

Regulation of 5-Aminolevunilic Acid and Its Application in Agroforestry
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Natural ALA Occurrence
3. Physicochemical Properties and Toxicity of ALA
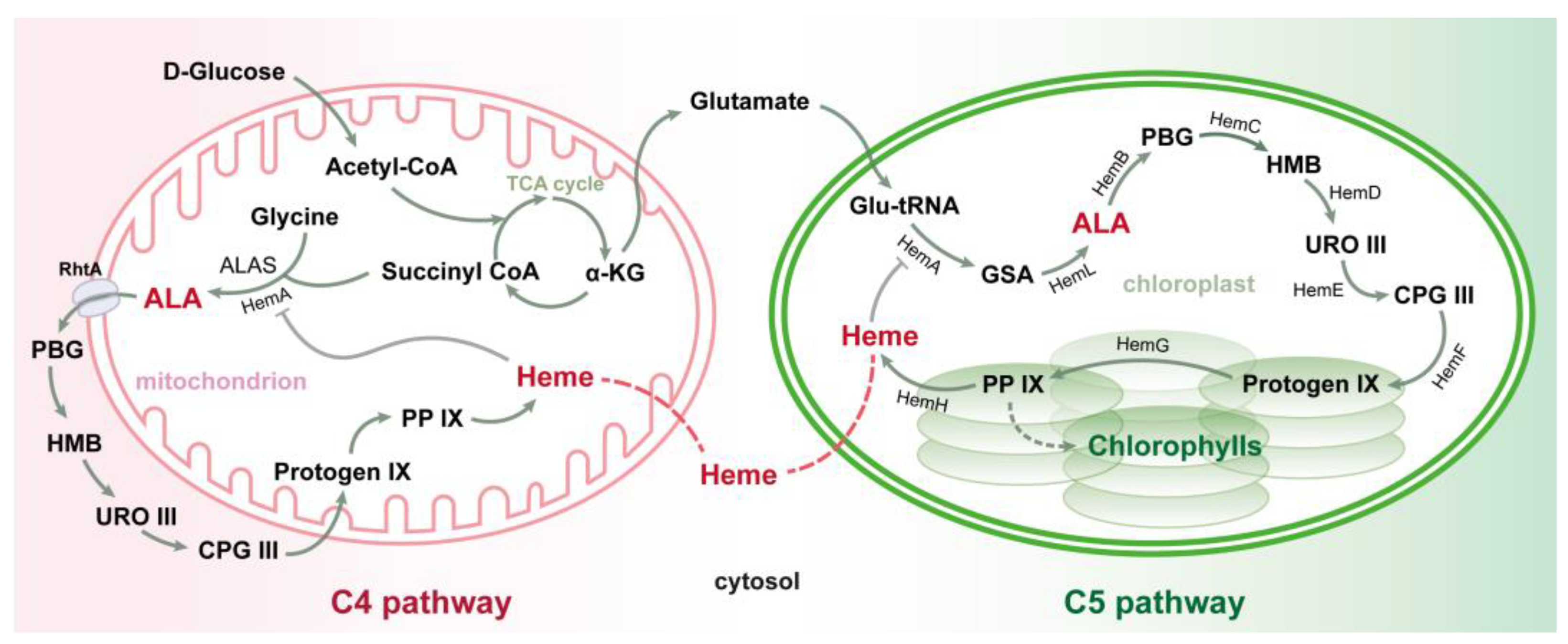
4. ALA Biosynthesis and Biological Production
4.1. The Pathways of ALA Biosynthesis
4.2. Biological Production of ALA
5. Multiple Applications of ALA in Agroforestry
5.1. As an Herbicide or a Toxicide of Herbicides
5.2. As an Insecticide or a Toxicide of Insecticides
5.3. Bidirectional Regulation on Probiotics and Pathogens
5.4. Plant Growth Substance
5.4.1. Application in Plant Tissue Culture
5.4.2. Promotion of Seed Germination
5.4.3. Improvement of Plant Growth and Crop Yield
5.4.4. Improvement of Plant Stress Tolerance
Low Temperature Stress
High Temperature Stress
Light Stresses
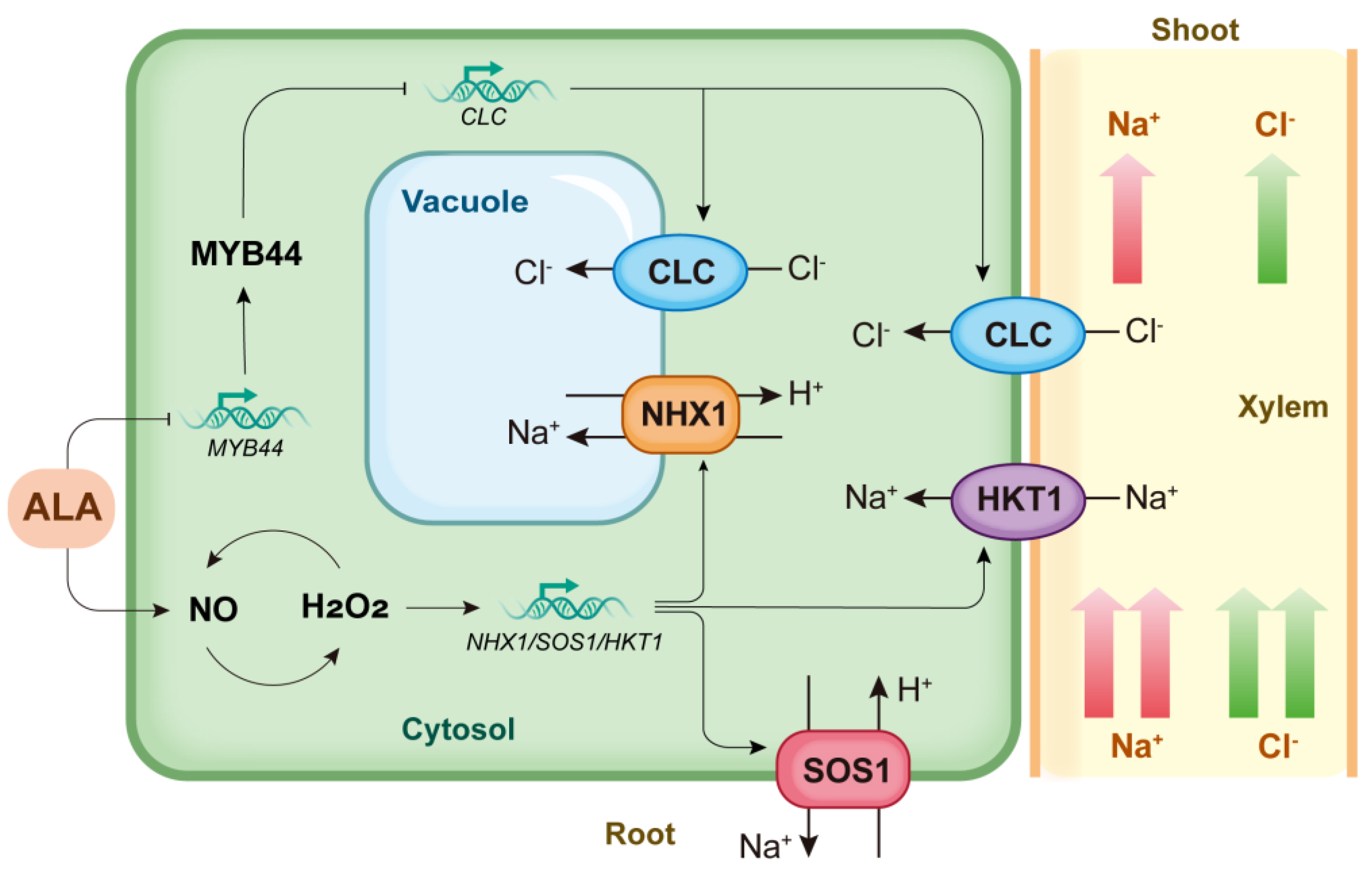
Salt Tolerance
Drought Stress
Waterlogging Stress
Toxic Metal Stresses
Other Environmental Pollution Stresses
Possible Mechanisms for ALA to Improve Stress Tolerance
Antioxidant Systems
Photosynthesis and Stomatal Regulation under Stressful Conditions
ALA-induced Ion Interception in Roots to Improve Salt Tolerance
ALA-induced Root Water Absorption to Increase Drought Tolerance
5.4.5. Delaying Senescence
5.4.6. Promoting Fruit Coloration and Flavonoid Biosynthesis

5.4.7. As a Thinning Agent to Remove Excess Flowers in Orchards
5.4.8. Promoting Nitrogen Absorption and Conversion with Less Nitrate Accumulation in Plant Tissues
6. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Rebeiz, C.A. (Ed.) Chlorophyll Biosynthesis and Technological Applications; Springer: Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; New York, NY, USA; London, UK, 2014; pp. 359–440. [Google Scholar]
- Hotta, Y.; Tanaka, H.; Takaoka, Y.; Takeuchi, Y.; Konnai, M. Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant Growth Regul. 1997, 22, 109–114. [Google Scholar] [CrossRef]
- Hotta, Y.; Tanaka, T.; Takaoka, H.; Takeuchi, Y.; Konnai, M. New physiological effects of 5-aminolevulinic acid in plants: The increase of photosynthesis, chlorophyll content, and plant growth. Biosci. Biotech. Biochem. 1997, 61, 2025–2028. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Jiang, W.B.; Liu, H.; Liu, W.Q.; Kang, L.; Hou, X.L. Promotion by 5-aminolevulinic acid of germination of pakchoi (Brassica campestris ssp. chinese var. communis Tsen et Lee) seeds under salt stress. J. Intergr. Plant Biol. 2005, 47, 1084–1091. [Google Scholar] [CrossRef]
- Dadario, N.B.; Khatri, D.; Reichman, N.; Nwagwu, C.D.; D’Amico, R.S. 5-Aminolevulinic acid- shedding light on where to focus. World Neurosurg. 2021, 150, 9–16. [Google Scholar] [CrossRef]
- Hendawy, A.O.; Khattab, M.S.; Sugimura, S.; Sato, K. Effects of 5-aminolevulinic acid as a supplement on animal performance, iron status, and immune response in farm animals: A review. Animals 2020, 10, 1352. [Google Scholar] [CrossRef]
- Wang, L.J.; Jiang, W.B.; Zhang, Z.; Yao, Q.H.; Matsui, H.; Ohara, H. Biosynthesis and physiological activities of 5-aminolevulinic acid (ALA) and its potential application in agriculture. Plant Physiol. Commun. 2003, 39, 185–192. [Google Scholar]
- Jahn, D.; Heinz, D.W. Biosynthesis of 5-aminolevulinic acid. In Tetrapyrroles: Birth, Lift and Death; Warren, M.J., Smith, A.G., Eds.; Landes Bioscience; Springer Science+Business Media: New York, NY, USA, 2009; pp. 29–42. [Google Scholar]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. In The Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2011; Volume 9, pp. 1–40. [Google Scholar] [CrossRef]
- Akram, N.A.; Ashraf, M. Regulation in plant stress tolerance by a potential plant growth regulator, 5-aminolevulinic acid. J. Plant Growth Regul. 2013, 32, 663–679. [Google Scholar] [CrossRef]
- Wu, Y.; Liao, W.B.; Dawuda, M.M.; Hu, L.L.; Yu, J.H. 5-Aminolevulinic acid (ALA) biosynthetic and metabolic pathways and its role in higher plants: A review. Plant Growth Regul. 2019, 87, 357–374. [Google Scholar] [CrossRef]
- Tan, S.Y.; Cao, J.; Xia, X.L.; Li, Z.H. Advances in 5-aminolevulinic acid priming to enhance plant tolerance to abiotic stress. Int. J. Mol. Sci. 2022, 23, 702. [Google Scholar] [CrossRef]
- Adachi, Y.; Umeda, M.; Kawazoe, A.; Sato, T.; Ohkawa, Y.; Kitajima, S.; Izawa, S.; Sagami, I.; Taketani, S. The novel heme-dependent inducible protein, SRRD regulates heme biosynthesis and circadian rhythms. Arch. Biochem. Biophys. 2017, 631, 19–29. [Google Scholar] [CrossRef]
- Zhang, J.L.; Kang, Z.; Chen, J.; Du, G.C. Optimization of the heme biosynthesis pathway for the production of 5-aminolevulinic acid in Escherichia coli. Sci. Rep. 2015, 5, 8584. [Google Scholar] [CrossRef] [PubMed]
- McCormac, A.C.; Fischer, A.; Kumar, A.M.; Soll, D.; Terry, M.J. Regulation of HEMA1 expression by phytochrome and a plastid signal during de-etiolation in Arabidopsis thaliana. Plant J. 2001, 25, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Gilles, H.; Jaenehen, R.; Thauer, R.K. Biosynthesis of 5-aminolevulinic acid in Methanobacterium thermoautotrophicum. Arch. Microbiol. 1983, 135, 237–240. [Google Scholar] [CrossRef]
- Nishikawa, S.; Watanabe, K.; Tanaka, T.; Miyachi, N. Rhodobacter sphaeroides mutants which accumulate 5-aminolevulinic acid under aerobic and dark conditions. J. Biosci. Bioengin. 1999, 87, 798–804. [Google Scholar] [CrossRef]
- Chen, J.Z.; Wang, Y.; Pu, W.; Zheng, P.; Sun, J.B. Advances and perspective on bioproduction of 5-aminolevulinic acid. Synthetic Biol. J. 2021, 2, 1000–1016. [Google Scholar]
- Pu, W.; Chen, J.Z.; Zhou, Y.Y.; Qiu, H.M.; Shi, T.; Zhou, W.J.; Guo, X.; Cai, N.Y.; Tan, Z.J.; Liu, J.; et al. Systems metabolic engineering of Escherichia coli for hyper-production of 5-aminolevulinic acid. Biotechnol. Biofuels Bioprod. 2023, 16, 31. [Google Scholar] [CrossRef]
- Van der Werf, M.; Zeikus, J.G. 5-Aminolevulinate production by Escherichia coli containing the Rhodobacter sphaeroides hemA gene. Appl. Environ. Microbiol. 1996, 62, 3560–3566. [Google Scholar] [CrossRef]
- Kang, Z.; Wang, Y.; Gu, P.F.; Wang, Q.; Qi, Q.S. Engineering Escherichia coli for efficient production of 5-aminolevulinic acid from glucose. Metab. Eng. 2011, 13, 492–498. [Google Scholar] [CrossRef]
- Shang, K.; Guo, X.F.; Wang, Y.P.; Zheng, P. Influence of 5-aminolevulinic acid dehydratase deletion on E. coli growth. Mod. Food Sci. Technol. 2011, 27, 742–746. [Google Scholar]
- Xue, C.F.; Yu, T.H.; Ng, I.S. Engineering pyridoxal kinase PdxY-integrated Escherichia coli strain and optimization for high-level 5-aminolevulinic acid production. J. Taiwan Inst. Chem. Eng. 2021, 120, 49–58. [Google Scholar] [CrossRef]
- Zhang, B.; Ye, B.C. Pathway engineering in Corynebacterium glutamicum S9114 for 5-aminolevulinic acid production. 3 Biotech 2018, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.J.; You, S.K.; Kim, M.; Lee, E.; Shin, S.K.; Park, H.M.; Oh, Y.; Han, S.O. Enhanced production of 5-aminolevulinic acid via flux redistribution of TCA cycle toward l-glutamate in Corynebacterium glutamicum. Biotechnol. Bioproc. Eng. 2019, 24, 915–923. [Google Scholar] [CrossRef]
- Cui, Z.Y.; Zhu, Z.W.; Zhang, J.H.; Jiang, Z.N.; Liu, Y.H.; Wang, Q.; Hou, J.; Qi, Q.S. Efficient 5-aminolevulinic acid production through reconstructing the metabolic pathway in SDH-deficient Yarrowia lipolytica. Biochem. Eng. J. 2021, 174, 108125. [Google Scholar] [CrossRef]
- Ge, F.L.; Li, X.K.; Ge, Q.R.; Zhu, D.; Li, W.; Shi, F.H.; Chen, H.J. Modular control of multiple pathways of Corynebacterium glutamicum for 5-aminolevulinic acid production. AMB Express 2021, 11, 179. [Google Scholar] [CrossRef]
- Ting, W.W.; Ng, I.S. Adaptive laboratory evolution and metabolic regulation of genetic Escherichia coli W3110 toward low-carbon footprint production of 5-aminolevulinic acid. J. Taiwan Inst. Chem. Eng. 2022, 141, 104612. [Google Scholar] [CrossRef]
- Yu, T.H.; Tan, S.I.; Yi, Y.C.; Xue, C.F.; Ting, W.W.; Chang, J.J.; Ng, I.S. New insight into the codon usage and medium optimization toward stable and high-level 5-aminolevulinic acid production in Escherichia coli. Biochem. Eng. J. 2022, 177, 108259. [Google Scholar] [CrossRef]
- He, G.M.; Jiang, M.R.; Cui, Z.Z.; Sun, X.; Chen, T.; Wang, Z.W. Construction of 5-aminolevulinic acid synthase variants by cysteine-targeted mutation to release heme inhibition. J. Biosci. Bioeng. 2022, 134, 416–423. [Google Scholar] [CrossRef]
- Yu, X.; Cao, Y.J.; Chen, Y.N.; Qi, Z.G.; Chen, Q.; Wang, J.C.; Kong, Q.S. Efficient production of 5-aminolevulinic acid from low-cost and sustainable bioresources of corn by engineered Corynebacterium glutamicum. Biores. Technol. Rep. 2022, 20, 101277. [Google Scholar] [CrossRef]
- Rebeiz, C.A.; Montazer-Zouhool, A.; Hopen, H.; Wu, S.M. Photodynamic herbicides I: Concepts and phenomenology. Enzym. Microb. Technol. 1984, 6, 390–401. [Google Scholar] [CrossRef]
- Yan, H.T.; Wang, B.F.; Li, H.J. Studies on the synergist of photoactivation pesticide δ-aminolevulinic acid. Bull. Sci. Technol. 1995, 11, 228–231. [Google Scholar]
- Chon, S.U. Herbicidal activity of δ-aminolevulinic acid on several plants as affected by application methods. Kor. J. Crop Sci. 2003, 48, 50–55. [Google Scholar]
- Phung, T.H.; Jung, S.Y. Differential antioxidant defense and detoxification mechanisms in photodynamically stressed rice plants treated with the deregulators of porphyrin biosynthesis, 5-aminolevulinic acid and oxyfluorfen. BBRC 2015, 459, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Phung, T.H.; Jung, S.Y. Perturbed porphyrin biosynthesis contributes to differential herbicidal symptoms in photodynamically stressed rice (Oryza sativa) treated with 5-aminolevulinic acid and oxyfluorfen. Pest. Biochem. Physiol. 2014, 116, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.F.; Zhang, F.; Jin, Z.L.; Huang, C.P.; Tang, G.X.; Ye, Q.F.; Zhou, W.J. ALA improving Brassica napus seedling tolerance to herbicide stress. J. Nucl. Agric. Sci. 2008, 22, 488–494. [Google Scholar]
- Xu, L.; Zhang, W.F.; Ali, B.; Islam, F.; Zhu, J.W.; Zhou, W.J. Synergism of herbicide toxicity by 5-aminolevulinic acid is related to physiological and ultra-structural disorders in crickweed (Malachium aquaticum L.). Pest. Biochem. Physiol. 2015, 125, 53–61. [Google Scholar] [CrossRef]
- Xu, L.; Islam, F.; Zhang, W.F.; Ghani, M.A.; Ali, B. 5-Aminolevulinic acid alleviates herbicide-induced physiological and ultrastructural changes in Brassica napus. J. Integr. Agric. 2018, 17, 579–592. [Google Scholar] [CrossRef]
- Sun, H.L.; Hou, Y.L.; Mei, Y.M.; Hao, P.; Wang, X.Q.; Lyu, D.G. Role of NADPH oxidase-mediated hydrogen peroxide in 5-aminolevulinic acid induced photooxidative stress tolerance in pear leaves. Sci. Hortic. 2022, 294, 110771. [Google Scholar] [CrossRef]
- Rebeiz, C.A.; Juvik, J.A.; Rebeiz, C.C. Porphyric insecticides I: Concept and phenomenology. Pestic. Biochem. Physiol. 1988, 30, 11–27. [Google Scholar] [CrossRef]
- Li, Y.M.; Guo, Y.P.; Li, Q.; Zheng, H.L.; Ma, E.B. Effect of porphyrin insecticides agent on Oxya chinensis and mechanisms. J. Shanxi Univ. Nat. Sci. Ed. 2005, 28, 196–201. [Google Scholar]
- Yin, K.; Ma, E.B.; Xue, C.R.; Wu, H.H.; Guo, Y.P.; Zhang, J.Z. Insecticidal activities of 5-aminolevulinic acid on Oxya chinensis and effect on three kinds of enzymes. Sci. Agric. Sin. 2008, 41, 2003–2007. [Google Scholar] [CrossRef]
- Yang, M.L.; Yin, K.; Guo, Y.P.; Ma, E.B.; Zhang, J.Z. A photosensitivity insecticide, 5-aminolevulinic acid, exerts effective toxicity to Oxya chinensis (Orthoptera: Acridoidea). Agric. Sci. China 2011, 10, 1056–1063. [Google Scholar] [CrossRef]
- Cheng, F.X. Toxicity on Nematodes and Mechanism of 5-Aminolevulinic Acid Produced by Photosynthetic Bacteria. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2011. [Google Scholar]
- Taspinar, M.S.; Aydin, M.; Arslan, E.; Yaprak, M.; Agar, G. 5-Aminolevulinic acid improves DNA damage and DNA methylation changes in deltamethrin-exposed Phaseolus vulgaris seedlings. Plant Physiol. Biochem. 2017, 118, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Szocs, K.; Gabor, F.; Csik, G.; Fidy, J.; Fidy, J. Delta-aminolaevulinic acid-induced porphyrin synthesis and photodynamic inactivation of Escherichia coli B. J. Photochem. Photobiol. B Biol. 1999, 50, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Lukšienė, Ž.; Danilčenko, H.; Tarasevičienė, Ž.; Anusevičius, Ž.; Marozienė, A.; Nivinskas, H. New approach to the fungal decontamination of wheat used for wheat sprouts: Effects of aminolevulinic acid. Intern. J. Food Microbiol. 2007, 116, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, J.T.; Zhang, H.W.; Cao, R.X.; Tang, D.L.; Wang, L.J. 5-Aminolevulinic acid against strawberry fusarium wilt: Bidirectional regulation of biocontrol agents and pathogens. Hortic. Plant J. 2023, in press. [Google Scholar]
- Bindu, R.C.; Vivekanandan, M. Hormonal activities of 5-aminolevulinic acid in callus induction and micropropagation. Plant Growth Regul. 1998, 26, 15–18. [Google Scholar] [CrossRef]
- Tabuchi, K.; Mizuta, H.; Yasui, H. Promotion of callus propagation by 5-aminolevulinic acid in a Laminaria japonica sporophyte. Aquac. Res. 2009, 41, 1–10. [Google Scholar] [CrossRef]
- Awad, M.A. Promotive effects of a 5-aminolevulinic acid-based fertilizer on growth of tissue culture- derived date palm plants (Phoenix dactylifera L.) during acclimatization. Sci. Hortic. 2008, 118, 48–52. [Google Scholar] [CrossRef]
- Nahar, S.J.; Haque, S.M.; Kazuhiko, S. Effect of 5-aminolevulinic acid on PLB culture of Cymbidium dayanum in vitro. Plant Tissue Cult. Biotech. 2015, 25, 99–102. [Google Scholar] [CrossRef]
- Nahar, S.J.; Himasaki, K.S. Application of 5-aminolevulinic acid for the in vitro micropropagation of cymbidium as a potential novel plant regulator. Environ. Control Biol. 2014, 52, 117–121. [Google Scholar] [CrossRef]
- Ling, L.J.; Wang, M.; Pan, C.Q.; Tang, D.B.; Yuan, E.; Zhang, Y.Y.; Chen, J.G.; Peng, D.Y.; Yin, Z.P. Investigating the induction of polyphenol biosynthesis in the cultured Cycolocarya paliurus cells and the stimulatory mechanism of co-induction with 5-aminolevulinic acid and salicylic acid. Front. Bioeng. Biotechnol. 2023, 11, 1150842. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kang, L.; Wang, L.J. Promotion of 5-aminolevunlinic acid on seed germination of watermelon (Citrullus lanatus) under salt stress. J. Fruit Sci. 2006, 23, 854–859. [Google Scholar]
- Ghanbari, F.; Zarei, B.; Pour-Aboughadareh, A.; Kordi, S. Investigation of 5-aminolevolinic acid (ALA) effects on seed germination and seedling growth of Silybum marianum under salinity stress. Intern. J. Biosci. 2013, 3, 95–101. [Google Scholar]
- Zhang, C.P.; He, P.; Yuan, F.G.; Hu, S.J.; Wei, P.X. Effect of exogenous ALA on seed germination and physiological characteristics of Glycyrrhiza uralensis seedlings under drought stress. Acta Bot. Boreal-Occident. Sin. 2011, 31, 1603–1610. [Google Scholar]
- Li, S.; Li, Z.S.; Miao, L.H.; Shang, L.R.; Zhou, X.X.; Nie, Z.N.; Li, X.L.; Wan, L.Q. Effect of 5-ALA on seed germination of alfalfa under drought stress. Grassl. Turf. 2021, 41, 92–98. [Google Scholar]
- Sheteiwy, M.; Shen, H.Q.; Xu, J.G.; Guan, Y.J.; Song, W.J.; Hu, J. Seed polyamines metabolism induced by seed priming with spermidine and 5-aminolevulinic acid for chilling tolerance improvement in rice (Oryza sativa L.) seedlings. Environ. Exp. Bot. 2017, 137, 58–72. [Google Scholar] [CrossRef]
- Kanto, U.; Jutamanee, K.; Osotsapar, Y.; Chai-arree, W.; Jattupornpong, S. Promotive effect of priming with 5-aminolevulinic acid on seed germination capacity, seedling growth and antioxidant enzyme activity in rice subjected to accelerated ageing treatment. Plant Prod. Sci. 2015, 18, 443–454. [Google Scholar] [CrossRef]
- Gupta, D.; Prasad, S.M. Priming with 5-aminolevulinic acid (ALA) attenuates UV-B induced damaging effects in two varieties of Cajanus cajan L. seedlings by regulating photosynthetic and antioxidant systems. S. Afric. J. Bot. 2021, 138, 129–140. [Google Scholar] [CrossRef]
- Lou, B.S.; Hotta, Y.; Qu, Y.L.; Zhao, J.S.; Tanaka, T.; Takeuchi, Y.; Konnai, M. Effects of 5-aminolevulinic acid on the growth and ripening of wheat. J. Pestic. Sci. 1998, 23, 300–303. [Google Scholar] [CrossRef]
- Wang, L.J.; Shi, W.; Liu, H.; Liu, W.Q.; Jiang, W.B.; Hou, X.L. Effects of exogenous 5-aminolevulinic acid treatment on leaf photosynthesis of pak-choi. J. Nanjing Agric. Univ. 2004, 27, 34–38. [Google Scholar]
- Wang, L.J.; Liu, W.Q.; Sun, G.R.; Wang, J.B.; Jiang, W.B.; Liu, H.; Li, Z.Q.; Zhuang, M. Effects of 5-aminolevulinic acid on photosynthesis and chlorophyll fluorescence of radish seedlings. Acta Bot. Boreal-Occident. Sin. 2005, 25, 488–496. [Google Scholar]
- Sheng, Y.F.; Yang, L.L.; Wang, J.K.; Chen, Y. Effect of 5-aminolevulinic acid on the yield and benefit of carrot. Hortic. Seed 2020, 40, 8–9, 11. [Google Scholar]
- Xu, C.; Liu, S.G.; Liu, H.J.; Yin, B.Q.; Ma, Y.F.; Zhang, M. Effects of 5-aminolevulinic acid on carrot growth and quality under tape for direct sowing mode. J. Agric. 2023, 13, 33–36. [Google Scholar]
- Li, M.A.; Ma, L.; Hao, Q.; An, Y.Y.; Wang, L.J. Effect of 5-aminolevulinic acid on leaf photosynthetic characteristics, yield and quality of potato. China Veg. 2020, 11, 43–52. [Google Scholar]
- Al-Thabet, S.S. Promotive effect of 5-aminolevulinic acid on growth and yield of wheat grown under dry conditions. J. Agron. 2006, 5, 45–49. [Google Scholar] [CrossRef]
- Al-Khateeb, S.A. Promotive effect of 5-aminolevulinic acid on growth, yield and gas exchange capacity of barley (Hordeum vulgare L.) grown under different irrigation regimes. J. King Saud Univ. 2006, 18, 103–111. [Google Scholar]
- Al-Khateeb, A.A.; Al-khateeb, S.A.; Okawara, R.; Al-abdoulhady, I.A. Promotive effects of 5-aminolevulinic acid (5-ALA) fruit yield and quality of date palm cv., Khalas. J. Biol. Sci. 2006, 6, 1118–1121. [Google Scholar]
- Yao, S.M.; Wang, W.J.; Chen, G.X. Effects of 5-aminolevulinic acid on phosphate absorption and distribution in rice using 32P tracer method. PIant Nutr. Fertiliz. Sci. 2006, 12, 70–75. [Google Scholar]
- Yang, W.P.; Wang, C.H.; Li, H.C. Effects of foliage spray with ALA on photosynthesis characteristic and yield of Yujing 6. Guangdong Agric. Sci. 2011, 36–37. [Google Scholar] [CrossRef]
- Xu, M.; Xu, F.L. Effects of exogenous 5-aminolevulinic acid on the growth and the yield of the tomato in sunlight greenhouse. J. Northwest A&F Univ. Nat. Sci. Ed. 2008, 6, 128–132. [Google Scholar]
- Feng, Z.W.; Guo, S.; Mao, L.P.; Zhao, C.F. Effect of ALA treatment to quality and yield in spinach. J. Shanxi Agric. Sci. 2009, 37, 42–43. [Google Scholar]
- Guo, Z.; Xu, F.L.; Wang, Y.K. Effects of 5-aminolevulinic acid on the growth, the yield and quality of compact jujube in mountainous region. J. Northwest Forest. Univ. 2010, 25, 93–96. [Google Scholar]
- Zheng, Q.Q.; Chen, Q.L.; Li, M.; Wang, J.J.; Zhi, J.H. Effects of compound plant growth regulator on fruit growth and development, yield and quality of Jun-jujube. Guizhou Agric. Sci. 2015, 43, 131–134. [Google Scholar]
- Duan, C.H.; Shen, M.; Zhang, Z.P.; Zhang, C.; Sun, L.C.; Ruan, S.C.; Wang, L.J. Effects of amino-acid fertilizer on leaf photosynthesis and yield of soybean. J. Nanjing Agric. Univ. 2012, 35, 15–20. [Google Scholar]
- Cheng, X.H.; Feng, X.X.; Zhang, Z.P.; Shen, M.; Wang, L.J. Effects of “Alstrong” amino-acid fertilizer on photosynthetic efficiency and yield of strawberry in plastic tunnels. J. Fruit Sci. 2012, 29, 883–889. [Google Scholar]
- Kong, L.G.; Wang, Y.H.; Hang, X.D.; Bo, H.J.; Sun, J.; Wang, L.J. Effect of Cropspring foliar fertilizer on growth and quality of rice under different nitrogen supplies. Jiangsu J. Agric. Sci. 2018, 34, 790–798. [Google Scholar]
- Tang, H.H.; Xu, Y.L.; Wang, Q.Y.; Ma, Z.B.; Li, G.Y.; Dong, H.; Dong, Z.Q. Effects of foliar spraying 5-aminolevulinic acid on spring maize growth and yield under different planting densities. Crops 2019, 2, 136–141. [Google Scholar]
- Liang, R.L.; Wang, L.J.; Wang, X.Q.; Zhang, J.T.; Gan, X. Effects of exogenous ALA on leaf photosynthesis, photosynthate transport, and sugar accumulation in Prunus persica L. Forests 2023, 14, 723. [Google Scholar] [CrossRef]
- An, Y.Y.; Cheng, D.X.; Rao, Z.X.; Sun, Y.P.; Tang, Q.; Wang, L.J. 5-Aminolevulinic acid (ALA) promotes primary root elongation through modulation of auxin transport in Arabidopsis. Acta Physiol. Plant. 2019, 41, 85. [Google Scholar] [CrossRef]
- Belda-Palazón, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-González, E. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef]
- Zhang, Q.; Yuan, W.; Wang, Q.W.; Cao, Y.Y.; Xu, F.Y.; Dodd, I.C.; Xu, W.F. ABA regulation of root growth during soil drying and recovery can involve auxin response. Plant Cell Environ. 2021, 45, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Rao, Z.X.; An, Y.Y.; Cao, R.X.; Tang, Q.; Wang, L.J. Study on possible mechanisms of ALA alleviating ABA inhibiting root growth of strawberry. Acta Hortic. Sin. 2023, 50, 461–474. [Google Scholar]
- Hotta, Y.; Tanaka, T.; Bingshan, L.; Takeuchi, Y.; Konnai, M. Improvement of cold resistance in rice seedlings by 5-aminolevulinic acid. J. Pest. Sci. 1998, 23, 29–33. [Google Scholar] [CrossRef]
- Hotta, Y.; Tanaka, T.; Konnai, M.; Takeuchi, Y. Effects of 5-aminolevulinc acid on the growth of manilagrass and bentgrass. Turfgrass Res. 2000, 28, 97–102. [Google Scholar]
- Wang, L.J.; Jiang, W.B.; Huang, B.J. Promotion of 5-aminolevulinic acid on photosynthesis of melon seedlings under low light and chilling stress conditions. Physiol. Plant. 2004, 121, 258–264. [Google Scholar] [CrossRef]
- Kang, L.; Cheng, Y.; Wang, L.J. Effects of 5-aminolevulinic acid (ALA) on the photosynthesis and anti-oxidative enzymes activities of the leaves of greenhouse watermelon in autumn and winter. Acta Bot. Boreal-Occident. Sin. 2006, 26, 2297–2301. [Google Scholar]
- Su, C.H.; Zhong, X.L.; Wang, D.L.; Li, M.S. Effects of different chemicals on resistance of tomato seedling to chilling under low light. J. Nat. Disasters 2006, 15, 312–317. [Google Scholar]
- Yin, L.L.; Yu, X.C.; Wang, Y.H.; Xu, Z.H.; Li, K.; Han, D.J. Effect of 5-aminolevulinic acid on chilling tolerance in cucumber seedlings. Acta Agric. Boreali-Occident. Sin. 2007, 16, 166–169. [Google Scholar]
- Korkmaz, A.; Korkmaz, Y.; Demirkıran, A.R. Enhancing chilling stress tolerance of pepper seedlings by exogenous application of 5-aminolevulinic acid. Environ. Exp. Bot. 2010, 67, 495–501. [Google Scholar] [CrossRef]
- Korkmaz, A.; Korkmaz, Y. Promotion by 5-aminolevulenic acid of pepper seed germination and seedling emergence under low-temperature stress. Sci. Hortic. 2009, 119, 98–102. [Google Scholar] [CrossRef]
- Wang, H.P.; Liu, Z.C.; Luo, S.L.; Li, J.; Zhang, J.; Li, L.S.; Xie, J.M. 5-Aminolevulinic acid and hydrogen sulphide alleviate chilling stress in pepper (Capsicum annuum L.) seedlings by enhancing chlorophyll synthesis pathway. Plant Physiol. Biochem. 2021, 167, 567–576. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Tomaro, M.L.; Batlle, A.; Noriega, G.O. The role of 5-aminolevulinic acid in the response to cold stress in soybean plants. Phytochemistry 2010, 71, 2038–2045. [Google Scholar] [CrossRef]
- Manafi, E.; Sanavy, S.A.M.M.; Aghaalikhani, M.; Dolatabadian, A. Exogenous 5-aminolevulenic acid promotes antioxidative defence system, photosynthesis and growth in soybean against cold stress. Not. Sci. Biol. 2015, 7, 486–494. [Google Scholar] [CrossRef]
- Mao, L.P.; Ren, J.; Zhang, X.H. Effects of ALA on photosynthetic characteristics of summer squash seedlings under chilling stress. Chin. Agric. Sci. Bull. 2011, 27, 142–145. [Google Scholar]
- Huang, F.; Li, M.F.; Wang, L.J.; Li, S.P.; He, J. Mitigative effects of foliage spraying and root irrigation of ALA on banana plantlets exposed to cold stress. Agric. Sci. Technol. 2013, 14, 858–862. [Google Scholar]
- Liu, H.; Huang, J. Effects of 5-aminolevulinic acid on chilling tolerance of eggplant seedlings. J. Changjiang Veg. 2014, 43–46. [Google Scholar]
- Zhang, Z.P.; Zhang, L.L. Effect of 5-aminolevulinic acid on cold tolerance and antioxidant systems of young seedlings of rape (Brassica napus L.). Jiangsu Agric. Sci. 2014, 42, 52–55. [Google Scholar]
- Fu, J.J.; Chu, X.T.; Sun, Y.F.; Xu, Y.F.; Hu, T.M. Involvement of nitric oxide in 5-aminolevulinic acid-induced antioxidant defense in roots of Elymus nutans exposed to cold stress. Biol. Plant. 2016, 60, 585–594. [Google Scholar] [CrossRef]
- Chen, H.; Xu, L.; Li, X.; Wang, D.Y.; An, Y.Y.; Wang, L.J. Promotive effect of 5-aminolevulinic acid on freezing tolerance of Rhododendron simsii and Cinnamomum camphora leaves. Plant Physiol. J. 2017, 53, 2103–2113. [Google Scholar]
- Zhang, Z.D.; Yuan, L.Q.; Ma, Y.B.; Kang, Z.; Zhou, F.; Gao, Y.; Yang, S.C.; Li, T.L.; Hu, X.H. Exogenous 5-aminolevulinic acid alleviates low-temperature damage by modulating the xanthophyll cycle and nutrient uptake in tomato seedlings. Plant Physiol. Biochem. 2022, 189, 83–93. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Zhang, Y.H.; Yuan, L.Q.; Zhou, F.; Gao, Y.; Kang, Z.; Li, T.L.; Hu, X.H. Exogenous 5-aminolevulinic acid alleviates low-temperature injury by regulating glutathione metabolism and β-alanine metabolism in tomato seedling roots. Ecotoxicol. Environ. Saf. 2022, 245, 114112. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Q.; Han, Z.X.; Lai, J.X.; Yang, S.H.; Luo, G.H. Effect of 5-aminolevulinic acid on fast chlorophyll fluorescence induction kinetics of Scenedesmus obliquus under different temperature conditions. Acta Bot. Boreal-Occident. Sin. 2018, 38, 932–938. [Google Scholar]
- Anwar, A.; Yan, Y.; Liu, Y.M.; Li, Y.S.; Yu, X.C. 5-Aminolevulinic acid improves nutrient uptake and endogenous hormone accumulation, enhancing low-temperature stress tolerance in cucumbers. Int. J. Mol. Sci. 2018, 19, 3379. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Ai, X.Z.; Yu, X.C. Effects of ALA on photosynthesis of cucumber seedlings under suboptimal temperature and light intensity. Acta Hortic. Sin. 2010, 37, 65–71. [Google Scholar]
- Jiang, W.H.; Yan, F.; Fu, J.; Qu, D.; Feng, Z.L. Effect of exogenous 5-aminolevulinic acid on photosynthetic characteristics of tea plants under low temperature stress. J. Northwest A&F Univ. Nat. Sci. Ed. 2020, 48, 108–115. [Google Scholar]
- Yan, F.; Jiang, W.H.; Qu, D.; Fu, J.; Zhao, X. Effects of exogenous 5-aminolevulinic acid on photosynthetic and physiological characteristics of tea plants under low temperature stress. J. Tea Sci. 2020, 40, 597–606. [Google Scholar]
- Yan, F.; Qu, D.; Chen, X.H.; Zeng, H.T.; Li, X.S.; Hu, C.Y. Metabolomics reveals 5-aminolevulinic acid improved the ability of tea leaves (Camellia sinensis L.) against cold stress. Metabolites 2022, 12, 392. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Nie, G.W.; Li, K.; Zhang, X.P.; Dai, L.R. Ameliorating effects of 5-aminolevulinic acid on damage to sweet cherry floral organ under low temperature stress. Acta Agric. Boreal-Occident. Sin. 2020, 29, 595–602. [Google Scholar]
- Zhang, J.T.; Wei, X.; Zhang, H.W.; Liang, R.L.; An, Y.Y.; Wang, L.J. Mitigative effect of 5-aminolevulinic acid pretreatment on chilling injury of young leaves in peach. J. Fruit Sci. 2022, 39, 845–855. [Google Scholar]
- Liu, F.; Guo, J.J.; Gong, L.L.; Xu, X.M. Increasing resistance of rice to low temperature photoinhibition by spraying 5-aminolevulinic acid was associated with activities of anti-oxidant enzymes. Chin. J. Rice Sci. 2008, 22, 411–415. [Google Scholar]
- Liu, T.; Xu, J.J.; Li, J.M.; Hu, X.H. NO is involved in JA- and H2O2-mediated ALA-induced oxidative stress tolerance at low temperatures in tomato. Environ. Exp. Bot. 2019, 161, 334–343. [Google Scholar] [CrossRef]
- Sun, Y.P. Study on the Mechanism of Photosynthesis and Photoinhibition of 5-Aminolevulinic Acid (ALA) in Watermelon Seedlings under Stress Conditions. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2009. [Google Scholar]
- Al-Qurashi, A.D.; Awad, M.A. 5-Aminolevulinc acid increases tree yield and improves fruit quality of ‘Rabia’ and ‘Sukkariat-Yanbo’ date palm cultivars under hot arid climate. Sci. Hortic. 2011, 129, 441–448. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.M.; Gao, Y.; Yu, B.; Xia, C.X.; Bai, J.G. Pretreatment with 5-aminolevulinic acid mitigates heat stress of cucumber leaves. Biol. Plant. 2012, 56, 780–784. [Google Scholar] [CrossRef]
- Ma, N.; Qi, L.; Gao, J.J.; Chao, K.C.; Hu, Q.F.; Jiang, H.G.; Wang, L.J. Effects of 5-aminolevulinic acid on cutting growth under high temperature condition and leaf chlorophyll fast fluorescence characteristics of Ficus carica L. J. Nanjing Agric. Univ. 2015, 38, 546–553. [Google Scholar]
- Wang, C.K.; Guo, M. Effect of 5-aminolevulinic acid on growth and antioxidant enzyme activities of Avena nuda under normal and high temperature conditions. J. Henan Agric. Sci. 2017, 46, 30–34. [Google Scholar]
- Li, X.L.; Hua, Z.R.; Zhang, D.T. Induction effect of 5-aminoevulinic acid (ALA) on heat resistance in Qinling alpine rhododendrons. Jiangsu Agric. Sci. 2017, 45, 176–179. [Google Scholar]
- Wang, D.Y.; Li, X.; Xu, L.; An, Y.Y.; Wang, L.J. Effect of 5-aminolevulinic acid on leaf heat tolerance in Ligustrum Japonicum and Spiraea japonica. Bot. Res. 2018, 7, 350–365. [Google Scholar]
- Li, W.X.; Wang, S.M.; Zhao, Y. Effect of spraying ALA on heat resistance of Medicago sativa. Acta Agrestia Sin. 2019, 27, 1188–1194. [Google Scholar]
- Wang, L.J.; Sun, Y.P.; Kang, L.; Zhang, Z.P. Effects of 5-aminolevulinic acid (ALA) on photosynthesis and chlorophyll fluorescence of watermelon seedlings grown under low light and low temperature conditions. Acta Hortic. 2010, 856, 159–166. [Google Scholar] [CrossRef]
- Sun, Y.P.; Wang, L.J. Effects of 5-aminolevulinic acid (ALA) on chlorophyll fluorescence dynamics of watermelon seedlings under shade condition. Acta Hortic. Sin. 2007, 34, 901–908. [Google Scholar]
- Sun, Y.P.; Zhang, Z.P.; Wang, L.J. Promotion of 5-aminolevulinic acid treatment on leaf photosynthesis is related with increase of antioxidant enzyme activity in watermelon seedlings grown under shade condition. Photosynthetica 2009, 47, 347–354. [Google Scholar] [CrossRef]
- Guo, X.Q.; Li, C.H.; Li, Q.Z.; Li, X.; Li, Y.S.; Yu, X.C. Effects of foliar spraying 5-aminolevulinic acid on growth, photosynthesis and yield of tomato under shade conditions. Shandong Agric. Sci. 2011, 9, 30–34. [Google Scholar]
- Tang, X.Q.; Wang, K.C.; Xiao, Y.H.; Wang, L.J. Effects of 5-aminolevulinic acid (ALA) on the growth, leaf stomatal gas exchange, and leaf alkaloids content of Isatis indigotica under shading condition. Chin. J. Ecol. 2013, 32, 1155–1160. [Google Scholar]
- An, Y.Y.; Zhang, L.Y.; Feng, X.X.; Tian, F.; Li, J.; Wang, L.J. Effect of 5-aminolevulinic acid on low light tolerance of apple leaves. Acta Bot. Boreal-Occident. Sin. 2016, 36, 987–995. [Google Scholar]
- Anwar, A.; Wang, J.; Yu, X.C.; He, C.X.; Li, Y.S. Substrate application of 5-aminolevulinic acid enhanced low-temperature and weak-light stress tolerance in cucumber (Cucumis sativus L.). Agronomy 2020, 10, 472. [Google Scholar] [CrossRef]
- Liu, W.Q.; Kang, L.; Wang, L.J. Effects of ALA on strawberry photosynthesis and relations to anti-oxidant enzymes. Acta Bot. Boreal-Occident. Sin. 2006, 26, 57–62. [Google Scholar]
- Kang, L.; Wang, L.J. Effects of ALA treatments on light response curves of chlorophyll fluorescence of watermelon leaves. J. Nanjing Agric. Univ. 2008, 31, 31–36. [Google Scholar]
- Aksakal, O.; Algur, O.F.; Aksakal, F.I.; Aysin, F. Exogenous 5-aminolevulinic acid alleviates the detrimental effects of UV-B stress on lettuce (Lactuca sativa L.) seedlings. Acta Physiol. Plant. 2017, 39, 55. [Google Scholar] [CrossRef]
- Watanabe, K.; Tanaka, T.; Hotta, Y. Improving salt tolerance of cotton seedlings with 5-aminolevulinic acid. Plant Growth Regul. 2000, 32, 99–103. [Google Scholar] [CrossRef]
- Nishihara, E.; Kondo, K.; Parvez, M.M.; Takahashi, K.; Watanabe, K.; Tanaka, K. Role of 5-aminolevulinic acid (ALA) on active oxygen-scavenging system in NaCl-treated spinach (Spinacia oleracea). J. Plant Physiol. 2003, 160, 1085–1091. [Google Scholar] [CrossRef]
- Zhou, H.F.; Zou, Z.R.; Meng, C.J.; Guo, H.T.; Yang, X.J. The effect of exogenous ALA, CaCl2 and SA on physiological characteristics of muskmelon seedling under salt stress. Agric. Res. Arid. Areas 2007, 25, 212–215. [Google Scholar]
- Wang, N.J.; Kang, B.W.; Liu, J.J.; Xi, S.H. Effect of ALA on seedling and planting in salina soil. J. Northwest A&F Univ. Nat. Sci. Ed. 2008, 36, 76–80. [Google Scholar]
- Qi, X.L.; Zou, Z.R.; Yang, R. Mitigative Effect of 5-aminolevulinic acid on lettuce under NaCl stress. Acta Agric. Boreal-Occident. Sin. 2008, 17, 202–206. [Google Scholar]
- Youssef, T.; Awad, M.A. Mechanisms of enhancing photosynthetic gas exchange in date palm seedlings (Phoenix dactylifera L.) under salinity stress by a 5-aminolevulinic acid-based fertilizer. J. Plant Growth Regul. 2008, 27, 1–9. [Google Scholar] [CrossRef]
- Zhao, X.J.; Li, Y.F.; Mai, W.X.; Tian, X.H.; Lai, H.X.; Lv, J.L. A study on promotion of ALA on resistance of wheat to saline soil and amendment of saline-alkali soil through addition of sulfur. Acta Agric. Boreal-Occident. Sin. 2008, 17, 303–308. [Google Scholar]
- Gao, N.C.; Sun, Y.P.; Zhang, Q.; Shao, H.P.; Zhu, J.Y. Effect of exogenous 5-aminolevulinic acid (ALA) on photosynthesis of NaCl-stressed strawberry plants. Jiangsu J. Agric. Sci. 2010, 26, 1329–1333. [Google Scholar]
- Naeem, M.S.; Jin, Z.L.; Wan, G.L.; Liu, D.; Liu, H.B.; Yoneyama, K.; Zhou, W.J. 5-Aminolevulinic acid improves photosynthetic gas exchange capacity and ion uptake under salinity stress in oilseed rape (Brassica napus L.). Plant Soil 2010, 332, 405–415. [Google Scholar] [CrossRef]
- Zhang, C.P.; He, P.; Wei, P.X.; Du, D.D.; Yu, Z.L. Effect of exogenous 5-aminolevulinic acid on seed germination and antioxidase activities of Perilla frutescens seedlings under NaCl stress. Chin. Trad. Herb. Drug. 2011, 42, 1194–1200. [Google Scholar]
- Zhang, C.P.; He, P.; Yu, Z.; Du, D.D.; Wei, P.X. Effect of exogenous Ca2+, ALA, SA and Spd on seed germination and physiological characteristics of Perilla frutescens seedlings under NaCl stress. Chin. J. Chin. Mater. Medica 2010, 35, 3260–3265. [Google Scholar]
- Akram, N.A.; Ashraf, M.; Al-Qurainy, F. Aminolevulinic acid-induced changes in some key physiological attributes and activities of antioxidant enzymes in sunflower (Helianthus annuus L.) plants under saline regimes. Sci. Hortic. 2012, 142, 143–148. [Google Scholar] [CrossRef]
- Zhen, A.; Bie, Z.L.; Huang, Y.; Liu, Z.X.; Fan, M.L. Effects of 5-aminolevulinic acid on the H2O2-content and antioxidative enzyme gene expression in NaCl-treated cucumber seedlings. Biol. Plant. 2012, 56, 566–570. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, N.; Hu, L.L.; Liao, W.B.; Tang, Z.Q.; Xiao, X.M.; Lyu, J.; Xie, J.M.; Calderón-Urrea, A.; Yu, J.H. 5-Aminolevulinic acid improves morphogenesis and Na+ subcellular distribution in the apical cells of Cucumis sativus L. under salinity stress. Front. Plant. Sci. 2021, 12, 636121. [Google Scholar] [CrossRef]
- Liu, P.; Zhao, B.L.; Wang, W.J.; Sun, J.L.; Ma, H.X.; Zhang, X. Study on mitigative effect of 5-aminolevulinic acid on grape under salt stress. North. Hortic. 2014, 9, 19–22. [Google Scholar]
- Nunkaew, T.; Kantachote, D.; Kanzaki, H.; Nitoda, T.; Ritchie, R. Effects of 5-aminolevulinic acid (ALA)-containing supernatants from selected Rhodopseudomonas palustris strains on rice growth under NaCl stress, with mediating effects on chlorophyll, photosynthesis electron transport and antioxidative enzymes. Electronic. J. Biotechnol. 2014, 17, 19–26. [Google Scholar] [CrossRef]
- Liu, L.Y.; Nguyen, N.T.; Ueda, A.; Saneoka, H. Effects of 5-aminolevulinic acid on Swiss chard (Beta vulgaris L. subsp. cicla) seedling growth under saline conditions. Plant Growth Regul. 2014, 74, 219–228. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Miao, M.M.; Wang, C.L. Effects of ALA on photosynthesis, antioxidant enzyme activity, and gene expression, and regulation of proline accumulation in tomato seedlings under NaCl stress. J. Plant Growth Regul. 2015, 34, 637–650. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Yan, F.; Hu, L.P.; Zhou, X.T.; Zou, Z.R.; Cu, L.R. Effects of exogenous 5-aminolevulinic acid on photosynthesis, stomatal conductance, transpiration rate, and PIP gene expression of tomato seedlings subject to salinity stress. Gene Mol. Res. 2015, 14, 6401–6412. [Google Scholar] [CrossRef]
- Freije, A.; Saleh, K.; Islam, S.; Al-Mannai, M. The mechanism behind the promotive effect of foliar application of 5-aminolevulinic acid (ALA) in tomato plants under salt stress. J. Adv. Agric. 2018, 8, 1415–1423. [Google Scholar] [CrossRef]
- Ye, J.B.; Chen, Q.W.; Tao, T.T.; Wang, G.Y.; Xu, F. Promotive effects of 5-aminolevulinic acid on growth, photosynthetic gas exchange, chlorophyll, and antioxidative enzymes under salinity stress in Prunnus persica (L.) Batseh seedling. Emirates J. Food Agric. 2016, 28, 786–795. [Google Scholar] [CrossRef]
- Anjum, S.A.; Li, J.H.; Lv, J.; Zong, X.F.; Wang, L.; Yang, A.J.; Yan, R.; Ali, Z.; Song, J.X.; Wang, S.G. Regulation mechanism of exogenous ALA on growth and physiology of Leymus chinensis (Trin.) under salt stress. Chil. J. Agric. Res. 2016, 76, 314–320. [Google Scholar] [CrossRef]
- Wu, X.H.; Feng, J.M. Effects of seed soaking with different concentrations of 5-aminolevulinic acid on the germination and salt tolerance of pumpkin seeds under NaCl stress. Seed 2016, 35, 90–93. [Google Scholar]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Biochem. 2018, 129, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Guo, S.; Wei, Y.; Zhang, C.P.; Wu, N.B. Effects of exogenous 5-aminolevulinic acid on physiological characteristics and secondary metabolite contents of Atropa belladonna L. seedlings under NaCl stress. J. Southwest Univ. Nat. Sci. Ed. 2018, 9, 13–20. [Google Scholar]
- Fan, X.L.; Yang, Y.L.; Ren, J.; Gao, Y.; Li, A.; Gao, Y.; Li, H. Effects of exogenous 5-aminolevulinic acid on physiological characteristics of cauliflower seedlings under salt stress. Tianjin Agric. Sci. 2019, 12, 1–4. [Google Scholar]
- Chang, X.Y.; Sun, J.L.; Zhao, B.L.; Li, F.F.; Liu, L.L.; He, W. Effects of exogenous ALA on photosynthesis and membrane peroxidation in the leaves of jujube seedlings under NaCl treatment. Xinjiang Agric. Sci. 2019, 56, 1635–1644. [Google Scholar]
- Chen, L.L.; Sun, J.L.; Chang, X.Y.; Ye, J.F. Effects of exogenous ALA on chlorophyll synthesis in jujube seedlings under NaCl stress. Xinjiang Agric. Sci. 2022, 59, 1659–1665. [Google Scholar]
- Yang, S.; Zhao, L.Y.; Yan, J.M.; Zhang, J.L.; Guo, F.; Geng, Y.; Wang, Q.; Yang, F.Y.; Wan, S.B.; Li, X.G. Peanut genes encoding tetrapyrrole biosynthetic enzymes, AhHEMA1 and AhFC1, alleviating the salt stress in transgenic tobacco. Plant Physiol. Biochem. 2019, 137, 14–24. [Google Scholar] [CrossRef]
- Liu, L.Y.; El-Shemy, H.A.; Saneoka, H. Effects of 5-aminolevulinic acid on water uptake, ionic toxicity, and antioxidant capacity of Swiss chard (Beta vulgaris L.) under sodic-alkaline conditions. J. Plant Nutr. Soil Sci. 2017, 180, 535–543. [Google Scholar] [CrossRef]
- Zhang, J.X.; Guo, Y.F.; Zhang, M. Effects of 5-aminolevulinic acid on seed germination and seedling growth of maize under salt stress. J. Agric. 2021, 7, 7–12. [Google Scholar]
- Li, X.; Li, J.J.; Islam, F.; Najeeb, U.; Pan, J.M.; Huo, Z.N.; Shou, J.Y.; Qin, Y.B.; Xu, L. 5-Aminolevulinic acid could enhance the salinity tolerance by alleviating oxidative damages in Salvia miltiorrhiza. Food Sci. Technol. 2022, 42, e103121. [Google Scholar] [CrossRef]
- Brilli, F.; Pignattelli, S.; Baraldi, R.; Neri, L.; Pollastri, S.; Gonnelli, C.; Giovannelli, A.; Loreto, F.; Cocozza, C. Root exposure to 5-aminolevulinic acid (ALA) affects leaf element accumulation, isoprene emission, phytohormonal balance, and photosynthesis of salt-stressed Arundo donax. Int. J. Mol. Sci. 2022, 23, 4311. [Google Scholar] [CrossRef] [PubMed]
- Nunkaew, T.; Kantachote, D.; Nitoda, T.; Kanzaki, H. Selection of salt tolerant purple nonsulfur bacteria producing 5-aminolevulinic acid (ALA) and reducing methane emissions from microbial rice straw degradation. Appl. Soil Ecol. 2015, 86, 113–120. [Google Scholar] [CrossRef]
- Abdelhadi, A.W.; Salih, A.A.; Sultan, K.; Alsafi, A.; Tashtoush, F. Actual water use of young date palm trees as affected by aminolevulinic acid application and different irrigation water salinities. Irrig. Drain. 2020, 69, 427–440. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Yao, Q.H.; Wang, L.J. Expression of yeast Hem1 controlled by Arabidopsis HemA1 promoter enhances leaf photosynthesis in transgenic tobacco. Mol. Biol. Rep. 2011, 38, 4369–4379. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Yang, K.; Lee, D.E.; Back, K. Expression of Bradyrhizobium japonicum 5-aminolevulinic acid synthase induces severe photodynamic damage in transgenic rice. Plant Sci. 2004, 167, 789–795. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Yao, Q.H.; Wang, L.J. Expression of yeast Hem1 gene controlled by Arabidopsis HemA1 promoter improves salt tolerance in Arabidopsis plants. BMB Rep. 2010, 43, 330–336. [Google Scholar] [CrossRef]
- Sun, X.E.; Feng, X.X.; Li, C.; Zhang, Z.P.; Wang, L.J. Study on salt tolerance with YHem1 transgenic canola (Brassica napus). Physiol. Plant. 2015, 154, 223–242. [Google Scholar] [CrossRef]
- Lu, J.C.; Mou, B.M.; Zheng, D.F.; Feng, N.J.; Liang, X.Y.; Feng, S.J.; Xiang, H.T. Effects of 5-aminolevulinic acid on biomass and physiological characteristics of soybean seedlings under saline-alkali stress. Soybean Sci. 2020, 39, 84–89. [Google Scholar]
- Liu, H.; Wang, Z.; Dong, S.; Zou, J.X.; Jin, H. Mitigation effect of exogenous 5-aminolevulinic acid on Populus wutunensis seedling under saline-alkali stress. J. Northeast Forest. Univ. 2022, 7, 47–52. [Google Scholar]
- Cheng, J.E.; Xiao, Q.M.; Cheng, F.X.; Liu, Y. Effect of 5-aminolevulinic acid on photosynthesis of greenhouse tobacco and its stress tolerance. Hunan Agric. Sci. 2007, 4, 58–60. [Google Scholar]
- Li, D.M.; Zhang, J.; Sun, W.J.; Li, Q.; Dai, A.H.; Bai, J.G. 5-Aminolevulinic acid pretreatment mitigates drought stress of cucumber leaves through altering antioxidant enzyme activity. Sci. Hortic. 2011, 130, 820–828. [Google Scholar] [CrossRef]
- Liu, D.; Pei, Z.F.; Naeem, M.S.; Ming, D.F.; Liu, H.B.; Khan, F.; Zhou, W.J. 5-Aminolevulinic acid activates antioxidative defence system and seedling growth in Brassica napus L. under water-deficit stress. J. Agron. Crop. Sci. 2011, 197, 284–295. [Google Scholar] [CrossRef]
- Liu, D.; Hu, L.Y.; Ali, B.; Yang, A.G.; Wan, G.L.; Xu, L.; Zhou, W.L. Influence of 5-aminolevulinic acid on photosynthetically related parameters and gene expression in Brassica napus L. under drought stress. Soil Sci. Plant Nutri. 2016, 62, 254–262. [Google Scholar] [CrossRef]
- Akram, N.A.; Iqbal, M.; Muhammad, A.; Ashraf, M.; Al-Qurainy, F.; Shafiq, S. Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. Protoplasma 2018, 255, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.X.; He, P.; Zhang, C.P.; Xie, Y.Z.; Liu, H.Y.; Duan, C.X.; Hu, B. Effects of exogenous 5-aminolevulinic acid (ALA) and nitric oxide donor sodium nitroprusside (SNP) on the physiological characteristics of Fagopyrum dibotrys seedlings under drought stress. J. Southwest China Norm. Univ. Nat. Sci. Ed. 2012, 10, 52–58. [Google Scholar]
- Zhang, C.P.; He, P.; Yuan, F.G.; Yu, Z.L.; Du, D.D.; Wei, P.X.; Hu, S.J. Effects of exogenous 5-aminolevulinic acid on chlorophyll fluorescence characteristics and energy dissipation of Sarcandra glabra under drought stress. Chin. Trad. Herb. Drug. 2012, 43, 64–172. [Google Scholar]
- Kosar, F.; Akram, N.A.; Ashraf, M. Exogenously-applied 5-aminolevulinic acid modulates some key physiological characteristics and antioxidative defense system in spring wheat (Triticum aestivum L.) seedlings under water stress. South Afric. J. Bot. 2015, 96, 71–77. [Google Scholar] [CrossRef]
- Akram, N.A.; Kausar, S.; Farid, N.; Ashraf, M.; Al-Qurainy, F. 5-Aminolevulinic acid induces regulation in growth, yield and physio biochemical characteristics of wheat under water stress. Sains Malays. 2018, 47, 661–670. [Google Scholar] [CrossRef]
- Wang, Y.X.; Wei, S.M.; Wang, J.N.; Su, X.Y.; Suo, B.; Qin, F.J.; Zhao, H.J. Exogenous application of 5-aminolevulinic acid on wheat seedlings under drought stress enhances the transcription of psbA and psbD genes and improves photosynthesis. Brazil J. Bot. 2018, 41, 275–285. [Google Scholar] [CrossRef]
- Ostrowska, A.; Biesaga-Kościelniak, J.; Grzesiak, M.T.; Hura, T. Physiological responses of spring wheat to 5-aminolevulinic acid under water stress applied at seedling stage. Cereal Res. Commun. 2019, 47, 32–41. [Google Scholar] [CrossRef]
- Suliman, M.S.E.; Elradi, S.B.M.; Nimir, N.E.A.; Zhou, G.S.; Zhu, G.L.; Ibrahim, M.E.H.; Ali, A.Y.A. Foliar application of 5-aminolevulinic acid alleviated high temperature and drought stresses on wheat plants at seedling stage. Chil J. Agric. Res. 2021, 81, 291–299. [Google Scholar] [CrossRef]
- Chen, X.Y.; Song, Y.H.; Zhang, M.H.; Li, X.Y.; Li, H.; Wang, Y.X.; Qi, X.L. Effects of water deficit on physiology and biochemistry of seedlings of different wheat varieties and the alleviation effect of exogenous application of 5-aminolevulinic acid. Acta Agron. Sin. 2022, 48, 478–487. [Google Scholar] [CrossRef]
- Liu, M.R.; Li, J.H.; Niu, J.H.; Wang, R.; Song, J.X.; Lv, J.; Zong, X.F.; Wang, S.G. Interaction of drought and 5-aminolevulinic acid on growth and drought resistance of Leymus chinensis seedlings. Acta Ecol. Sin. 2016, 36, 180–188. [Google Scholar] [CrossRef]
- Ou, C.; Yao, X.M.; Yao, X.J.; Ji, J.; Wang, W.G.; Guo, J. Changes of photosynthetic and antioxidant properties of Gardenia jasmioides seedlings treated with 5-aminolevulinic acid and PEG. Agric. Res. Arid Area. 2016, 34, 235–241. [Google Scholar]
- Wang, H.X.; Ju, C.H.; Li, X.L.; Chen, B.; Leng, N. Effects of exogenous 5-aminolevulinic acid on growth and root physiological characteristics of Dalbergia odorifera T. Chen under drought stress. Chin. J. Trop. Crop. 2017, 38, 1823–1829. [Google Scholar]
- Niu, K.J.; Ma, H.L. The positive effects of exogenous 5-aminolevulinic acid on the chlorophyll biosynthesis photosystem and Calvin cycle of Kentucky bluegrass seedlings in response to osmotic stress. Environ. Exp. Bot. 2018, 155, 260–271. [Google Scholar] [CrossRef]
- Niu, K.J.; Ma, X.; Liang, G.L.; Ma, H.L.; Jia, Z.F.; Liu, W.H.; Yu, Q.Q. 5-Aminolevulinic acid modulates antioxidant defense systems and mitigates drought-induced damage in Kentucky bluegrass seedlings. Protoplasma 2017, 254, 2083–2094. [Google Scholar] [CrossRef]
- Alikhani-Koupaei, M.; Fatahi, R.; Zamani, Z.; Salimi, S. 5-Aminolevulinic acid moderates environmental stress- induced bunch wilting and stress markers in date palm. Acta Physiol. Plant. 2018, 40, 159. [Google Scholar] [CrossRef]
- Rasheed, R.; Yasmeen, H.; Hussain, I.; Iqbal, M.; Ashraf, M.A.; Parveen, A. Exogenously applied 5-aminolevulinic acid modulates growth, secondary metabolism and oxidative defense in sunflower under water deficit stress. Physiol. Mol. Biol. Plants 2020, 26, 489–499. [Google Scholar] [CrossRef]
- Wang, F.Z.; Jin, Y.N.; Li, Z.W.; Chen, B.; Xiong, Y.N.; Hao, H.H.; Xu, Z.H. Effects of exogenous ALA (5-aminolevulinic acid) on photosynthesis and antioxidant system of flue-cured tobacco seedlings under drought stress. Chin. Tob. Sci. 2020, 41, 22–29. [Google Scholar]
- Yang, A.J.; Chang, Q.L.; Wang, P.; Wang, F.; Gao, Y.T.; Zhou, G.K.; Song, X.J.; Wei, E.C. Effects of exogenous 5-aminolevulinic acid on seed germination and seedling growth of maize under drought stress. Crops 2022, 3, 194–199. [Google Scholar]
- Islam, M.R.; Naznin, T.; Gupta, D.R.; Haque, M.A.; Hasanuzzaman, M.; Rohman, M.M. Insight into 5-aminolevulinic acid-induced modulation of cellular antioxidant metabolism to confer salinity and drought tolerance in maize. Biocell 2020, 44, 713–730. [Google Scholar] [CrossRef]
- Jiao, Z.Y.; Han, S.; Yu, X.; Huang, M.B.; Lian, C.L.; Liu, C.; Yin, W.L.; Xia, X.L. 5-Aminolevulinic acid pretreatment mitigates drought and salt stresses in poplar plants. Forests 2021, 12, 1112. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.M.; Elsheery, N.I.; Kalaji, H.M.; Santos-Villalobos, S.D.; Wróbel, J.; Hassan, I.F.; Gaballah, M.S.; Abdelrhman, L.A.; Mira, A.M.; et al. 5-Aminolevulinic acid and 24-epibrassinolide improve the drought stress resilience and productivity of banana plants. Plants 2022, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.Y.; He, S.S.; An, Y.Y.; Wang, L.J. Exogenous 5-aminolevulinic acid improves strawberry tolerance to osmotic stress and its possible mechanisms. Physiol. Plant. 2020, 168, 948–962. [Google Scholar] [CrossRef]
- An, Y.Y.; Qi, L.; Wang, L.J. ALA pretreatment improves waterlogging tolerance of fig plants. PLoS ONE 2016, 11, e0147202. [Google Scholar] [CrossRef] [PubMed]
- Kriswantoro, H.; Lakitan, B.; Lesbani, A.; Wijaya, A. Foliar application of 5-aminolevulinic acid for offsetting unfavorable effects of shallow water table on growth and yield in snap bean (Phaseolus vulgaris). Bul. J. Agric. Sci. 2020, 26, 638–645. [Google Scholar]
- Noriega, G.O.; Balestrasse, K.B.; Batlle, A.; Tomaro, M.L. Cadmium induced oxidative stress in soybean plants also by the accumulation of δ-aminolevulinic acid. Biometals 2007, 20, 841–851. [Google Scholar] [CrossRef]
- Ali, B.; Tao, Q.J.; Zhou, Y.F.; Gill, R.A.; Ali, S.; Rafiq, M.T.; Xu, L.; Zhou, W.J. 5-Aminolevolinic acid mitigates the cadmium-induced changes in Brassica napus as revealed by the biochemical and ultra-structural evaluation of roots. Ecotoxicol. Environ. Safe 2013, 92, 271–280. [Google Scholar] [CrossRef]
- Ali, B.; Wang, B.; Ali, S.; Ghani, M.A.; Hayat, M.T.; Yang, C.; Xu, L.; Zhou, W.J. 5-Aminolevulinic acid ameliorates the growth, photosynthetic gas exchange capacity, and ultrastructural changes under cadmium stress in Brassica napus L. J. Plant Growth Regul. 2013, 32, 604–614. [Google Scholar] [CrossRef]
- Xu, L.; Ali, B.; Gill, R.A.; Li, L.; Zhou, W.J. Alleviation of cadmium toxicity by 5-aminolevulinic acid is related to improved nutrients uptake and lowered oxidative stress in Brassica napus. Int. J. Agric. Biol. 2016, 18, 557–564. [Google Scholar] [CrossRef]
- Yang, L.J.; Wu, Y.; Wang, X.M.; Lv, J.; Tang, Z.Q.; Hu, L.L.; Luo, S.L.; Wang, R.D.; Ali, B.; Yu, J.H. Physiological mechanism of exogenous 5-aminolevulinic acid improved the tolerance of Chinese cabbage (Brassica pekinensis L.) to cadmium stress. Front. Plant Sci. 2022, 13, 845396. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.E.; Li, X.; Najeeb, U.; Hou, Z.N.; Buttar, N.A.; Yang, Z.Q.; Ali, B.; Xu, L. Soil applied silicon and manganese combined with foliar application of 5-aminolevulinic acid mediate photosynthetic recovery in Cd-stressed Salvia miltiorrhiza by regulating Cd-transporter genes. Front. Plant Sci. 2022, 13, 1011872. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.J.; Zhang, Y.T.; Guo, Q.X.; Tan, H.F.; Han, J.H.; Lin, H.R.; Wei, H.W.; Xu, G.W.; Zhu, C. Effects of exogenous 5-aminolevulinic acid and 24-epibrassinolide on Cd accumulation in rice from Cd-contaminated soil. Rice Sci. 2018, 25, 320–329. [Google Scholar]
- Xu, L.; Li, J.J.; Najeeb, U.; Li, X.; Pan, J.M.; Huang, Q.; Zhou, W.J.; Liang, Z.S. Synergistic effects of EDDS and ALA on phytoextraction of cadmium as revealed by biochemical and ultrastructural changes in sunflower (Helianthus annuus L.) tissues. J. Haz. Mat. 2022, 407, 124764. [Google Scholar] [CrossRef]
- Gill, R.A.; Ali, B.; Islam, F.; Farooq, M.A.; Gill, M.B.; Mwamba, T.M.; Zhou, W.J. Physiological and molecular analyses of black and yellow seeded Brassica napus regulated by 5-aminolivulinic acid under chromium stress. Plant Physiol. Biochem. 2015, 94, 130–143. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Hannan, F.; Rizwan, M.; Iqbal, M.; Hassan, Z.; Akram, N.A.; Maqbool, S.; Abbas, F. Promotive role of 5-aminolevulinic acid on chromium-induced morphological, photosynthetic, and oxidative changes in cauliflower (Brassica oleracea botrytis L.). Environ. Sci. Pollut. Res. 2017, 24, 8814–8824. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Nasir, T.; Abbasi, G.H.; Rehmani, M.I.A.; Ata-Ul-Karim, S.T.; Bukhari, S.A.H.; et al. Phyto-management of chromium contaminated soils through sunflower under exogenously applied 5-aminolevulinic acid. Ecotoxicol. Environ. Saf. 2018, 151, 255–265. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Saeed, R.; Rizwan, M.; Asad, S.; Bukhari, H.; Abbasi, G.H.; Hussain, A.; Ali, B.; Zamir, M.S.I.; et al. Combined application of citric acid and 5-aminolevulinic acid improved biomass, photosynthesis and gas exchange attributes of sunflower (Helianthus annuus L.) grown on chromium contaminated soil. Intern. J. Phytoremediat. 2019, 21, 760–767. [Google Scholar] [CrossRef]
- Tian, T.; Ali, B.; Qin, Y.B.; Malik, Z.; Gill, R.A.; Ali, S.; Zhou, W.J. Alleviation of lead toxicity by 5-aminolevulinic acid is related to elevated growth, photosynthesis, and suppressed ultrastructural damages in oilseed rape. Biomed. Res. Int. 2014, 2014, 530642. [Google Scholar] [CrossRef]
- Ali, B.; Xu, X.; Gill, R.A.; Yang, S.; Ali, S.; Tahir, M.; Zhou, W.J. Promotive role of 5-aminolevulinic acid on mineral nutrients and antioxidative defense system under lead toxicity in Brassica napus. Ind. Crop. Prod. 2014, 52, 617–626. [Google Scholar] [CrossRef]
- Singh, R.; Kesavan, A.K.; Landi, M.; Kaur, S.; Thakur, S.; Zheng, B.S.; Bhardwaj, R.; Sharma, A. 5-Aminolevulinic acid regulates Krebs cycle, antioxidative system and gene expression in Brassica juncea L. to confer tolerance against lead toxicity. J. Biotechnol. 2020, 323, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Rinklebe, J.; Ahmad, P. Alleviation of arsenic toxicity in pepper plants by aminolevulinic acid and heme through modulating its sequestration and distribution within cell organelles. Environ. Pollut. 2023, 330, 121747. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.X.; Fang, X.; Chen, M.X.; Wang, L.Y.; Xia, J.X.; Wang, Z.C.; Fang, J.G.; Tran, L.P.; Shangguan, L.F. Copper stress in grapevine: Consequences, responses, and a novel mitigation strategy using 5-aminolevulinic acid. Environ. Pollut. 2022, 307, 119561. [Google Scholar] [CrossRef] [PubMed]
- Kai, T.; Ikeura, H.; Ayanokoji, A.; Tamaki, M. Effects of 5-aminolevulinic acid (ALA) on Zinnia hybrida growth and phytoremediation effects in oil-contaminated soil. J. Agric. Chem. Environ. 2020, 9, 97–106. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, J.T.; Zhang, H.W.; Xu, Y.; An, Y.Y.; Wang, L.J. Effect of 5-aminolevulinic acid (5-ALA) on leaf chlorophyll fast fluorescence characteristics and mineral element content of Buxus megistophylla grown along urban roadsides. Horticulturae 2021, 7, 95. [Google Scholar] [CrossRef]
- Nishihara, E.; Takahashi, K.; Nakata, N.; Tanaka, K.; Watanabe, K. Effect of 5-aminolevulinic acid (ALA) on photosynthetic rate, hydrogen peroxide content, antioxidant level and active oxygen-scavenging enzymes in spinach (Spinacia oleracea L.). J. Jpn. Soc. Hort. Sci. 2001, 70, 346–352. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, L.L.; Liao, W.B.; Dawuda, M.M.; Lyu, J.; Xie, J.M.; Feng, Z.; Calderón-Urrea, A.; Yu, J.H. Foliar application of 5-aminolevulinic acid (ALA) alleviates NaCl stress in cucumber (Cucumis sativus L.) seedlings through the enhancement of ascorbate-glutathione cycle. Sci. Hortic. 2019, 257, 108761. [Google Scholar] [CrossRef]
- Liu, T.; Du, Q.J.; Li, S.Z.; Yang, J.Y.; Li, X.J.; Xu, J.J.; Chen, P.X.; Li, J.M.; Hu, X.H. GSTU43 gene involved in ALA-regulated redox homeostasis, to maintain coordinated chlorophyll synthesis of tomato at low temperature. BMC Plant Biol. 2019, 19, 323. [Google Scholar] [CrossRef]
- Wu, W.W.; He, S.S.; An, Y.Y.; Cao, R.X.; Sun, Y.P.; Tang, Q.; Wang, L.J. Hydrogen peroxide as a mediator of 5-aminolevulinic acid-induced Na+ retention in roots for improving salt tolerance of strawberries. Physiol. Plant. 2019, 167, 5–20. [Google Scholar] [CrossRef]
- He, S.S.; Yang, H.; Cao, R.Q.; Tang, Q.; An, Y.Y.; Wang, L.J. 5-Aminolevulinic acid-induced salt tolerance in strawberry (cv. ‘Benihoppe’): Possible role of nitric oxide on interception of salt ions in roots. Sci. Hortic. 2022, 304, 111294. [Google Scholar] [CrossRef]
- Xiong, J.L.; Wang, H.C.; Tan, X.Y.; Zhang, C.L.; Naeem, M.S. 5-Aminolevulinic acid improves salt tolerance mediated by regulation of tetrapyrrole and proline metabolism in Brassica napus L. seedlings under NaCl stress. Plant Physiol. Biochem. 2018, 124, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Liu, C.; Wei, B.; Zhang, J.; An, Y.; Wang, L. Exogenous 5-aminolevulinic acid promotes osmotic stress tolerance of walnuts by modulating photosynthesis, osmotic adjustment and antioxidant systems. Forests 2023, 14, 1789. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, Z.P.; Wang, L.J. Effect of 5-aminolevulinic Acid (ALA) on leaf diurnal photosynthetic characteristics and antioxidant activity in pear (Pyrus pyrifolia Nakai). In Artificial Photosynthesis; Najafpour, M., Ed.; In Techopen: London, UK, 2012; pp. 239–256. [Google Scholar]
- An, Y.Y.; Liu, L.B.; Chen, L.H.; Wang, L.J. ALA inhibits ABA-induced stomatal closure via reducing H2O2 and Ca2+ levels in guard cells. Front. Plant Sci. 2016, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Liu, L.B.; An, Y.Y.; Zhang, Z.P.; Wang, L.J. Preliminary Studies on the possible mechanism underlying 5-aminolevulinic acid-induced stomatal opening in apple leaves. Acta Hortic. Sin. 2014, 41, 1965–1974. [Google Scholar]
- Hu, J.; An, Y.Y.; Cai, C.Y.; He, S.S.; Wang, L.J. Cytoplasmic pH is involved in 5-aminolevulinic acid (ALA)-induced stomatal opening in apple leaves. Acta Hortic. Sin. 2019, 46, 1869–1881. [Google Scholar]
- Hu, S.; An, Y.Y.; Wang, L.J. Ethylene is involved in the regulation of stomatal movement by ALA-ABA/dark in apple leaves. Acta Hortic. Sin. 2020, 47, 409–420. [Google Scholar]
- An, Y.Y.; Xiong, L.J.; Hu, S.; Wang, L.J. PP2A and microtubules function in 5-aminolevulinic acid-mediated H2O2 signaling in Arabidopsis guard cells. Physiol. Plant. 2020, 168, 709–724. [Google Scholar] [CrossRef]
- An, Y.Y.; Feng, X.X.; Liu, L.B.; Xiong, L.J.; Wang, L.J. ALA-induced flavonols accumulation in guard cells is involved in scavenging H2O2 and inhibiting stomatal closure in Arabidopsis cotyledons. Front. Plant Sci. 2016, 7, 1713. [Google Scholar] [CrossRef]
- Xiong, L.J.; An, Y.Y.; Wang, L.J. The role of microtubule skeleton and PP1/PP2A protein phosphatase in ALA-ABA regulating stomatal movement in apple leaves. Acta Hortic. Sin. 2018, 45, 2073–2088. [Google Scholar]
- Chen, Z.; An, Y.Y.; Wang, L.J. ALA reverses ABA-induced stomatal closure by modulating PP2AC and SnRK2.6 activity in apple leaves. Hortic. Res. 2023, 10, uhad067. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, L.J. ALA upregulates MdPTPA expression to increase the PP2A activity and promote stomatal opening in apple leaves. Plant Sci. 2022, 325, 111490. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lou, Y.Y.R.; Wang, L.J. MdDGK3-like as a negative regulator participates in ALA-induced PP2AC to promote stomatal opening in apple leaves. Hortic. Plant J. 2023, in press. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, J.T.; Wang, L.J. ALA induces stomatal opening through regulation among PTPA, PP2AC, and SnRK2.6. Front. Plant Sci. 2023, 14, 1206728. [Google Scholar] [CrossRef]
- Wang, T.; Rao, J.P.; Song, Y.L.; Zhang, H.Y.; Zou, Z.R. Effects of 5-aminolevulinic acid on the quality and postharvest physiology of tomato fruit. J. Northwest A&F Univ. Nat. Sci. Ed. 2008, 10, 127–131. [Google Scholar]
- Song, Y.L.; Rao, J.P.; Wang, T.; Wang, L.L. Effects of ALA on post-harvest quality and storability of spinach. Acta Agric. Boreal-Occident. Sin. 2008, 17, 258–262. [Google Scholar]
- Wei, Z.Y.; Zhang, Z.P.; Lee, M.; Sun, Y.P.; Wang, L.J. Effect of 5-aminolevulinic acid on leaf senescence and nitrogen metabolism of pakchoi under different nitrate levels. J. Plant Nutri. 2010, 35, 49–63. [Google Scholar] [CrossRef]
- Niu, J.J.; Zhang, S.P.; Zhang, K.; Han, L.X.; Qu, Z.F.; Miao, J.Y. Effects of 5-aminolevulinic acid treatment on storage of Fuji apple during fruit expansion stage. J. Henan Agric. Sci. 2021, 10, 146–153. [Google Scholar]
- Wang, L.J.; Wang, Z.H.; Li, Z.Q.; Liu, H.; Liu, W.Q.; Chen, Z.Y.; Yan, P.; Sun, D.Q. Effect of 5-aminolevulinic acid on enhancing fruit coloration. J. Fruit Sci. 2004, 21, 512–515. [Google Scholar]
- Wang, Z.H.; Tang, G.H.; Li, Z.Q.; Wang, L.J. Promotion of 5-aminolevulinic acid and genistein on anthocyanin accumulation in apples. Acta Hortic. Sin. 2006, 33, 1055–1058. [Google Scholar]
- Feng, X.X.; An, Y.Y.; Zheng, J.; Sun, M.; Wang, L.J. Proteomics and SSH analyses of ALA-promoted fruit coloration and evidence for the involvement of a MADS-box gene, MdMADS1. Front. Plant Sci. 2016, 7, 1615. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhang, L.Z.; Wang, L.J. The transcription factor MdERF78 is involved in ALA-induced anthocyanin accumulation in apples. Front. Plant Sci. 2022, 13, 915197. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhang, L.Z.; Shangguan, L.F.; Wang, L.J. MdMYB110a, directly and indirectly, activates the structural genes for the ALA-induced accumulation of anthocyanin in apple. Plant Sci. 2023, 326, 111511. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, L.B.; Tao, H.H.; An, Y.Y.; Wang, L.J. Transcriptomic profiling of apple calli with a focus on the key genes for ALA-induced anthocyanin accumulation. Front. Plant Sci. 2021, 12, 640606. [Google Scholar] [CrossRef]
- Xie, L.; Wang, Z.H.; Cheng, X.H.; Gao, J.J.; Zhang, Z.P.; Wang, L.J. 5-Aminolevulinic acid promotes anthocyanin accumulation in Fuji apples. Plant Growth Regul. 2013, 69, 295–303. [Google Scholar] [CrossRef]
- Guo, L.; Cai, Z.X.; Zhang, B.B.; Xu, J.L.; Song, H.F.; Ma, R.J. The mechanism analysis of anthocyanin accumulation in peach accelerated by ALA. Acta Hortic. Sin. 2013, 40, 1043–1050. [Google Scholar]
- Xiao, C.C.; Zhang, S.L.; Hu, H.J.; Tian, R.; Wu, J.; Yang, Z.J.; Ma, C.Y. Effects of bagging and exogenous 5-aminolevulinic acid treatment on coloration of ‘Yunhongli 2’. J. Nanjing Agric. Univ. 2012, 6, 25–29. [Google Scholar]
- Liu, H.F.; Jia, Y.X.; Feng, X.X.; Li, L.L. Application of 5-aminolevulinic acid on Yuluxiang pear production. J. Shanxi Agric. Sci. 2022, 50, 155–160. [Google Scholar]
- Watanabe, K.; Nishihara, E.; Watanabe, S.; Tanaka, T.; Takahashi, K.; Takeuchi, Y. Enhancement of growth and fruit maturity in 2-year-old grapevines cv. Delaware by 5-aminolevulinic acid. Plant Growth Regul. 2006, 49, 35–42. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Sun, J.L.; Zhao, B.L.; Liu, J.J.; Zhang, S.; Li, G. Effects of exogenous ALA on fruit quality and PAL enzyme activity of grapevine. Sino-Overseas Grapevine Wine 2017, 3, 16–19. [Google Scholar]
- Xie, L.; Cheng, X.H.; Feng, X.X.; Yang, T.; Zhang, Z.P.; Wang, L.J. Effects of an amino acid fertilizer on the leaf photosynthesis and fruit quality of ‘Summer Black’ grape. J. Nanjing Agric. Univ. 2013, 2, 31–37. [Google Scholar]
- Liu, L.L.; Sun, J.L.; Zhao, B.L.; Chang, X.Y.; He, W. Effects of ALA solutions with different pH values on the coloration and quality of Crimson grape in solar greenhouse. J. Henan Agric. Sci. 2019, 5, 99–105. [Google Scholar]
- Feng, S.; Li, M.F.; Wu, F.; Li, W.L.; Li, S.P. 5-Aminolevulinic acid affects fruit coloration, growth, and nutrition quality of Litchi chinensis Sonn. cv. Feizixiao in Hainan, tropical China. Sci. Hortic. 2015, 193, 188–194. [Google Scholar] [CrossRef]
- Li, M.F.; Feng, S.; Li, S.P.; Wu, F.; Wang, F.; Li, C.L.; Fu, Y.N.; Bao, D.H.; Wang, L.J. Preharvest promotion or inhibition of colouration: Which is the more conducive to improving litchi postharvest quality? Sci. Hortic. 2019, 254, 124–132. [Google Scholar] [CrossRef]
- Liu, S.S.; Liu, Y.M.; He, H.T.; Lin, Z.Y.; Sun, J.; Zhang, F.X.; Zhou, L.L.; Wang, Z.B.; Zhang, Z.B.; Zou, H.S. Effects of 5-aminolevulinic acid (5-ALA) on physicochemical characteristics and growth of pomegranate (Punica granatum L.). Horticulturae 2023, 9, 860. [Google Scholar] [CrossRef]
- Meng, F.X.; Duan, Y.J.; Yang, Y.J.; Lu, Z.Y.; Liu, H.G. Effect of 5-aminoevulinic acid treatment on physicochemical properties and color development of mango fruit. S. China Fruits 2019, 3, 57–62. [Google Scholar]
- Averina, N.G.; Savina, S.M.; Dremuk, I.A.; Yemelyanava, H.V.; Pryshchepchyk, Y.V.; Usatov, A.V. Influence of 5-aminolevulinic acid on physiological and biochemical characteristics of winter wheat varieties with different levels of anthocyanins in coleoptiles. Proc. Natl. Acad. Sci. USA 2022, 2, 135–146. [Google Scholar] [CrossRef]
- Xu, F.; Cheng, S.Y.; Zhu, J.; Zhang, W.W.; Wang, Y. Effects of 5-aminolevulinic acid on chlorophyll, photosynthesis, soluble sugar and flavonoids of Ginkgo biloba. Not. Bot. Hort. Agrobot. Cluj. 2011, 39, 41–47. [Google Scholar] [CrossRef]
- Liu, L.B.; An, Y.Y.; Xiong, L.J.; Wang, L.J. Flavonols induced by 5-aminolevulinic acid are involved in regulation of stomatal opening in apple leaves. Acta Hortic. Sin. 2016, 43, 817–828. [Google Scholar] [CrossRef]
- Zheng, J.; An, Y.Y.; Feng, X.X.; Wang, L.J. Rhizospheric application with 5-aminolevulinic acid improves coloration and quality in ‘Fuji’ apples. Sci. Hortic. 2017, 224, 74–83. [Google Scholar] [CrossRef]
- Zhang, H.W.; Tao, H.H.; Yang, H.; Zhang, L.Z.; Feng, G.Z.; An, Y.Y.; Wang, L.J. MdSCL8 as a negative regulator participates in ALA-induced FLS1 to promote flavonol accumulation in apples. Int. J. Mol. Sci. 2022, 23, 2033. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Duan, C.H.; Zhang, Z.P.; Cheng, Y.; Wang, L.J.; Li, B.J. Effects of exogenous ALA on thinning and fruit quality in ‘Hosui’ pear (Pyrus pyrifolia). Acta Hortic. Sin. 2011, 38, 1515–1522. [Google Scholar]
- An, Y.Y.; Li, J.; Duan, C.H.; Liu, L.B.; Sun, Y.P.; Cao, R.X.; Wang, L.J. 5-Aminolevulinic acid thins pear fruits by inhibiting pollen tube growth via Ca2+-ATPase-mediated Ca2+ efflux. Front. Plant Sci. 2016, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- An, Y.Y.; Lu, W.Y.; Li, J.; Wang, L.J. ALA inhibits pear pollen tube growth through regulation of vesicle trafficking. Sci. Hortic. 2018, 241, 41–50. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Z.Y.; Zhao, X.F.; Yu, G.K.; Zhang, X.Y.; Liang, J.; Zhang, L.S. Inhibitory effects of 5-aminolevulinic acid on apple pollen germination and growth, and its thinning effects on flowering. J. Shanxi Agric. Univ. Nat. Sci. Ed. 2023, 43, 52–59. [Google Scholar] [CrossRef]
- Mishra, S.N.; Srivastava, H.S. Stimulation of nitrate reductase activity by delta aminolevulinic acid in excised maize leaves. Experientia 1983, 39, 1118–1120. [Google Scholar] [CrossRef]
- Feng, X.X.; An, Y.Y.; Gao, J.J.; Wang, L.J. Photosynthetic responses of canola to exogenous application or endogenous overproduction of 5-aminolevulinic acid (ALA) under various nitrogen levels. Plants 2020, 9, 1419. [Google Scholar] [CrossRef]
- Zheng, J.; An, Y.Y.; Wang, L.J. 24-Epibrassinolide enhances 5-ALA-induced anthocyanin and flavonol accumulation in calli of ‘Fuji’ apple flesh. PCTOC 2018, 134, 319–330. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhang, J.; Zhong, Y.; Zhang, L.; Yang, H.; Liu, L.; Zhou, J.; Iqbal, M.M.; Gan, X. Regulation of 5-Aminolevunilic Acid and Its Application in Agroforestry. Forests 2023, 14, 1857. https://doi.org/10.3390/f14091857
Wang L, Zhang J, Zhong Y, Zhang L, Yang H, Liu L, Zhou J, Iqbal MM, Gan X. Regulation of 5-Aminolevunilic Acid and Its Application in Agroforestry. Forests. 2023; 14(9):1857. https://doi.org/10.3390/f14091857
Chicago/Turabian StyleWang, Liangju, Jianting Zhang, Yan Zhong, Liuzi Zhang, Hao Yang, Longbo Liu, Jiayi Zhou, Malik Mohsin Iqbal, and Xing Gan. 2023. "Regulation of 5-Aminolevunilic Acid and Its Application in Agroforestry" Forests 14, no. 9: 1857. https://doi.org/10.3390/f14091857