | I | 4.63 | 2 | 0.0177 * | 5.67 | 4 | 0.0016 ** | 0.83 | 8 | 0.5857 ns | 30 |
| II | 3.09 | 2 | 0.0576 ns | 0.81 | 5 | 0.5512 ns | 0.62 | 10 | 0.7905 ns | 36 | |
| III | 58.27 | 2 | <0.0001 *** | 1.56 | 5 | 0.1974 ns | 0.95 | 10 | 0.5007 ns | 36 | |
| WSOC (SQRT-transformed) | I | 3.57 | 2 | 0.0406 * | 20.30 | 4 | <0.0001 *** | 0.63 | 8 | 0.7480 ns | 30 |
| II | 6.01 | 2 | 0.0056 ** | 10.13 | 5 | <0.0001 *** | 1.1 | 10 | 0.3875 ns | 36 | |
| III | 35.90 | 2 | <0.0001 *** | 7.22 | 5 | 0.0001 *** | 2.3 | 10 | 0.0328 * | 36 | |
| BSR (LOG10-transformed) | I | 6.49 | 2 | 0.0046 ** | 7.45 | 4 | 0.0003 *** | 1.20 | 8 | 0.3337 ns | 30 |
| II | 3.01 | 2 | 0.0617 ns | 10.35 | 5 | <0.0001 *** | 1.34 | 10 | 0.2479 ns | 36 | |
| III | 84.86 | 2 | <0.0001 *** | 3.84 | 5 | 0.0068 ** | 1.94 | 10 | 0.0710 ns | 36 | |
| MBC (SQRT-transformed) | I | 4.79 | 2 | 0.0157 * | 25.02 | 4 | <0.0001 *** | 1.21 | 8 | 0.3258 ns | 30 |
| II | 6.62 | 2 | 0.0036 ** | 9.12 | 5 | <0.0001 *** | 0.53 | 10 | 0.8594 ns | 36 | |
| III | 39.09 | 2 | <0.0001 *** | 3.98 | 5 | 0.0056 ** | 1.94 | 10 | 0.0713 ns | 36 | |
| Parameter 2 | Treatment | Spring 1st | Summer 1st | Autumn 1st | Spring 2nd | Summer 2nd | Autumn 2nd | S C.l. 1 | |
|---|---|---|---|---|---|---|---|---|---|
| WHC | Block I | T0 | 32.62 ± 6.01 | 38.41 ± 3.25 | – | 36.64 ± 2.82 | 34.84 ± 1.78 | 34.48 ± 4.89 ab | ns |
| T60 | 33.03 ± 4.11 | 35.16 ± 4.84 | – | 34.87 ± 2.64 | 33.40 ± 0.73 | 36.95 ± 0.96 b | ns | ||
| T100 | 30.07 ± 0.71 | 33.94 ± 3.42 | – | 34.97 ± 1.19 | 32.73 ± 2.68 | 30.45 ± 2.43 a | ns | ||
| T C.l. 1 | ns | ns | – | ns | ns | * | |||
| Block II | T0 | 38.13 ± 5.95 ab b | 27.78 ± 3.82 ab a | 28.13 ± 2.3 ab | 32.11 ± 8.24 ab | 30.96 ± 1.71 ab ab | 37.36 ± 4.37 b | * | |
| T60 | 32.17 ± 6.88 a b | 25.44 ± 0.21 a a | 33.84 ± 2.14 ab | 30.56 ± 3.07 ab | 29.49 ± 8.89 a ab | 33.31 ± 4.34 b | * | ||
| T100 | 40.83 ± 3.82 b b | 31.76 ± 7.99 b a | 35.28 ± 2.98 ab | 35.88 ± 3.09 ab | 37.15 ± 5.21 b ab | 39.01 ± 2.28 b | * | ||
| T C.l. 1 | * | * | ns | ns | * | ns | |||
| Block III | T0 | 13.08 ± 5.23 | 10.63 ± 4.37 a | 16.6 ± 1.97 a | 12.79 ± 4.27 a | 11.92 ± 1.67 a | 13.06 ± 0.86 a | ns | |
| T60 | 26.69 ± 6.49 b | 20.10 ± 2.82 b ab | 28.8 ± 1.98 b ab | 24.34 ± 3.02 b ab | 18.06 ± 3.07 b a | 27.25 ± 2.31 b ab | ** | ||
| T100 | 15.32 ± 2.95 | 18.09 ± 4.02 ab | 19.2 ± 5.17 a | 19.7 ± 2.2 ab | 9.24 ± 2.91 a | 16.5 ± 5.3 a | ns | ||
| T C.l. 1 | ns | *** | *** | *** | *** | *** | |||
| SOC | Block I | T0 | 64.5 ± 14.44 a | 96.68 ± 19.93 b | – | 65.46 ± 6.48 ab | 62.00 ± 12.73 a | 66.76 ± 29.22 ab ab | ** |
| T60 | 60.62 ± 15.05 ab | 82.11 ± 19.3 b | – | 63.58 ± 6.94 ab | 57.7 ± 4.8 a | 82.39 ± 15.63 b ab | ** | ||
| T100 | 45.26 ± 0.96 a | 79.47 ± 20.24 b | – | 62.47 ± 7.99 ab | 49.50 ± 13.35 ab | 49.36 ± 3.13 a ab | *** | ||
| T C.l. 1 | ns | ns | ns | ns | * | ||||
| Block II | T0 | 55.78 ± 15.09 | 59.40 ± 9.89 | 41.47 ± 6.1 | 53.77 ± 13.16 | 44.06 ± 7.52 | 61.19 ± 14.47 | ns | |
| T60 | 45.81 ± 17.99 | 49.14 ± 3.92 | 55.91 ± 13.37 | 60.09 ± 2.18 | 43.96 ± 6.68 | 55.02 ± 16.96 | ns | ||
| T100 | 54.71 ± 5.43 | 75.70 ± 16.59 | 59.87 ± 11.67 | 46.77 ± 14.37 | 57.98 ± 10.82 | 61.54 ± 31.59 | ns | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ns | |||
| Block III | T0 | 12.85 ± 3.97 a | 9.97 ± 2.27 a | 10.09 ± 0.33 a | 12.50 ± 3.43 a | 12.41 ± 1.46 a | 13.56 ± 2.67 a | ns | |
| T60 | 47.48 ± 24.81 b | 33.03 ± 6.94 b | 40.08 ± 6.74 b | 46.34 ± 20.55 b | 24.66 ± 2.4 b | 52.03 ± 15.78 b | ns | ||
| T100 | 21.08 ± 6.26 a | 22.36 ± 2.24 a | 15.8 ± 8.56 a | 19.41 ± 2.98 a | 13.40 ± 4.92 a | 18.77 ± 12.36 a | ns | ||
| T C.l. 1 | *** | *** | *** | *** | *** | *** | |||
| WSOC | Block I | T0 | 223.10 ± 51.14 a | 532.31 ± 108.81 b | – | 167.48 ± 20.66 a | 304.87 ± 97.15 ab | 160.15 ± 93.82 a | *** |
| T60 | 211.35 ± 64.51 a | 465.69 ± 197.45 b | – | 125.13 ± 20.07 a | 239.78 ± 57.8 ab | 228.55 ± 96.5 a | *** | ||
| T100 | 180.38 ± 19.23 a | 374.07 ± 111.7 b | – | 119.29 ± 52.55 a | 232.86 ± 66.97 ab | 100.69 ± 13.06 a | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ||||
| Block II | T0 | 180.71 ± 32.63 ab | 222.78 ± 26.14 a b | 112.93 ± 24.61 a | 136.89 ± 80.27 a | 144.77 ± 39.93 ab | 174.40 ± 51.14 ab | *** | |
| T60 | 255.93 ± 97.78 a | 234.59 ± 65.95 ab a | 157.61 ± 48.98 ab | 101.20 ± 20.67 a | 208.13 ± 59.6 ab | 188.65 ± 37.96 ab | *** | ||
| T100 | 269.83 ± 12.28 bc | 361.18 ± 72.92c c | 155.47 ± 30.51 ab | 147.06 ± 25.58 a | 246.20 ± 54.67 abc | 163.72 ± 50.04 ab | *** | ||
| T C.l. 1 | ns | * | ns | ns | ns | ns | |||
| Block III | T0 | 74.66 ± 22.83 a | 66.98 ± 19.74 a | 85.57 ± 2.59 a | 142.50 ± 40.3 a | 120.2 ± 33.93 ab | 70.61 ± 11.5 a | ns | |
| T60 | 172.41 ± 93.18 b a | 175.58 ± 57.99 b ab | 163.94 ± 9.71 b a | 254.00 ± 54.53 b b | 151.23 ± 6.71 b ab | 150.59 ± 25.79 b a | ** | ||
| T100 | 51.6 ± 10.23 a a | 160.94 ± 31.93 ab ab | 58.12 ± 20.55 a a | 140.34 ± 45.3 a b | 75.33 ± 39.71 a ab | 69.82 ± 12.82 a a | * | ||
| T C.l. 1 | *** | *** | *** | *** | *** | *** | |||
| BSR | Block I | T0 | 26.48 ± 3.64 ab a | 46.15 ± 9.19 ab b | – | 35.97 ± 3.67 ab ab | 32.75 ± 2.39 ab ab | 31.62 ± 13.06 ab ab | *** |
| T60 | 29.33 ± 5.23 b a | 51.97 ± 19.32 b b | – | 29.97 ± 4.73 b ab | 27.6 ± 6.6 b ab | 41.90 ± 13.19 b ab | *** | ||
| T100 | 24.47 ± 2.71 a a | 37.51 ± 8.13 a b | – | 28.71 ± 0.67 a ab | 23.61 ± 1.56 a a | 21.27 ± 3.03 a ab | *** | ||
| T C.l. 1 | * | ** | * | * | * | ||||
| Block II | T0 | 25.96 ± 5.9 ab | 25.18 ± 1.09 ab | 18.13 ± 1.98 a | 25.14 ± 10.29 ab | 19.36 ± 1.31 a | 39.14 ± 9.2 b | *** | |
| T60 | 23.19 ± 7.79 ab | 24.1 ± 2.09 ab | 27.97 ± 7.72 a | 24.35 ± 3.45 ab | 16.99 ± 2.18 a | 41.07 ± 12.07 b | *** | ||
| T100 | 27.22 ± 0.43 a | 37.71 ± 11.84 ab | 25.44 ± 5.75 a | 31.14 ± 3.08 ab | 20.09 ± 1.69 a | 36.93 ± 4.39 b | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ns | |||
| Block III | T0 | 4.89 ± 1.61 a | 8.15 ± 1.61 a | 7 ± 0.67 a | 8.33 ± 2.87 a | 5.25 ± 0.82 a | 6.38 ± 1.05 a | ns | |
| T60 | 31.34 ± 16.08 b | 22.41 ± 3.11c | 31.2 ± 2.49 b | 24.71 ± 8.87 b | 16.34 ± 3.52 b | 35.73 ± 9.36 b | ns | ||
| T100 | 7.16 ± 2.14 a | 19.64 ± 5.78 b | 10.4 ± 3.99 a | 13.89 ± 6.6 a | 8.28 ± 4.63 a | 8.93 ± 2.93 a | ns | ||
| T C.l. 1 | *** | *** | *** | *** | *** | *** | |||
| MBC | Block I | T0 | 630.28 ± 51.81 a | 434.05 ± 35.38 a | – | 572.92 ± 126.66 a | 398.88 ± 51.05 a | 232.75 ± 43.27 a b | *** |
| T60 | 648.47 ± 219.19 b | 491.88 ± 183.09 ab | – | 768.48 ± 60.94 b | 492.76 ± 43.26 ab | 387.13 ± 31.45 b a | *** | ||
| T100 | 615.33 ± 67.42 b | 520.47 ± 70.74 b | – | 660.39 ± 103.37 b | 563.75 ± 130.52 b | 242.11 ± 16.74 ab a | *** | ||
| T C.l. 1 | ns | ns | ns | ns | * | ||||
| Block II | T0 | 483.1 ± 156.35 ab | 617.93 ± 126.75 b | 467.19 ± 103.6 b | 493.33 ± 255.47 ab | 239.77 ± 32.87 a | 447.04 ± 243.39 ab | *** | |
| T60 | 373.8 ± 84.87 ab | 526.97 ± 34.12 b | 641.98 ± 246.54 b | 456.88 ± 96.39 ab | 218.6 ± 85.57 a | 562.05 ± 76.75 b | *** | ||
| T100 | 552.98 ± 83.29 ab | 834.96 ± 323.17 b | 708.25 ± 146.35 b | 681.87 ± 130.21 b | 292.13 ± 121.8 a | 673.81 ± 134.62 b | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ns | |||
| Block III | T0 | 148.14 ± 51.6 a | 110.53 ± 38.48 a | 262.51 ± 45.06 a | 177.9 ± 70.64 a | 140.06 ± 35.54 a | 133.85 ± 38.26 a | ns | |
| T60 | 755.95 ± 423.05 b b | 327.49 ± 91.74 b ab | 682.26 ± 121.05 b b | 416.36 ± 141.23 b ab | 314.49 ± 86.94 b a | 508.77 ± 30.26 b ab | * | ||
| T100 | 339.86 ± 105.64 a | 378.95 ± 131.84 ab | 272.9 ± 144.47 a | 274.42 ± 78.01 ab | 148.33 ± 79.51 ab | 209.23 ± 102.90 a | ns | ||
| T C.l. 1 | *** | *** | *** | *** | *** | *** |
| Parameter 2 | Factor/Interaction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment (T) | Season (S) | T × S | Residuals | ||||||||
| Block | F | df | p-Value 1 | F | df | p-Value 1 | F | df | p-Value 1 | df | |
| β-glucosidase (SQRT-transformed) | I | 2.47 | 2 | 0.1012 ns | 29.05 | 4 | <0.0001 *** | 1.34 | 8 | 0.2626 ns | 30 |
| II | 3.17 | 2 | 0.0539 ns | 6.21 | 5 | 0.0003 *** | 0.76 | 10 | 0.6632 ns | 36 | |
| III | 13.48 | 2 | <0.0001 *** | 4.84 | 5 | 0.0017 ** | 0.87 | 10 | 0.5689 ns | 36 | |
| Acid phosphatase (SQR- transformed) | I | 1.70 | 2 | 0.1998 ns | 6.80 | 4 | 0.0005 *** | 0.67 | 8 | 0.7101 ns | 30 |
| II | 7.88 | 2 | 0.0014 ** | 5.03 | 5 | 0.0014 ** | 0.38 | 10 | 0.9472 ns | 36 | |
| III | 14.03 | 2 | <0.0001 *** | 5.92 | 5 | 0.0004 *** | 1.36 | 10 | 0.2358 ns | 36 | |
| Urease (non-transformed) | I | 0.22 | 2 | 0.8025 ns | 3.12 | 4 | 0.0293 * | 2.29 | 8 | 0.0477 * | 30 |
| II | 8.29 | 2 | 0.0011 ** | 3.42 | 5 | 0.0125 * | 1.48 | 10 | 0.1864 ns | 36 | |
| III | 5.69 | 2 | 0.0071 ** | 8.15 | 5 | <0.0001 *** | 0.97 | 10 | 0.4853 ns | 36 | |
| Dehydrogenase (SQRT-transformed) | I | 1.47 | 2 | 0.2463 ns | 92.05 | 4 | <0.0001 *** | 1.11 | 8 | 0.3841 ns | 30 |
| II | 4.60 | 2 | 0.0167 * | 6.38 | 5 | 0.0002 *** | 0.54 | 10 | 0.8491 ns | 36 | |
| III | 32.04 | 2 | <0.0001 *** | 8.4 | 5 | <0.0001 *** | 0.74 | 10 | 0.6847 ns | 36 | |
| Parameter 2 | Treatment | Spring 1st | Summer 1st | Autumn 1st | Spring 2nd | Summer 2nd | Autumn 2nd | S C.l. 1 | |
|---|---|---|---|---|---|---|---|---|---|
| β-Glucosidase | Block I | T0 | 0.78 ± 0.228 a | 1.26 ± 0.23 b | – | 1.6 ± 0.27 bc | 1.28 ± 0.35 bc | 2.16 ± 0.13 c | *** |
| T60 | 0.75 ± 0.282 a | 1.28 ± 0.47 ab | – | 1.92 ± 0.2 bc | 1.9 ± 0.27 bc | 2.14 ± 0.30 c | *** | ||
| T100 | 0.71 ± 0.040 a | 1.49 ± 0.25 b | – | 2.34 ± 0.82 b | 1.96 ± 0.26 b | 1.96 ± 0.22 b | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ||||
| Block II | T0 | 1.23 ± 0.35 ab | 0.86 ± 0.38 ab | 0.85 ± 0.35 a | 1.42 ± 0.68 ab | 1.11 ± 0.40 ab | 2.10 ± 0.89 b | *** | |
| T60 | 0.95 ± 0.43 ab | 0.96 ± 0.27 a | 1.02 ± 0.36 a | 1.73 ± 0.52 b | 1.51 ± 0.18 ab | 1.40 ± 0.69 b | ** | ||
| T100 | 1.27 ± 0.15 ab | 1.25 ± 0.27 ab | 0.94 ± 0.16 a | 2.05 ± 0.12 ab | 1.87 ± 0.87 ab | 2.14 ± 0.35 b | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ns | |||
| Block III | T0 | 0.53 ± 0.17 a ab | 0.25 ± 0.18 a a | 0.49 ± 0.09 a ab | 0.44 ± 0.12 a ab | 0.67 ± 0.11 a ab | 0.56 ± 0.11 a b | ** | |
| T60 | 0.94 ± 0.4 b ab | 0.49 ± 0.15 b a | 0.99 ± 0.43 b ab | 0.95 ± 0.36 b ab | 0.91 ± 0.13 b b | 1.28 ± 0.09 b b | ** | ||
| T100 | 0.67 ± 0.1 ab | 0.52 ± 0.3 ab | 0.44 ± 0.17 ab | 0.79 ± 0.06 ab | 0.75 ± 0.26 ab | 0.76 ± 0.39 ab | ns | ||
| T C.l. 1 | * | ** | ** | ** | * | *** | |||
| Phosphatase | Block I | T0 | 2.5 ± 0.5 a | 3.62 ± 0.63 ab | – | 2.858 ± 0.31 ab | 3.545 ± 0.85 ab | 3.89 ± 1.44 b | ** |
| T60 | 2.19 ± 0.6 a | 3.97 ± 1.64 ab | – | 2.535 ± 0.21 ab | 2.981 ± 0.41 ab | 4.70 ± 0.64 b | *** | ||
| T100 | 2.26 ± 0.42 a | 3.67 ± 1.20 b | – | 2.487 ± 0.07 ab | 2.490 ± 0.56 ab | 3.14 ± 0.60 b | ** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ||||
| Block II | T0 | 1.73 ± 0.62 a ab | 2.37 ± 0.84 a ab | 1.16 ± 0.41 a a | 1.86 ± 1.17 a a | 2.31 ± 1.37 a ab | 3.37 ± 1.25 b | ** | |
| T60 | 2.43 ± 0.77 ab ab | 2.59 ± 0.39 ab ab | 2.22 ± 0.12 ab a | 1.65 ± 0.49 ab a | 2.48 ± 0.58 ab ab | 3.42 ± 1.84 b | * | ||
| T100 | 3.02 ± 0.24 b ab | 3.61 ± 0.42 b ab | 2.14 ± 0.52 b a | 2.52 ± 0.46 b a | 3.61 ± 0.58 b ab | 4.15 ± 1.06 b | ** | ||
| T C.l. 1 | * | * | * | * | * | ns | |||
| Block III | T0 | 2.17 ± 0.26 a b | 0.69 ± 0.15 a a | 1.6 ± 0.6 a ab | 1.57 ± 0.74 a ab | 1.46 ± 0.47 a ab | 2.17 ± 0.7 a ab | *** | |
| T60 | 4.05 ± 1.74 b b | 2.02 ± 0.74 b a | 2.45 ± 0.6 b ab | 2.58 ± 0.75 b ab | 2.13 ± 0.16 ab ab | 3.7 ± 0.53 b ab | ** | ||
| T100 | 3.48 ± 0.37 ab | 2.98 ± 0.81 ab | 1.93 ± 1.42 ab | 2.09 ± 0.36 ab | 2.55 ± 0.98 b | 2.89 ± 0.28 ab | ns | ||
| T C.l. 1 | ** | *** | * | * | * | ** | |||
| Urease | Block I | T0 | 5.04 ± 0.85 | 5.6 ± 1.56 | – | 4.75 ± 0.82 | 4.3 ± 1.63 | 5.76 ± 0.69 | ns |
| T60 | 3.33 ± 1.02 a | 6.29 ± 1.93 ab | – | 4.12 ± 0.97 ab | 3.59 ± 1.07 ab | 7.51 ± 2 b | ** | ||
| T100 | 4.74 ± 0.59 | 5.62 ± 0.81 | – | 4.96 ± 0.64 | 6.31 ± 2.29 | 4.75 ± 0.33 | ns | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ||||
| Block II | T0 | 3.85 ± 0.7 ab | 4.82 ± 1.19 b | 3.18 ± 1.46 a ab | 4.14 ± 1.77 a ab | 3.11 ± 1.24 a ab | 1.93 ± 1.18 a | * | |
| T60 | 4.05 ± 1.31 | 3.86 ± 0.22 | 5.58 ± 2.72 ab | 3.71 ± 0.97 a | 3.98 ± 1.66 ab | 3.74 ± 1.34 | ns | ||
| T100 | 4.75 ± 0.47 ab | 5.96 ± 1.78 b | 5.63 ± 0.94 b ab | 6.39 ± 0.69 b b | 6.4 ± 0.3 b ab | 2.52 ± 1.3 a | ** | ||
| T C.l. 1 | ns | ns | * | * | ** | ns | |||
| Block III | T0 | 4.01 ± 0.38 a b | 3.11 ± 1.06 ab | 1.08 ± 0.38 a | 2.05 ± 1.06 ab | 1.47 ± 0.64 ab | 2.49 ± 0.99 a ab | *** | |
| T60 | 6.59 ± 5.07 b c | 3.46 ± 0.89 abc | 1.17 ± 0.04 a | 4.07 ± 1.27 abc | 2.57 ± 0.45 ab | 6.45 ± 0.83 b bc | *** | ||
| T100 | 4.89 ± 1.08 ab b | 4.12 ± 1.69 ab | 1.29 ± 1.15 a | 2.64 ± 1.23 ab | 0.99 ± 0.42 a | 2.87 ± 1.86 a ab | *** | ||
| T C.l. 1 | * | ns | ns | ns | ns | * | |||
| Dehydrogenase | Block I | T0 | 0.09 ± 0.02 a | 0.16 ± 0.07 b | – | 0.41 ± 0.1 c | 0.4 ± 0.01 c | 0.23 ± 0.04 b | *** |
| T60 | 0.08 ± 0.01 a | 0.23 ± 0.05 b | – | 0.46 ± 0.05 d | 0.35 ± 0.05 cd | 0.3 ± 0.03 bc | *** | ||
| T100 | 0.08 ± 0.03 a | 0.21 ± 0.06 b | – | 0.45 ± 0.03 d | 0.41 ± 0.05 cd | 0.29 ± 0.02 bc | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ||||
| Block II | T0 | 0.32 ± 0.11 ab | 0.16 ± 0.07 a | 0.34 ± 0.1 b | 0.33 ± 0.14 b | 0.33 ± 0.08 ab | 0.27 ± 0.08 ab | ** | |
| T60 | 0.35 ± 0.14 ab | 0.16 ± 0.06 a | 0.51 ± 0.13 b | 0.32 ± 0.05 ab | 0.27 ± 0.01 ab | 0.22 ± 0.13 ab | *** | ||
| T100 | 0.44 ± 0.18 ab | 0.26 ± 0.17 a | 0.44 ± 0.04 b | 0.48 ± 0.12 ab | 0.4 ± 0.14 ab | 0.34 ± 0.02 ab | *** | ||
| T C.l. 1 | ns | ns | ns | ns | ns | ns | |||
| Block III | T0 | 0.15 ± 0.04 a ab | 0.08 ± 0.002 a a | 0.21 ± 0.02 a b | 0.12 ± 0.06 a ab | 0.17 ± 0.03 a ab | 0.11 ± 0.02 a ab | ** | |
| T60 | 0.54 ± 0.3 b b | 0.18 ± 0.05 b a | 0.38 ± 0.09 b b | 0.29 ± 0.15 b ab | 0.35 ± 0.02 b ab | 0.27 ± 0.02 b ab | *** | ||
| T100 | 0.25 ± 0.06 a ab | 0.12 ± 0.02 ab a | 0.29 ± 0.11 ab b | 0.15 ± 0.05 ab ab | 0.18 ± 0.05 ab ab | 0.15 ± 0.05 ab ab | * | ||
| T C.l. 1 | *** | *** | *** | *** | *** | *** |
References
- Bonari, G.; Fernández-González, F.; Çoban, S.; Monteiro-Henriques, T.; Bergmeier, E.; Didukh, Y.P.; Xystrakis, F.; Angiolini, C.; Chytrý, K.; Acosta, A.T.R.; et al. Classification of the Mediterranean lowland to submontane pine forest vegetation. Appl. Veg. Sci. 2021, 24, e12544. [Google Scholar] [CrossRef]
- Quézel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; G. Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 1–12. [Google Scholar]
- de las Heras, J.; Moya, D.; Vega, J.A.; Daskalakou, E.; Vallejo, V.R.; Grigoriadis, N.; Tsitsoni, T.; Baeza, J.; Valdecantos, A.; Fernández, C.; et al. Post-fire management of serotinous pine forests. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 121–150. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J.; Bautista, S.; Bellot, J. Does Pinus halepensis facilitate the establishment of shrubs in Mediterranean semi-arid afforestations? For. Ecol. Manag. 2003, 176, 147–160. [Google Scholar] [CrossRef]
- Resco de Dios, V. What is a Mediterranean pine? Physiological trade-offs under stress and perturbation. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Osem, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 103–115. [Google Scholar] [CrossRef]
- Rodríguez-García, E.; Santana, V.M.; Alloza, J.A.; Ramón Vallejo, V. Predicting natural hyperdense regeneration after wildfires in Pinus halepensis (Mill.) forests using prefire site factors, forest structure and fire severity. For. Ecol. Manag. 2022, 512, 120164. [Google Scholar] [CrossRef]
- Bueis, T.; Bravo, F.; Pando, V.; Turrión, M.-B. Site factors as predictors for Pinus halepensis Mill. productivity in Spanish plantations. Ann. For. Sci. 2017, 74, 6. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Are Pinus halepensis plantations useful as a restoration tool in semiarid Mediterranean areas? For. Ecol. Manag. 2004, 198, 303–317. [Google Scholar] [CrossRef]
- Ruiz-Navarro, A.; Barberá, G.G.; Navarro-Cano, J.A.; Albaladejo, J.; Castillo, V.M. Soil dynamics in Pinus halepensis reforestation: Effect of microenvironments and previous land use. Geoderma 2009, 153, 353–361. [Google Scholar] [CrossRef]
- MITECO (Ministerio para la Transición Ecológica y el Reto Demográfico). Spanish Forest Strategy 2050. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/politica-forestal/estrategiaforestalespanolahorizonte2050_tcm30-549806.pdf (accessed on 16 June 2023).
- Jiménez, M.N.; Spotswood, E.N.; Cañadas, E.M.; Navarro, F.B. Stand management to reduce fire risk promotes understorey plant diversity and biomass in a semi-arid Pinus halepensis plantation. Appl. Veg. Sci. 2015, 18, 467–480. [Google Scholar] [CrossRef]
- Ne’eman, G.; Goubitz, S.; Nathan, R. Reproductive traits of Pinus halepensis in the light of fire—A critical review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
- Molina, A.J.; del Campo, A.D. The effects of experimental thinning on throughfall and stemflow: A contribution towards hydrology-oriented silviculture in Aleppo pine plantations. For. Ecol. Manag. 2012, 269, 206–213. [Google Scholar] [CrossRef]
- Bellot, J.; Maestre, F.T.; Chirino, E.; Hernández, N.; de Urbina, J.O. Afforestation with Pinus halepensis reduces native shrub performance in a Mediterranean semiarid area. Acta Oecol. 2004, 25, 7–15. [Google Scholar] [CrossRef]
- Hernandez-Tecles, E.; Osem, Y.; Alfaro Sánchez, R.; Heras, J. Vegetation structure of planted versus natural Aleppo pine stands along a climatic gradient in Spain. Ann. For. Sci. 2015, 72, 641–650. [Google Scholar] [CrossRef]
- Vilà-Cabrera, A.; Coll, L.; Martínez-Vilalta, J.; Retana, J. Forest management for adaptation to climate change in the Mediterranean basin: A synthesis of evidence. For. Ecol. Manag. 2018, 407, 16–22. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Rathgeber, C.; Bonnesoeur, V.; Bravo, F. Thinning has a positive effect on growth dynamics and growth–climate relationships in Aleppo pine (Pinus halepensis) trees of different crown classes. Ann. For. Sci. 2014, 71, 395–404. [Google Scholar] [CrossRef]
- Calev, A.; Zoref, C.; Tzukerman, M.; Moshe, Y.; Zangy, E.; Osem, Y. High-intensity thinning treatments in mature Pinus halepensis plantations experiencing prolonged drought. Eur. J. For. Res. 2016, 135, 551–563. [Google Scholar] [CrossRef]
- Nunes, A.; Oliveira, G.; Cabral, M.S.; Branquinho, C.; Correia, O. Beneficial effect of pine thinning in mixed plantations through changes in the understory functional composition. Ecol. Eng. 2014, 70, 387–396. [Google Scholar] [CrossRef]
- Navarro, F.B.; Romero-Freire, A.; Del Castillo, T.; Foronda, A.; Jiménez, M.N.; Ripoll, M.A.; Sánchez-Miranda, A.; Huntsinger, L.; Fernández-Ondoño, E. Effects of thinning on litterfall were found after years in a Pinus halepensis afforestation area at tree and stand levels. For. Ecol. Manag. 2013, 289, 354–362. [Google Scholar] [CrossRef]
- Lado-Monserrat, L.; Lidón, A.; Bautista, I. Erratum to: Litterfall, litter decomposition and associated nutrient fluxes in Pinus halepensis: Influence of tree removal intensity in a Mediterranean forest. Eur. J. For. Res. 2016, 135, 203–214. [Google Scholar] [CrossRef]
- Lemay, A.; Krause, C.; Achim, A. Comparison of wood density in roots and stems of black spruce before and after commercial thinning. For. Ecol. Manag. 2018, 408, 94–102. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H.; Chen, X.; Wang, J.; Chen, B.; Wang, D.; Guan, Q. Soil labile organic carbon and carbon-cycle enzyme activities under different thinning intensities in Chinese fir plantations. Appl. Soil Ecol. 2016, 107, 162–169. [Google Scholar] [CrossRef]
- Bautista, I.; Lado-Monserrat, L.; Lull, C.; Lidón, A. Mid-term effects of forest thinning on N mineralization in a semi-arid Aleppo pine forest. Forests 2021, 12, 1470. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, D.; Li, W.; Sun, D.; Jin, C.; Yuan, F.; Wang, A.; Wu, J. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. For. Ecol. Manag. 2018, 429, 36–43. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.; Geng, Y.; Niu, S.; Tian, D.; Yan, T.; Liu, W.; Pan, J.; Zhao, X.; Zhang, C. Heavy thinning reduces soil organic carbon: Evidence from a 9-year thinning experiment in a pine plantation. Catena 2022, 211, 106013. [Google Scholar] [CrossRef]
- Erkan, N.; Güner, Ş.T.; Aydın, A.C. Thinning effects on stand growth, carbon stocks, and soil properties in Brutia pine plantations. Carbon Balance Manag. 2023, 18, 6. [Google Scholar] [CrossRef]
- Ma, J.; Kang, F.; Cheng, X.; Han, H. Moderate thinning increases soil organic carbon in Larix principis-rupprechtii (Pinaceae) plantations. Geoderma 2018, 329, 118–128. [Google Scholar] [CrossRef]
- Parton, W.J.; Schimel, D.S.; Cole, C.V.; Ojima, D.S. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Sci. Soc. Am. J. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Zou, X.M.; Ruan, H.H.; Fu, Y.; Yang, X.D.; Sha, L.Q. Estimating soil labile organic carbon and potential turnover rates using a sequential fumigation–incubation procedure. Soil Biol. Biochem. 2005, 37, 1923–1928. [Google Scholar] [CrossRef]
- Shen, Y.; Cheng, R.; Xiao, W.; Yang, S.; Guo, Y.; Wang, N.; Zeng, L.; Lei, L.; Wang, X. Labile organic carbon pools and enzyme activities of Pinus massoniana plantation soil as affected by understory vegetation removal and thinning. Sci. Rep. 2018, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.J.; Bautista, I.; Lull, C.; del Campo, A.; González-Sanchis, M.; Lidón, A. Effects of thinning intensity on forest floor and soil biochemical properties in an Aleppo pine plantation after 13 years: Quantity but also quality matters. Forests 2022, 13, 255. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, M.; Wang, G. Effects of thinning on soil organic carbon fractions and soil properties in Cunninghamia lanceolata stands in Eastern China. Forests 2017, 8, 198. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L.; Wang, Y.; Jiang, P.; Hu, Y.; Ouyang, S.; Wu, H.; Lei, P.; Kuzyakov, Y.; Xiang, W. Plantations thinning: A meta-analysis of consequences for soil properties and microbial functions. Sci. Total Environ. 2023, 877, 162894. [Google Scholar] [CrossRef]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties, 5.2; Weaver, R.W., Angle, S., Bottomley, P., Bezdicek, D., Smith, S., Tabatabai, A., Wollum, A., Eds.; SSSA Book Series; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Renella, G.; Wirth, S.; Islam, R. Enzyme activities in the rhizosphere of plants. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 149–166. [Google Scholar]
- Zuccarini, P.; Asensio, D.; Ogaya, R.; Sardans, J.; Peñuelas, J. Effects of seasonal and decadal warming on soil enzymatic activity in a P-deficient Mediterranean shrubland. Glob. Chang. Biol. 2020, 26, 3698–3714. [Google Scholar] [CrossRef]
- Fanin, N.; Mooshammer, M.; Sauvadet, M.; Meng, C.; Alvarez, G.; Bernard, L.; Bertrand, I.; Blagodatskaya, E.; Bon, L.; Fontaine, S.; et al. Soil enzymes in response to climate warming: Mechanisms and feedbacks. Funct. Ecol. 2022, 36, 1378–1395. [Google Scholar] [CrossRef]
- Wic Baena, C.; Andrés-Abellán, M.; Lucas-Borja, M.E.; Martínez-García, E.; García-Morote, F.A.; Rubio, E.; López-Serrano, F.R. Thinning and recovery effects on soil properties in two sites of a Mediterranean forest, in Cuenca Mountain (South-eastern of Spain). For. Ecol. Manag. 2013, 308, 223–230. [Google Scholar] [CrossRef]
- Hedo de Santiago, J.; Lucas-Borja, M.E.; Wic-Baena, C.; Andrés-Abellán, M.; de las Heras, J. Effects of thinning and induced drought on microbiological soil properties and plant species diversity at dry and semiarid locations. Land Degrad. Dev. 2016, 27, 1151–1162. [Google Scholar] [CrossRef]
- Trevors, J.T. Effect of substrate concentration, inorganic nitrogen, O2 concentration, temperature and pH on dehydrogenase activity in soil. Plant Soil 1984, 77, 285–293. [Google Scholar] [CrossRef]
- Zantua, M.I.; Bremner, J.M. Stability of urease in soils. Soil Biol. Biochem. 1977, 9, 135–140. [Google Scholar] [CrossRef]
- Cordero, I.; Snell, H.; Bardgett, R.D. High throughput method for measuring urease activity in soil. Soil Biol. Biochem. 2019, 134, 72–77. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Estiarte, M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Appl. Soil Ecol. 2008, 39, 223–235. [Google Scholar] [CrossRef]
- GVA (Generalitat Valenciana). Mapa de suelos de la Comunidad Valenciana. Chelva (666); Proyecto LUCDEME (Icona), Centro de Investigaciones sobre Desertificación y Conselleria d’Agricultura i Mig Ambient: Valencia, Spain, 1995; (Original in Spanish). [Google Scholar]
- Clarke Topp, G.; Parkin, G.W.; Ferre, P.A. Soil water content. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 939–961. [Google Scholar]
- Sonmez, S.; Buyuktas, D.; Okturen, F.; Citak, S. Assessment of different soil to water ratios (1:1, 1:2.5, 1:5) in soil salinity studies. Geoderma 2008, 144, 361–369. [Google Scholar] [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA Book Series: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar] [CrossRef]
- Richards, L.A.; Fireman, M. Pressure-plate apparatus for measuring moisture sorption and transmission by soils. Soil Sci. 1943, 56, 395–404. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Yakovchenko, V.; Sikora, L.J. Modified dichromate method for determining low concentrations of extractable organic carbon in soil. Commun. Soil Sci. Plant Anal. 1998, 29, 421–433. [Google Scholar] [CrossRef]
- Aoyama, M.; Nagumo, T. Effects of heavy metal accumulation in apple orchard soils on microbial biomass and microbial activities. Soil Sci. Plant Nutr. 1997, 43, 601–612. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ceccanti, B.; Cervelli, S.; Matarese, E. Extraction of phosphatase, urease, proteases, organic carbon, and nitrogen from soil. Soil Sci. Soc. Am. J. 1980, 44, 1011–1016. [Google Scholar] [CrossRef]
- Camiña, F.; Trasar-Cepeda, C.; Gil-Sotres, F.; Leirós, C. Measurement of dehydrogenase activity in acid soils rich in organic matter. Soil Biol. Biochem. 1998, 30, 1005–1011. [Google Scholar] [CrossRef]
- Torrent, J. Mediterranean soils. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; pp. 418–427. [Google Scholar]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; del Campo, A.D.; Colangelo, M.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728, 138536. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef]
- Maurya, S.; Abraham, J.S.; Somasundaram, S.; Toteja, R.; Gupta, R.; Makhija, S. Indicators for assessment of soil quality: A mini-review. Environ. Monit. Assess. 2020, 192, 604. [Google Scholar] [CrossRef] [PubMed]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Liu, Z.F.; Fu, B.J.; Liu, G.H.; Zhu, Y.G. Soil quality: Concept, indicators and its assessment. Acta Ecol. Sin. 2006, 26, 901–913. [Google Scholar]
- Thakur, P.; Paliyal, S.S.; Dev, P.; Datt, N. Methods and approaches—Soil quality indexing, minimum data set selection & interpretation—A critical review. Commun. Soil Sci. Plant Anal. 2022, 53, 1849–1864. [Google Scholar] [CrossRef]
- Castellano, M.J.; Lewis, D.B.; Kaye, J.P. Response of soil nitrogen retention to the interactive effects of soil texture, hydrology, and organic matter. J. Geophys. Res. Biogeosci. 2013, 118, 280–290. [Google Scholar] [CrossRef]
- Saxton, K.E.; Rawls, W.J. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef]
- Vinhal-Freitas, I.C.; Corrêa, G.F.; Wendling, B.; Bobuľská, L.; Ferreira, A.S. Soil textural class plays a major role in evaluating the effects of land use on soil quality indicators. Ecol. Indic. 2017, 74, 182–190. [Google Scholar] [CrossRef]
- Hillel, D. Fundamentals of Soil Physics, 1st ed.; Academic Press, Inc.: London, UK, 1980; pp. 1–413. [Google Scholar]
- McKinley, D.; Rice, C.; Blair, J. Conversion of grassland to coniferous woodland has limited effects on soil nitrogen cycle processes. Soil Biol. Biochem. 2008, 40, 2627–2633. [Google Scholar] [CrossRef]
- Erdozain, M.; Bonet, J.A.; Martínez de Aragón, J.; de-Miguel, S. Forest thinning and climate interactions driving early-stage regeneration dynamics of maritime pine in Mediterranean areas. For. Ecol. Manag. 2023, 539, 121036. [Google Scholar] [CrossRef]
- Centenaro, G.; de Miguel, S.; Amouzgar, L.; Piñuela, Y.; Son, D.; Bonet, J.A.; de Aragón, J.M.; Dashevskaya, S.; Castaño, C.; Alday, J.G. Silvicultural management and altitude prevail on soil properties and fungal community in shaping understorey plant communities in a Mediterranean pine forest. Sci. Total Environ. 2023, 858, 159860. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Zhang, W.; Chen, Y. Heavy thinning temporally reduced soil carbon storage by intensifying soil microbial phosphorus limitation. Plant Soil 2023, 484, 33–48. [Google Scholar] [CrossRef]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.P.; et al. Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Effect of organic matter on the water-holding capacity and the wilting point of mineral soils. Soil Sci. 1939, 47, 377–384. [Google Scholar] [CrossRef]
- Lull, C.; Bautista, I.; Lidón, A.; del Campo, A.D.; González-Sanchis, M.; García-Prats, A. Temporal effects of thinning on soil organic carbon pools, basal respiration and enzyme activities in a Mediterranean Holm oak forest. For. Ecol. Manag. 2020, 464, 118088. [Google Scholar] [CrossRef]
- Zhang, Z.; Hao, M.; Yu, Q.; Dun, X.; Xu, J.; Gao, P. The effect of thinning intensity on the soil carbon pool mediated by soil microbial communities and necromass carbon in coastal zone protected forests. Sci. Total Environ. 2023, 881, 163492. [Google Scholar] [CrossRef] [PubMed]
- Achat, D.L.; Fortin, M.; Landmann, G.; Ringeval, B.; Augusto, L. Forest soil carbon is threatened by intensive biomass harvesting. Sci. Rep. 2015, 5, 15991. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.; Kim, D.G.; Li, J.; Liu, Y.; Hai, X.; Liu, Q.; Huang, C.; Shangguan, Z.; Kuzyakov, Y. Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems. Earth Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]
- Das, S.; Deb, S.; Sahoo, S.S.; Sahoo, U.K. Soil microbial biomass carbon stock and its relation with climatic and other environmental factors in forest ecosystems: A review. Acta Ecol. Sin. 2023; in press. [Google Scholar] [CrossRef]
- Wang, J.; Liu, H.; Hu, M.; Du, Y.; Liu, Y.; Lu, L.; Han, S. Effects of decreased precipitation and thinning on soil respiration in a temperate forest: A one-year field experiment in Central China. Catena 2023, 229, 107239. [Google Scholar] [CrossRef]
- Tang, J.; Qi, Y.; Xu, M.; Misson, L.; Goldstein, A.H. Forest thinning and soil respiration in a ponderosa pine plantation in the Sierra Nevada. Tree Physiol. 2005, 25, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Concilio, A.; Chen, J.; Ma, S.; North, M. Precipitation drives interannual variation in summer soil respiration in a Mediterranean-climate, mixed-conifer forest. Clim. Change 2009, 92, 109–122. [Google Scholar] [CrossRef]
- Fernandez, I.; Álvarez-González, J.G.; Carrasco, B.; Ruíz-González, A.D.; Cabaneiro, A. Post-thinning soil organic matter evolution and soil CO2 effluxes in temperate radiata pine plantations: Impacts of moderate thinning regimes on the forest C cycle. Can. J. For. Res. 2012, 42, 1953–1964. [Google Scholar] [CrossRef]
- Gatica-Saavedra, P.; Aburto, F.; Rojas, P.; Echeverría, C. Soil health indicators for monitoring forest ecological restoration: A critical review. Restor. Ecol. 2023, 31, e13836. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Rao, M.A.; Scelza, R.; Acevedo, F.; Diez, M.C.; Gianfreda, L. Enzymes as useful tools for environmental purposes. Chemosphere 2014, 107, 145–162. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L. Resource allocation to extracellular enzyme production: A model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol. Biochem. 2013, 56, 60–68. [Google Scholar] [CrossRef]
- Zang, X.; Liu, M.; Fan, Y.; Xu, J.; Xu, X.; Li, H. The structural and functional contributions of β-glucosidase-producing microbial communities to cellulose degradation in composting. Biotechnol. Biofuels 2018, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Harhangi, H.R.; Steenbakkers, P.J.M.; Akhmanova, A.; Jetten, M.S.M.; van der Drift, C.; Op den Camp, H.J.M. A highly expressed family 1 β-glucosidase with transglycosylation capacity from the anaerobic fungus Piromyces sp. E2. BBA-Gene Struct. Expr. 2002, 1574, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Pang, Q.; Zhao, L.; Fan, S.; Shi, H. Thermoanaerobacterium thermosaccharolyticum β-glucosidase: A glucose-tolerant enzyme with high specific activity for cellobiose. Biotechnol. Biofuels 2012, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Solhtalab, M.; Thongsomboon, W.; Aristilde, L. Strategies of organic phosphorus recycling by soil bacteria: Acquisition, metabolism, and regulation. Environ. Microbiol. Rep. 2022, 14, 3–24. [Google Scholar] [CrossRef]
- Mndzebele, B.; Ncube, B.; Fessehazion, M.; Mabhaudhi, T.; Amoo, S.; Plooy, C.; Venter, S.; Modi, A. Effects of cowpea-amaranth intercropping and fertiliser application on soil phosphatase activities, available soil phosphorus, and crop growth response. Agronomy 2020, 10, 79. [Google Scholar] [CrossRef]
- Nourbakhsh, F.; Monreal, C.M. Effects of soil properties and trace metals on urease activities of calcareous soils. Biol. Fertil.Soils 2004, 40, 359–362. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Karim, M.R.; Majumder, B.R.; Akter, F. Microbial and enzymatic activity as influenced by existing cropping pattern in the soils of Ganges floodplain. Plant Sci. Today 2019, 6, 309–314. [Google Scholar] [CrossRef]
- Adetunji, A.; Ncube, B.; Mulidzi, A.; Lewu, F. Potential use of soil enzymes as soil quality indicators in agriculture. In Frontiers in Soil and Environmental Microbiology, 1st ed.; Nayak, S.K., Mishra, B.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Chapter 6. [Google Scholar] [CrossRef]
- Kumar, S. Soil dehydrogenase enzyme activity in natural and mine soil—A review. Middle East J. Sci. Res. 2013, 13, 898–906. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z. Dehydrogenase activity in the soil environment. In Dehydrogenases; Canuto, R.S., Ed.; IntechOpen: Rijeka, Croatia, 2012; Chapter 8. [Google Scholar] [CrossRef]
- Skujiņš, J.J. Enzymes in soil. In Soil Biochemistry; McLaren, A.D., Peterson, G.H., Eds.; Marcel Dekker: New York, NY, USA, 1967; Volume 1, pp. 371–414. [Google Scholar]

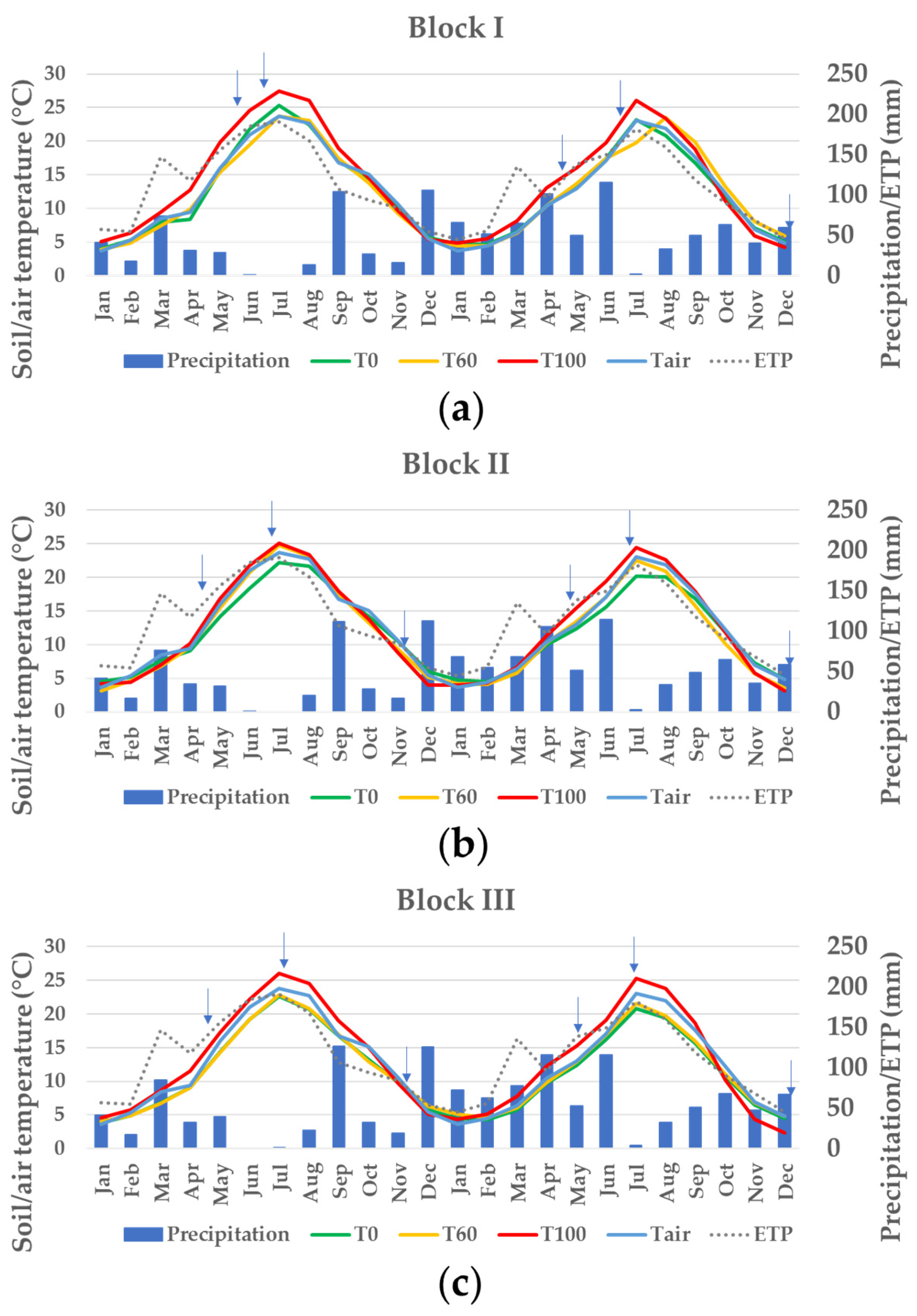

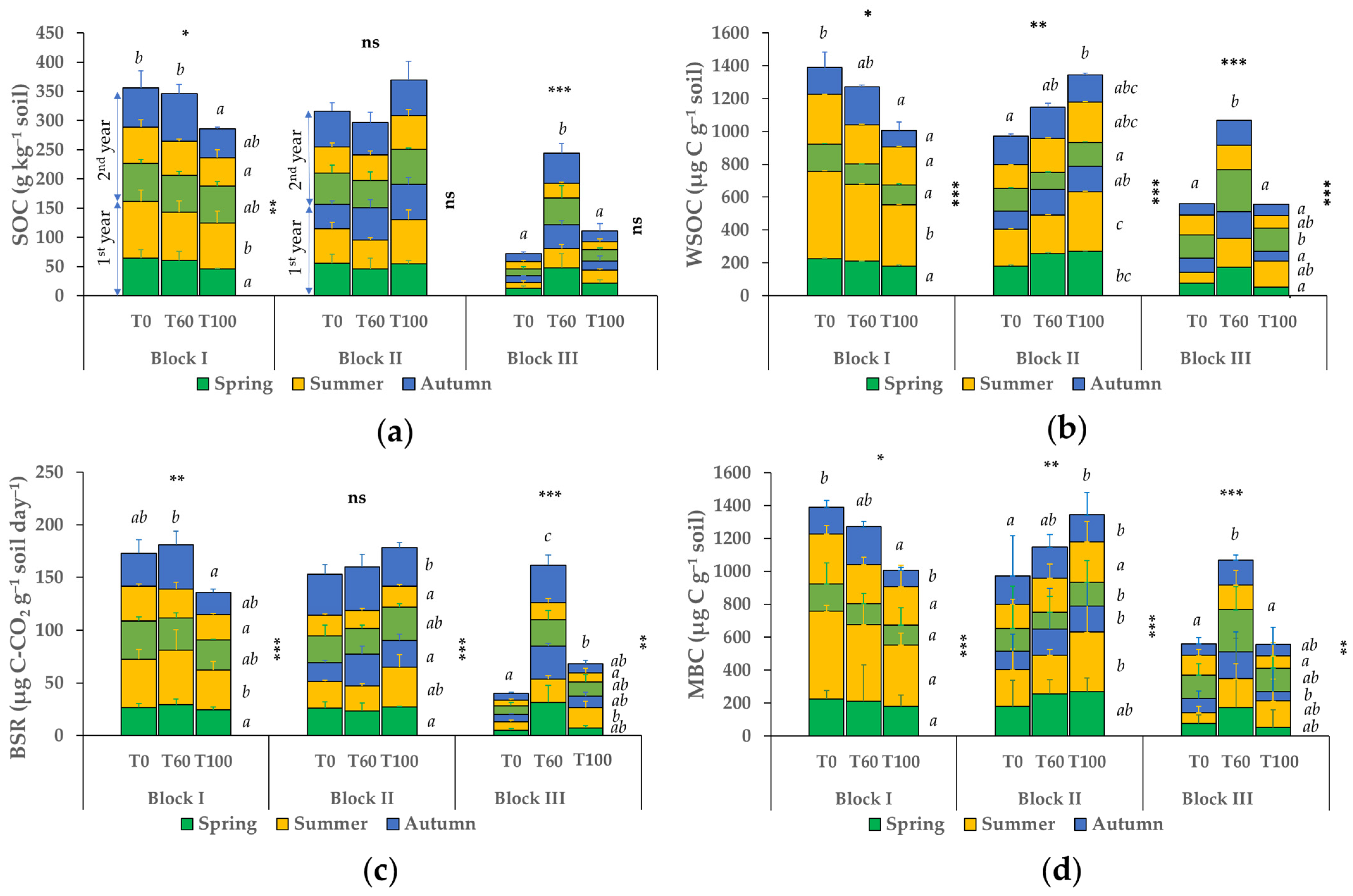
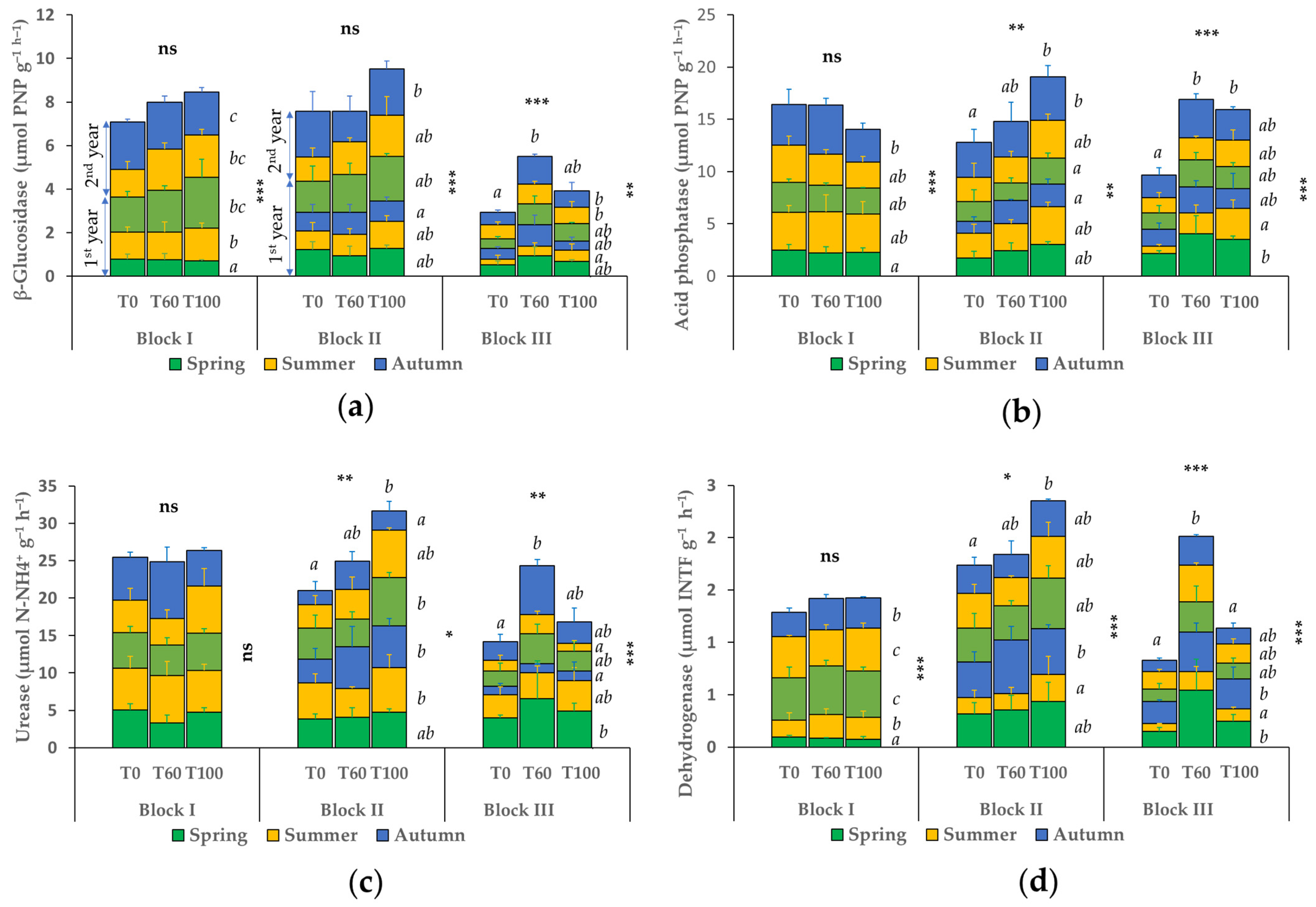

| pH | Carbonates | EC 1:2.5 1 | CEC 1 | Sand | Silt | Clay | Texture | ||
|---|---|---|---|---|---|---|---|---|---|
| (g kg−1) | (dS m−1) | (cmolc kg−1) | (%) | (%) | (%) | ||||
| Block I | T0 | 7.77 ± 0.04 a | 10.06 ± 1.11 a | 0.62 ± 0.05 a | 46.39 ± 8.83 a | 17.84 ± 3.76 a | 41.16 ± 4.30 a | 41.00 ± 5.00 a | Silty clay |
| T60 | 7.99 ± 0.23 a | 21.83 ± 14.68 ab | 0.59 ± 0.09 a | 43.35 ± 6.19 a | 19.35 ± 8.93 a | 38.98 ± 5.44 a | 41.67 ± 8.50 a | Clay | |
| T100 | 8.02 ± 0.12 a | 32.03 ± 7.18 b | 0.56 ± 0.07 a | 33.54 ± 3.36 a | 19.08 ± 8.89 a | 41.59 ± 7.05 a | 39.33 ± 9.45 a | Silty clay loam | |
| Block II | T0 | 8.14 ± 0.07 b | 45.63 ± 7.83 c | 0.54 ± 0.08 a | 34.44 ± 1.68 a | 30.64 ± 0.92 a | 36.10 ± 2.77 a | 33.27 ± 3.29 a | Clay loam |
| T60 | 7.88 ± 0.16 a | 5.15 ± 1.80 a | 0.64 ± 0.10 ab | 37.88 ± 5.08 a | 23.34 ± 5.58 a | 38.13 ± 8.20 a | 38.53 ± 6.93 a | Clay loam | |
| T100 | 8.01 ± 0.11 ab | 28.00 ± 5.39 b | 0.75 ± 0.11 b | 41.11 ± 5.61 a | 27.82 ± 5.61 a | 36.58 ± 1.39 a | 35.60 ± 5.72 a | Clay loam | |
| Block III | T0 | 7.25 ± 0.25 b | 0.55 ± 0.05 a | 0.28 ± 0.17 a | 19.63 ± 4.20 a | 79.84 ± 3.27 b | 9.82 ± 1.56 a | 10.33 ± 2.52 a | Sandy loam |
| T60 | 7.83 ± 0.15 c | 5.18 ± 1.35 b | 0.30 ± 0.03 a | 34.48 ± 3.34 b | 50.56 ± 4.18 a | 21.11 ± 1.06 b | 28.33 ± 5.13 b | Sandy clay loam | |
| T100 | 6.77 ± 0.13 a | 0.56 ± 0.10 a | 0.17 ± 0.02 a | 21.96 ± 1.45 a | 65.58 ± 8.78 a | 21.08 ± 5.65 b | 13.33 ± 6.11 a | Sandy loam |
| WHC | ||||||||||||||
| SOC | 0.86 ** | SOC | ||||||||||||
| WSOC | 0.58 ** | 0.68 ** | WSOC | |||||||||||
| BSR | 0.79 ** | 0.85 ** | 0.66 ** | BSR | ||||||||||
| MBC | 0.64 ** | 0.67 ** | 0.48 ** | 0.72 ** | MBC | |||||||||
| GLUC | 0.75 ** | 0.72 ** | 0.35 ** | 0.63 ** | 0.46 ** | GLUC | ||||||||
| PHOS | 0.48 ** | 0.57 ** | 0.44 ** | 0.56 ** | 0.33 ** | 0.54 ** | PHOS | |||||||
| UREA | 0.52 ** | 0.64 ** | 0.49 ** | 0.53 ** | 0.48 ** | 0.50 ** | 0.51 ** | UREA | ||||||
| DEHY | 0.55 ** | 0.42 ** | ns | 0.38 ** | 0.45 ** | 0.63 ** | 0.28 ** | 0.33 ** | DEHY | |||||
| pH | 0.59 ** | 0.54 ** | 0.36 ** | 0.43 ** | 0.47 ** | 0.51 ** | ns | 0.31 ** | 0.38 ** | pH | ||||
| EC | 0.73 ** | 0.67 ** | 0.52 ** | 0.58 ** | 0.51 ** | 0.57 ** | 0.28 ** | 0.45 ** | 0.40 ** | 0.52 ** | CE | |||
| CEC | 0.65 ** | 0.68 ** | 0.55 ** | 0.66 ** | 0.46 ** | 0.49 ** | 0.35 ** | 0.39 ** | 0.33 ** | 0.23 * | 0.77 ** | CEC | ||
| SAND | −0.70 ** | −0.73 ** | −0.52 ** | −0.64 ** | −0.48 ** | −0.61 ** | −0.35 ** | −0.50 ** | −0.27 ** | −0.46 ** | −0.73 ** | −0.75 ** | SAND | |
| SILT | 0.69 ** | 0.71 ** | 0.50 ** | 0.62 ** | 0.49 ** | 0.62 ** | 0.34 ** | 0.50 ** | 0.28 ** | 0.52 ** | 0.70 ** | 0.68 ** | −0.99 ** | SILT |
| CLAY | 0.71 ** | 0.75 ** | 0.53 ** | 0.66 ** | 0.49 ** | 0.62 ** | 0.36 ** | 0.47 ** | 0.28 ** | 0.47 ** | 0.73 ** | 0.81 ** | −0.97 ** | 0.95 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lull, C.; Gil-Ortiz, R.; Bautista, I.; Lidón, A. Seasonal Variation and Soil Texture-Related Thinning Effects on Soil Microbial and Enzymatic Properties in a Semi-Arid Pine Forest. Forests 2023, 14, 1674. https://doi.org/10.3390/f14081674
Lull C, Gil-Ortiz R, Bautista I, Lidón A. Seasonal Variation and Soil Texture-Related Thinning Effects on Soil Microbial and Enzymatic Properties in a Semi-Arid Pine Forest. Forests. 2023; 14(8):1674. https://doi.org/10.3390/f14081674
Chicago/Turabian StyleLull, Cristina, Ricardo Gil-Ortiz, Inmaculada Bautista, and Antonio Lidón. 2023. "Seasonal Variation and Soil Texture-Related Thinning Effects on Soil Microbial and Enzymatic Properties in a Semi-Arid Pine Forest" Forests 14, no. 8: 1674. https://doi.org/10.3390/f14081674
APA StyleLull, C., Gil-Ortiz, R., Bautista, I., & Lidón, A. (2023). Seasonal Variation and Soil Texture-Related Thinning Effects on Soil Microbial and Enzymatic Properties in a Semi-Arid Pine Forest. Forests, 14(8), 1674. https://doi.org/10.3390/f14081674