1. Introduction
Among the short rotation trees, black locust, poplar, and willow are the most important in the temperate climate zone. Under unstressful environmental conditions, poplar and willow perform better regarding their survivability and yield [
1]. However, black locust is one of the drought-resistant species, with relatively low sensitivity to the high variation in weather conditions, successfully grown in the regions classified as warm temperate dry (WTED) climate zones by the Intergovernmental Panel on Climate Change, covering approximately one-third of the area of Hungary [
2].
Widening the species palette in forestation, black locust (
Robinia pseudoacacia L.) was introduced in the 18–19th centuries in Central Europe. For example, in Hungary, it was found efficient to control blown sand that is typical in one-fifth of the country’s area. Economic reasons had priority, leading to its widespread introduction. Furthermore, it is used for many purposes, such as timber production, honey production, and land reclamation. Its timber has extraordinary quality with multipurpose utilization [
3,
4], Hungarian black locust honey is on the National Treasures list, and black locust plays a considerable role in agroforestry and in the afforestation of eroded and marginal lands [
3,
5,
6,
7,
8]. Additionally, as a carbon pool, the species contributes to the fight against global warming. Black locust is characterized by a high potential for acclimatization to Eastern European climatic conditions, suggesting high invasive potential [
9]. However, proper choice of the planting sites and cultivation technologies ensure keeping its invasive potential under control [
10,
11].
Predicted climate change is expected to lower the profitability of beech, hornbeam, sessile oak, and Turkey oak forests, e.g., in Hungary, resulting from the drift in forest climate classes toward drier and warmer categories, and an increase in summer mean temperature, in particular [
12]. As with all plants, black locust is sensitive to drought. For example, the lack of rain in areas with high atmospheric evaporative demand results in decreased sap flow density, stand transpiration, leaf area index, and stem diameter [
13]. In the Chinese Loess Plateau, the annual percentage loss of whole-plant hydraulic conductance shows strong variation due to climatic variability, which was found positively correlating with the annual potential evapotranspiration and the aridity index [
14]. Despite comparable bioclimatic conditions, the climate sensitivity of black locust stands was found to be highly variable, both temporally and spatially in Eastern Europe, suggesting a site-specific microclimate with winter and spring temperatures as key climatic drivers in the growth pattern. The growth response to the previous winter and current summer precipitation was different in the stands [
9]. Differences in air temperature and precipitation, even within the same region, such as near or within a city acting as an urban heat island, influence significantly the annual ring widths [
15]. The ecophysiological and morphological adaptation of black locust to prolonged drought—by reducing water loss through both reduced transpiration and leaf size—was demonstrated in the drier regions of Europe [
16]. The trees have different transpiration levels by age; tree and stand respiration together with stomatal and hydraulic conductance were found to be higher in the older trees [
17]. Drought memory affects plant growth performance; trees resprouting under drought conditions were found to be more drought tolerant than the well-watered ones. Black locust tolerates high variation in soil water availability without altering its water-use efficiency [
18]. In one study, planting density (i.e., short rotation forestry (1000–2000 trees ha
−1) vs. short rotation coppice (6000–15,000 trees ha
−1)) was not found to significantly affect growth dynamics and water-use efficiency [
19].
In most studies, the performance of a given clone has been tested under different environmental conditions to characterize its response to stressors such as drought, heat stress, and plantation density, as detailed above. Tree species used for wood biomass production have also been compared under the same environmental conditions to demonstrate one as an alternative to another, e.g., black locust vs. poplars (
Populus spp.) [
19] and/or willows (
Salix spp.) [
20,
21] or European linden (
Tilia × europaea) [
22]. Bhusal et al. [
23] studied and compared the growth responses of six gymnosperm species under long-term excessive irrigation. They also investigated the morphological, physiological, and biochemical traits of eleven tree species to assess their drought resistance. Highly resistant species have a large leaf mass per area and a high photosynthetic rate, according to this study [
24]. Few studies focused on the differences in the performance of black locust clones, especially in dry regions, with the aim of clonal selection [
25] or comparative analysis of selected clones [
26,
27,
28,
29]. In Hungary, forestry research to increase the yield of black locust, improve its stem quality, and enhance its drought tolerance has been ongoing for decades. This research effort resulted in several state-approved cultivars [
5,
28,
30,
31]. Suitability for the establishment of black locust plantations in marginal sites with extreme weather conditions, especially drought, can be expected to differ by clones, as proved by our preliminary investigation [
32]. It must be noted that utilization determines which parameter has priority.
The objective of this report is to describe the in-depth results of a juvenile growth and plant physiology study characterizing newly bred, vegetatively propagated, 3-year-old black locust clones. Hypothesis 1 assumes that there are significant differences between the growth (height, diameter at the breast, and increments of these) and the plant physiology traits (rate of assimilation and transpiration, stomatal conductance, intercellular CO2 concentration, leaf temperature, carboxylation, and water-use efficiency) of the investigated clones in an extreme dry year. Hypothesis 2 assumes that there are correlations among the physiological parameters. Lastly, hypothesis 3 states that the clones can be distinguished by their growth and physiological parameters.
4. Discussion
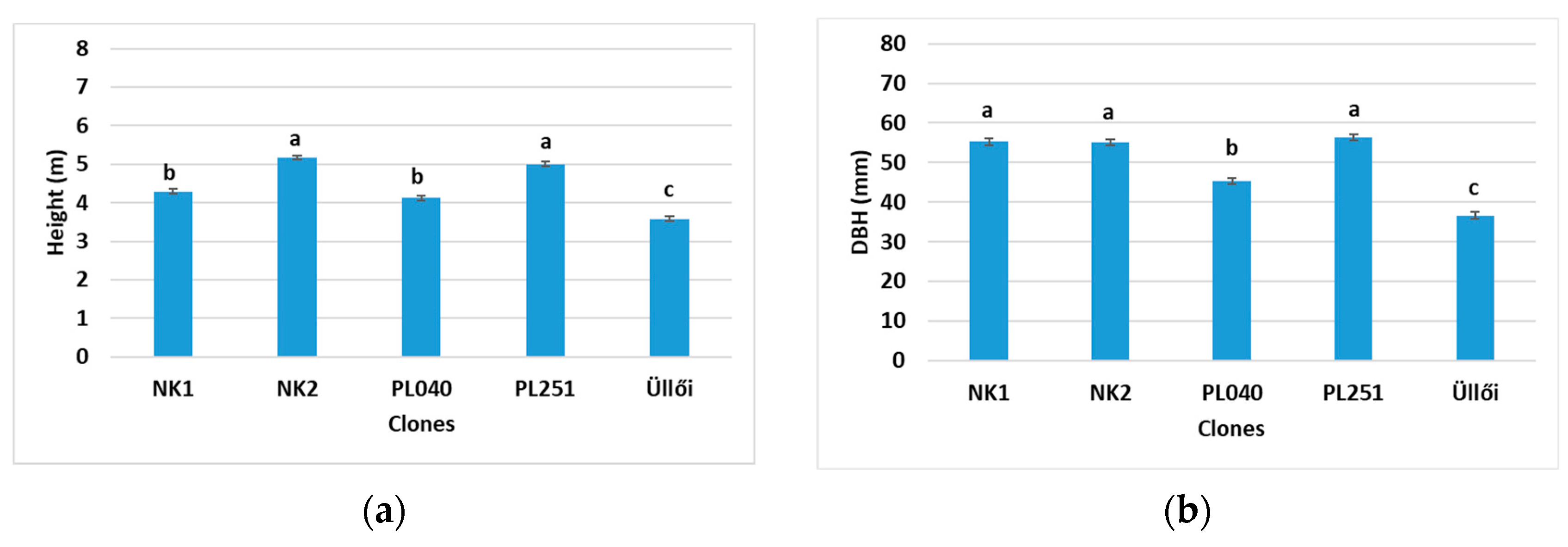
In 2022, we published the partial results from our experiment. In 2021, we studied the height, the diameter at the base, NDVI, assimilation, transpiration, and water-use efficiency of these clones only in the 3 × 3 m planting spacing. We found clone NK2 to be the best in almost all of the studied parameters [
35].
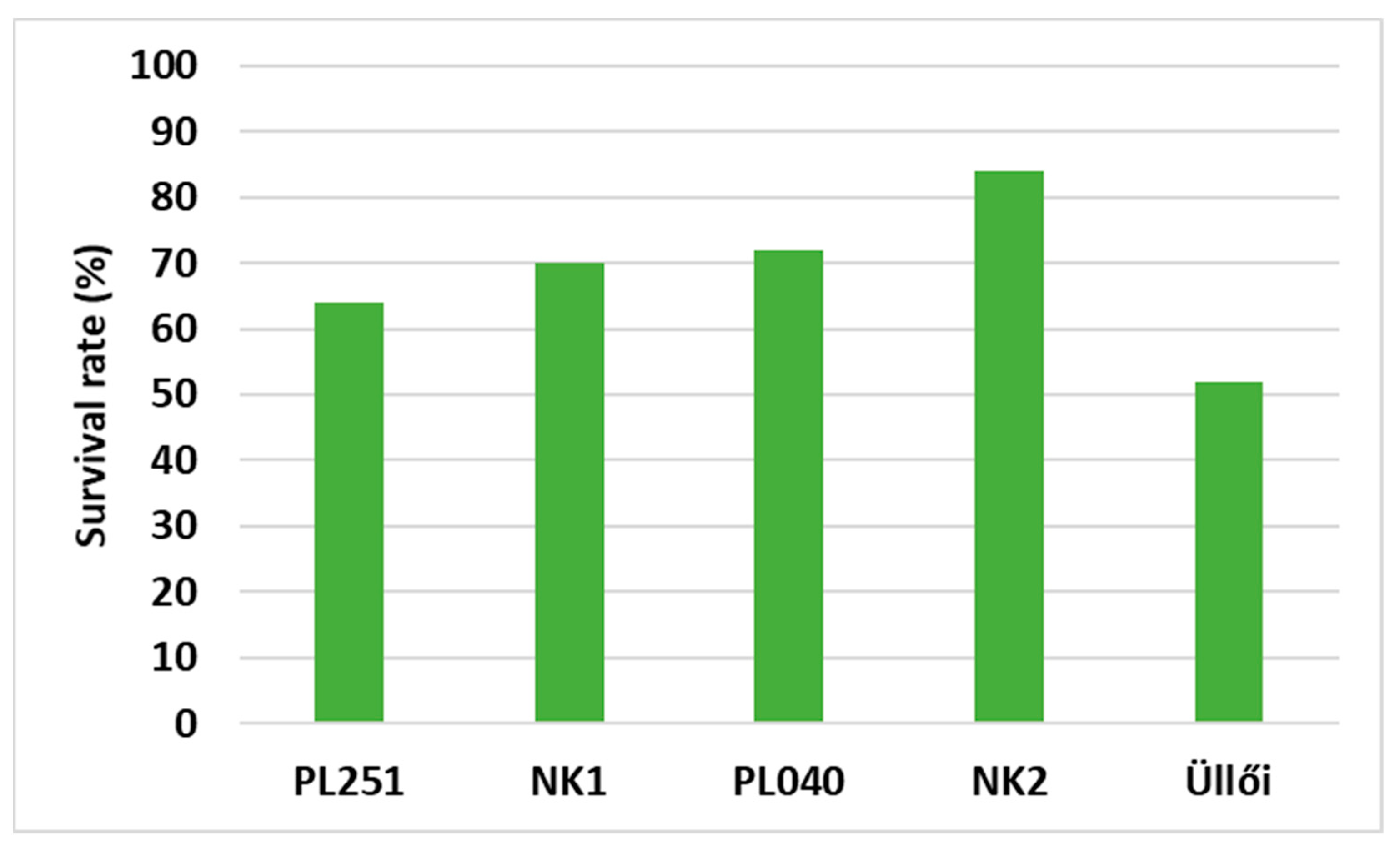
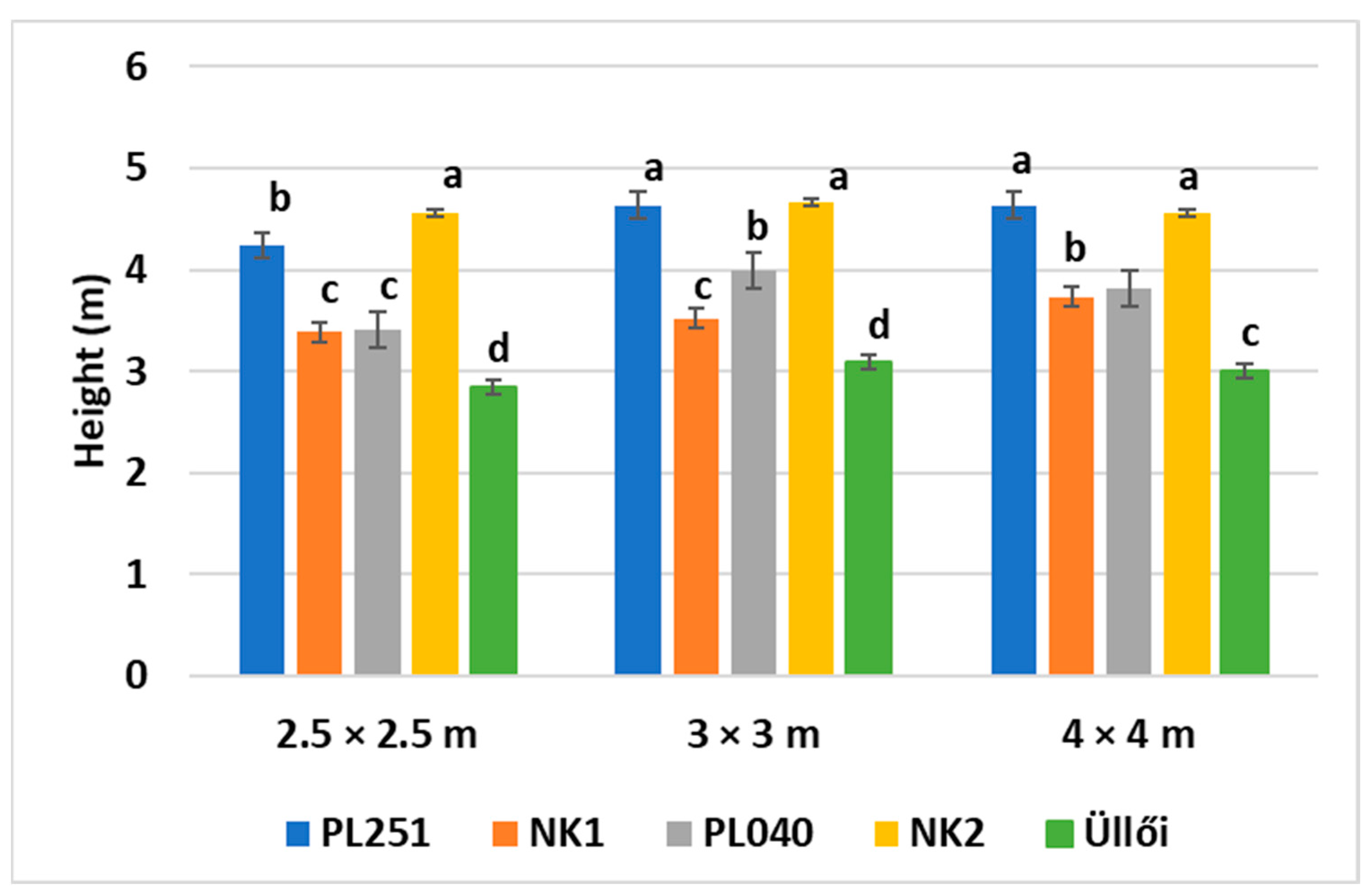
In this more recent article, we studied the survival rate and the height of the clones in all planting spacings, but we focused on the planting spacing of 2.5 × 2.5 m for the other parameters. We measured and evaluated the height and diameter increments of the clones in the main growing period (from May to August) of the sample trees and the physiological parameters. Here, we include not only the parameters we studied in 2021, but also add in the stomatal conductance, intercellular CO2 concentration, carboxylation efficiency, and the temperatures of the leaves. The correlation between the measured parameters was also analyzed according to clones.
Worldwide, tree plantations cover about 280 million hectares. Their importance is unquestionable, as approximately 80% of all industrial wood needs are met by such intensively cultivated plantations. Black locust is a fast-growing tree species with its most intensive growth in height and thickness in the first 10–15 years, depending on the site. It can grow up to 10 m in height by the age of 5, slowing after the age of 15. The diameter at breast height at 10-years-old can reach 10.8 cm [
3]. The better the site potential is, the earlier the peak of height growth. This slows after 15 years [
41], although growth remains relatively rapid in older trees; black locust stands in Europe can reach mean or dominant heights of over 20 m at 20 years and up to 25–30 m at 30 years.
The aim of our research work is to develop selected black locust varieties to improve yield and stem quality, and to achieve a uniform stand and better volume compared to common black locust at marginal sites.
The study of tree plantations is of great practical importance. In the future, a massive shortage of timber is forecast for the Central European region, especially in Hungary. Since black locust can tolerate even less favorable site conditions, the lack of timber can be reduced in the future by establishing black locust tree plantations [
32]. The economic impact of this tree species is unquestionable, so it plays a key role in forest management. However, we should mention again that the technology for growing black locust should strive to achieve a sustainable coexistence of this tree species with people and nature. It means the strict eradication of black locust at valuable sites and tolerance in some areas [
11]. Nevertheless, black locust has some positive effects on biodiversity. This finding is supported by the following facts: The black locust provides habitat for some rare and endangered plant species. Furthermore, in species-poor agricultural landscapes, black locust stands can increase diversity and play a significant role as migration corridors for forest animals [
42]. Thus, it can be concluded that black locust can also play an important role in a given habitat from a nature conservation point of view.
To be able to produce good quality industrial wood on the plantations, it is essential to study the ecophysiological properties of the trees (biomass production, photosynthetic activity, transpiration, carbon sequestration, and others) in addition to the traditional stand full inventories [
23,
24,
43].
It is well known that plants produce organic matter by using light energy. Photosynthesis is the basis for plant life. Its intensity is affected by numerous environmental factors such as CO
2 concentration, temperature, illumination, water, and nutrient availability. During assimilation, CO
2 is reduced, carbohydrates are incorporated, and oxygen is released by splitting water using the energy of incoming photons. Under stress, whether abiotic, biotic, or anthropogenic, the rate of photosynthesis decreases, with a concomitant decrease in the amount of assimilates produced and metabolic disturbances, so-called metabolic inhibition [
44,
45,
46,
47]. Nowadays, the most common stress factors are high temperatures and drought. Under water stress, stomata close and stomatal conductance becomes low, resulting in a decrease in intercellular CO
2 concentration depending on the intensity of photosynthesis [
46,
48,
49,
50]. And thus, these parameters affect tree growth [
51]. In the present study, we confirmed these effects and also found that different black locust clones respond differently to environmental stresses.
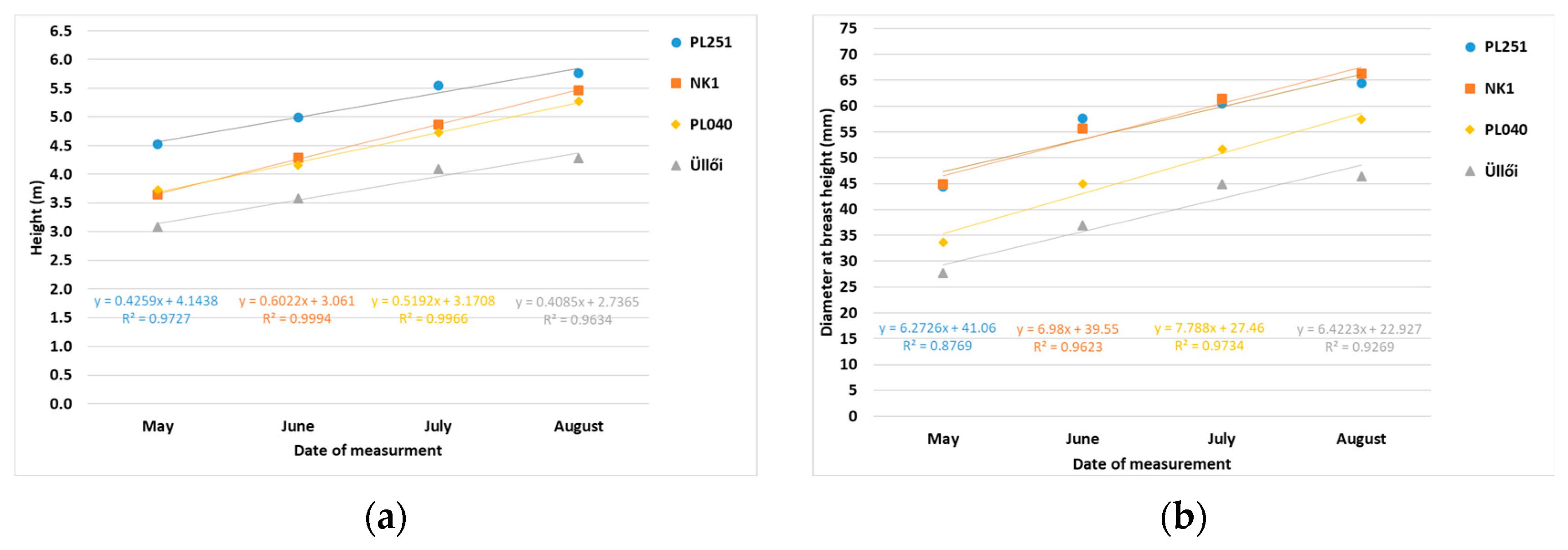
The extremely arid 2022 growing season allowed us to study black locust clones in a special situation. During the growth study, the height and diameter at breast height of sample trees or clones (plots) were measured monthly from May to August 2022. According to the measurement results, clone NK1 showed the best growth in height (1.88 m) and PL040 in diameter (23.76 mm) during the studied period. The NK2 clone was not measured because of its bark disease.
Transpiration is an important part of the hydrological cycle, where water in the soil is transferred through the plant to the air. As with photosynthesis (assimilation), transpiration in plants can be affected by several factors (environmental and anthropogenic). Jiao et al. [
17,
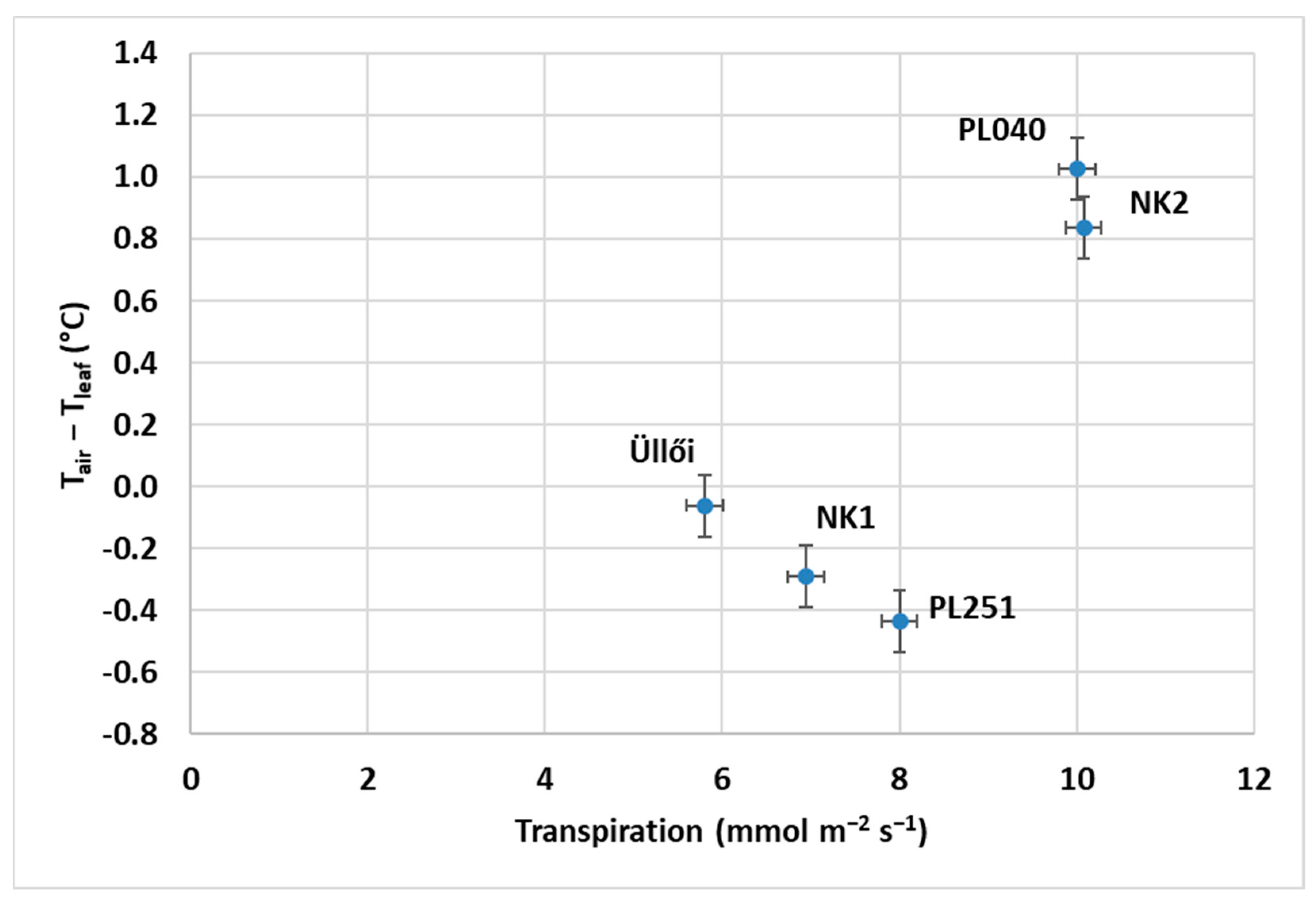
52] studied transpiration in black locust in a marginal (semiarid) site, the Loess Plateau, in northern China, southeast of the Gobi Desert. They found that as soil moisture content increases, transpiration increases, and with it, stomatal conductance. Their results also showed that older black locust trees are physiologically more active than younger ones. Our results show that the difference in transpiration between the clones is significant, and the difference in transpiration cooling effect on leaves can be clearly identified between the clones.
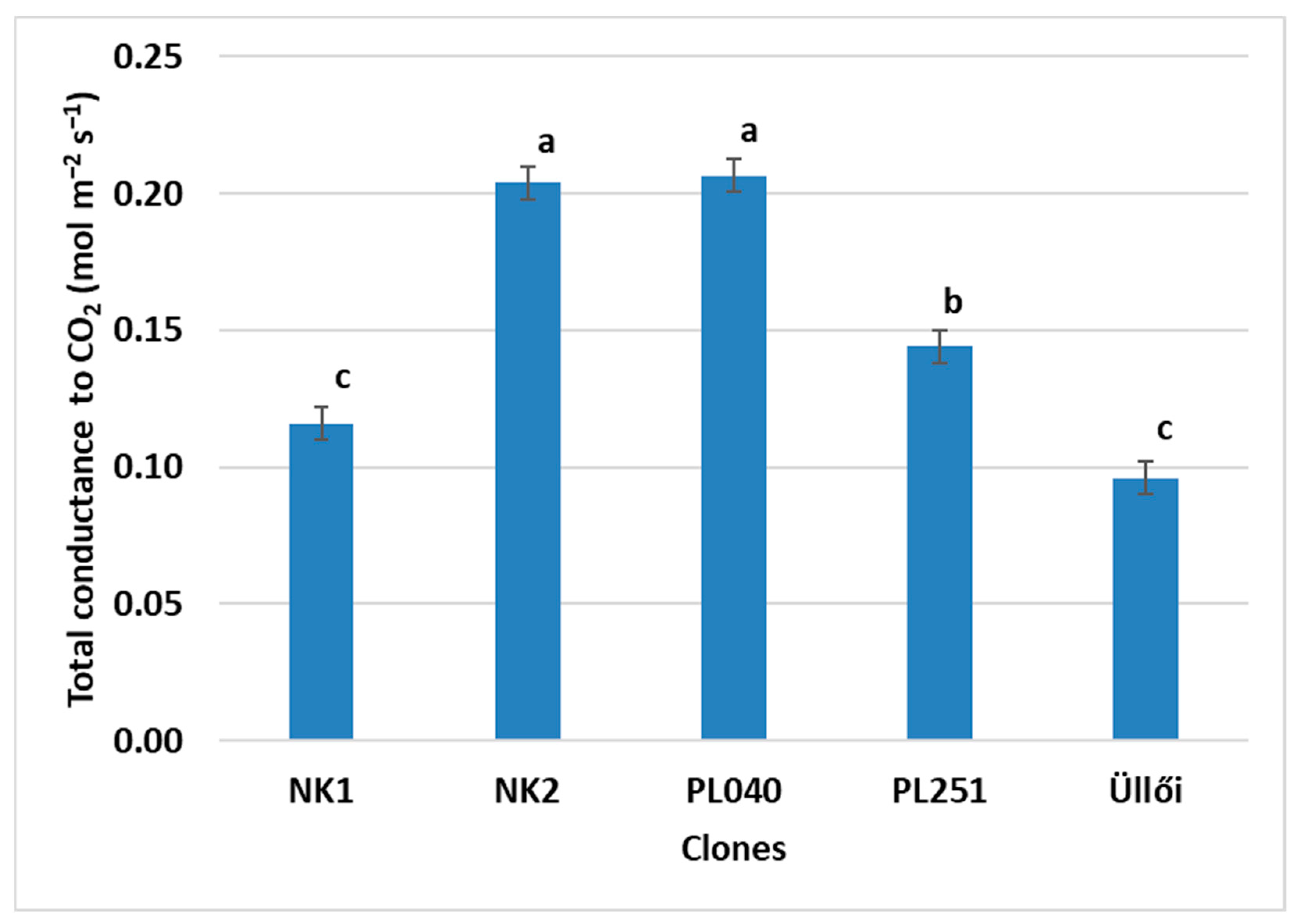
Significant differences were found between the clones for all the parameters tested. The T
leaf results were subtracted from the measured air temperature (T
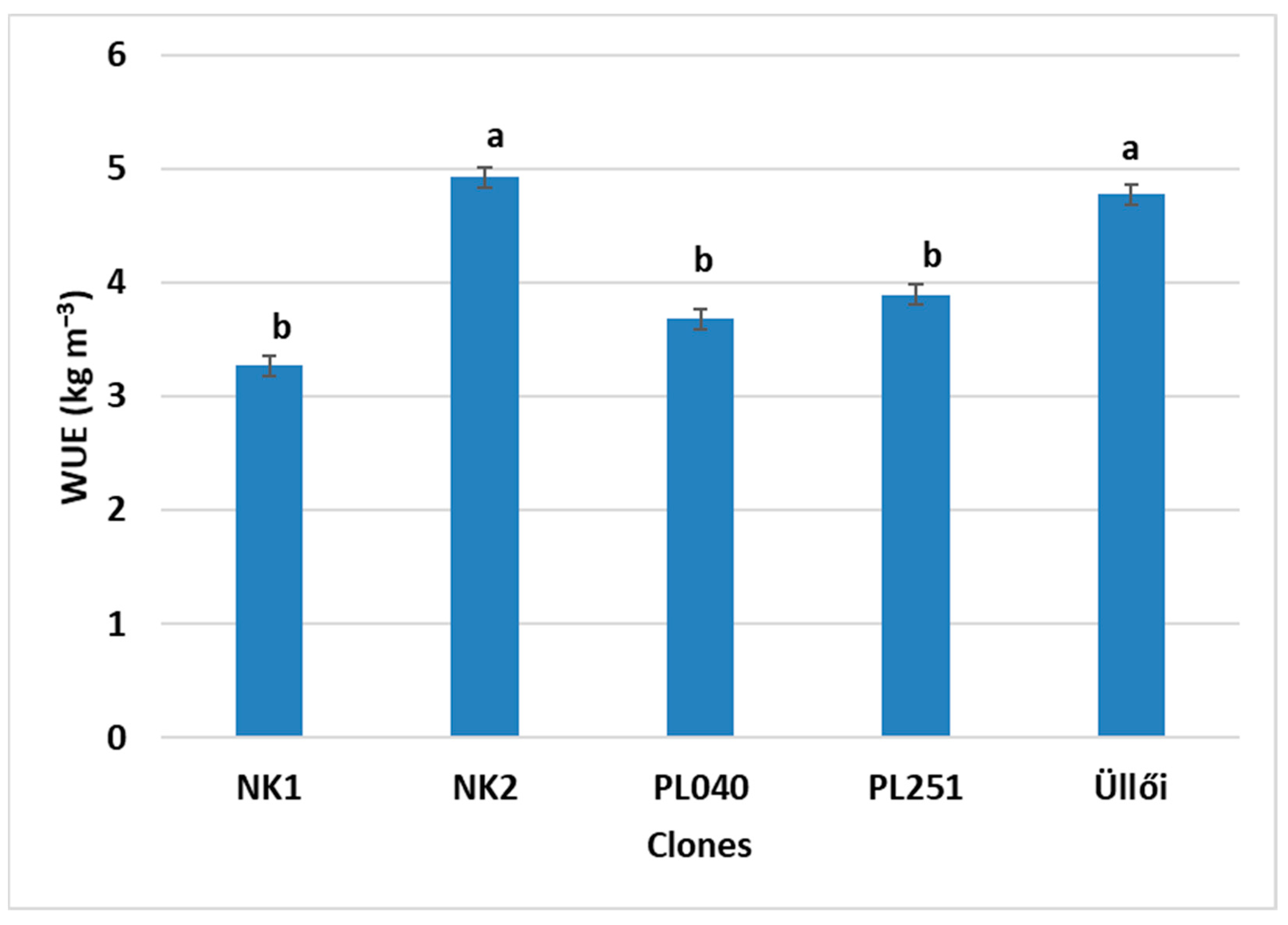
air) values. This allowed us to compare the clones on the basis of their water stress resistance. In this respect, PL040, with the highest diameter growth, proved to be the best. For CE, NK2 performed the best (0.077 µmol m
−2 s
−1) and NK1 the weakest (0.035 µmol m
−2 s
−1). However, the latter did not differ significantly from the ‘Üllői’ control. In the case of WUE, NK2 and ‘Üllői’ produced the best results with values of 4.92 and 4.78 kg m
−3, respectively, with no significant difference between them. For g
tc, the clone PL040 (0.2065 mol m
−2 s
−1) was the best and ‘Üllői’ was the weakest (0.0961 mol m
−2 s
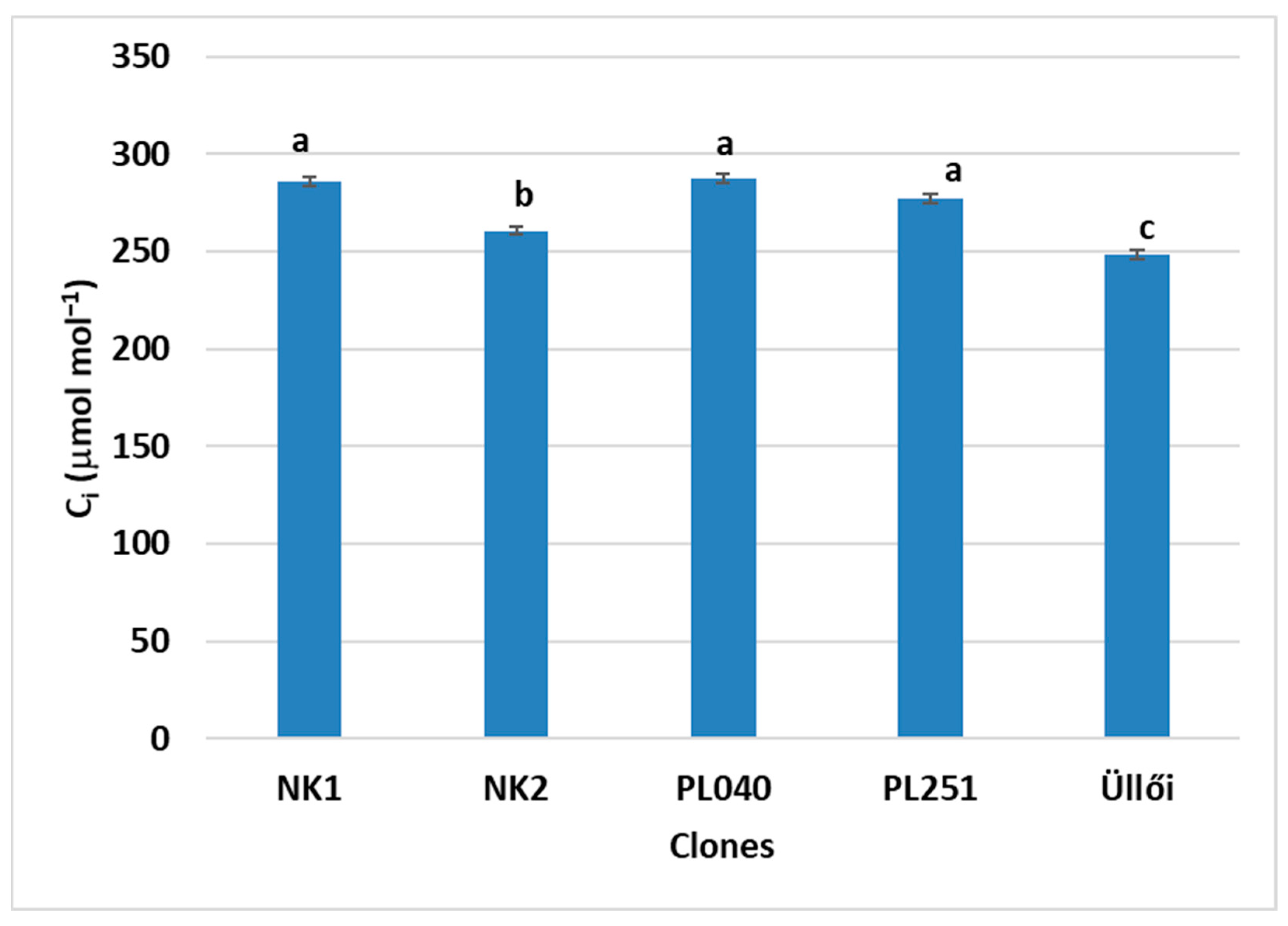
−1). For C
i, the highest values were measured for clones NK1 (286.15 µmol mol
−1) and PL040 (287.37 µmol mol
−1), and the lowest for ‘Üllői’ (248.30 µmol mol
−1). Meng et al. [
47] measured the C
i of 2-year-old diploid and tetraploid black locust, among others, which was 420 µmol mol
−1 for the diploid and 500 µmol mol
−1 for the tetraploid.
Zheng et al. [
53] measured in situ the photosynthetic gas exchange (A, Tr, g
tc and g
sw, C
i) in an 18-year-old black locust stand growing on the Loess Plateau, comparing the environmental factors that affect it. Their results show that photosynthetically active radiation, air temperature, and humidity are the most important factors affecting photosynthesis in the black locust. Later, the effects of thinning on net assimilation and carbon sequestration capacity were also studied, and it was found that an increase in these two parameters was observed with decreasing stand density [
54]. In black locust stands, similar studies using in situ phytophysiological measurements, have been carried out by several researchers on several occasions [
47,
55,
56,
57], but no such measurements have been made in a clone trial.
A correlation analysis was performed per clone to study the relationship between the ecophysiological traits. It was found that the correlation between physiological parameters varied between clones, which could be attributed to genetic differences between them. However, it should be emphasized that there was a strong positive correlation between A and Tr values for all clones (r = 0.843–0.994). For the relationship between A and g
sw, we also found a correlation (r = 0.710–0.859) for all clones, but it was very weak for NK1 (r = 0.449). According to Wang et al. [
51], A and g
sw are linearly correlated with either growth or WUE, so these parameters are useful predictors of tree growth.
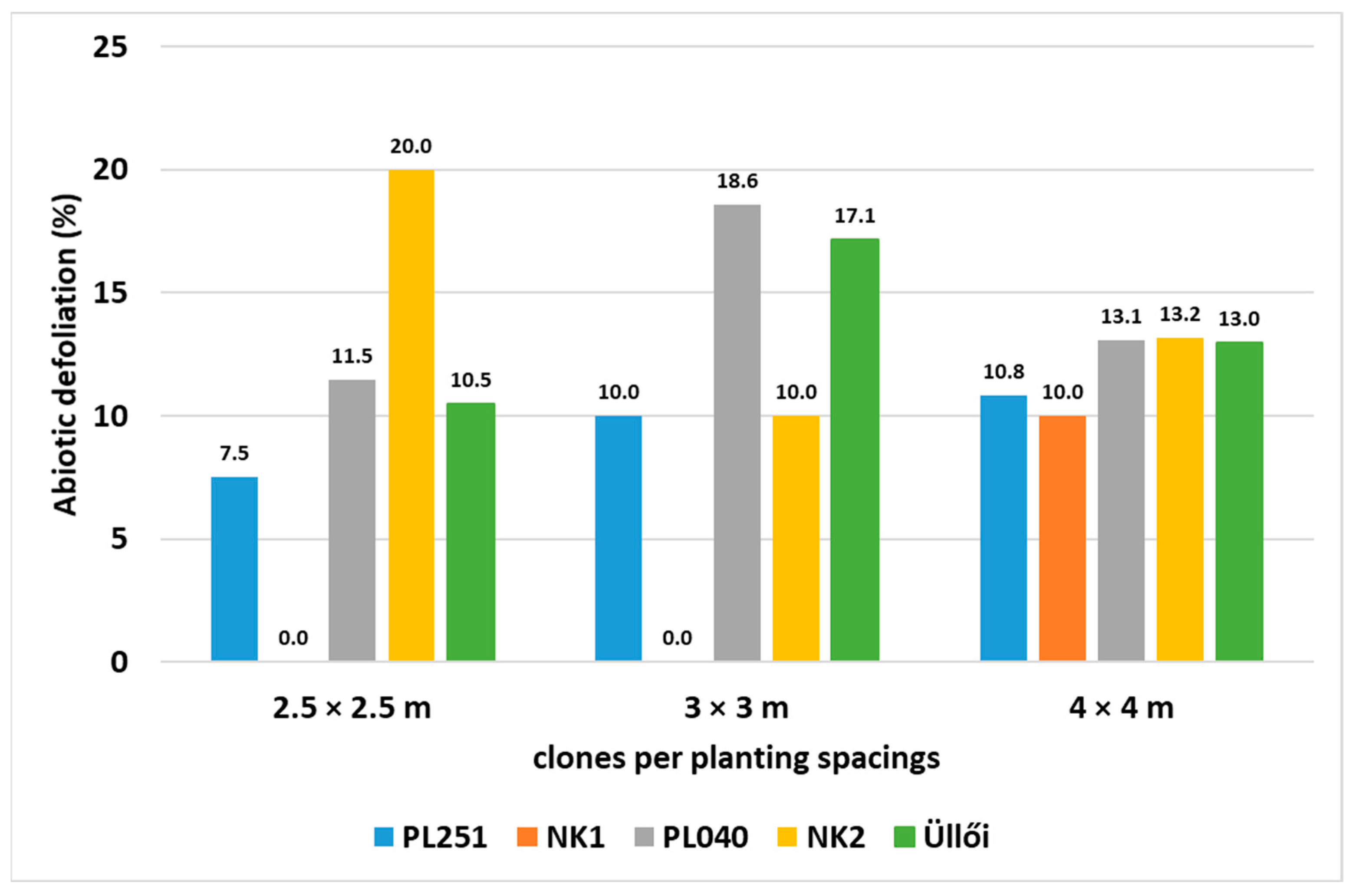
Abiotic defoliation of 30 randomly selected sample trees/clones was also investigated at the end of the main growing period (May–August) in the month of September. The test results showed low percentages for all clones, with clone NK1 highlighted at 0%.
We can improve the potential and resistance of this tree species to cope with extreme climates in a generative way, such as through human-assisted seedlot transfer from preadapted stands already in drier and warmer locations (e.g., southeastern Europe, e.g., Bulgaria and Turkey) or to establish a seed orchard from genotypes from these climatic conditions by grafting or using rooted cuttings and to exploit this seed stock; or by vegetative methods, for example, mass propagation of more drought-tolerant genotypes by vegetative means (tissue culture or micropropagation, establishment of a stool bed, production of rooting cuttings) based on ecophysiological studies.
5. Conclusions
The growth and physiological performance of different black locust (Robinia pseudoacacia L.) clones showed notable differences in response to the 2022 extremely dry season.
Of the clones previously considered the most promising, NK2 proved to be the best in terms of physiological parameters (CE, WUE, gtc, leaf temperature).
PL251, considered the second-best, performed poorly in physiological and growth parameters.
The clone PL040 seems promising under water stress conditions, as it showed the best thickness growth from May to August and performed well in drought tolerance parameters. Clone NK1 also showed good performance in height increment and abiotic stress tolerance.
The correlation between physiological parameters varied between clones, which could be attributed to genetic differences between clones. However, it should be pointed out that there was a strong positive correlation between A and Tr values for all clones.
We can conclude that it is possible to produce added value on marginal sites with selected black locust cultivars. Technology in growing and improving black locust is of increasing international importance, particularly in the face of the negative effects of global and local climate change. The additional cost of selective planting material can be compensated by the value-added tree plantations at a given age, or planting seed orchards of black locust clones could be a solution to reduce these costs.
Our findings can be adapted to any region potentially exposed to extreme climatic conditions, and clones of different species are expected to respond differently to abiotic stress, especially drought.



 ,
, 















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}