Abstract
This study delves into the phytocenotic structure and biodiversity of forest ecosystems dominated by species of the family Betulaceae (Betula pendula, Betula pubescens, and Alnus glutinosa) in the Kostanay region of Kazakhstan. The research is a significant step toward understanding the influence of environmental factors, particularly hydrological regimes and soil conditions, on the formation and functioning of these forest communities. Field studies were conducted across multiple plots, where detailed geobotanical descriptions and analyses of species diversity were performed. The results underscore the significant species diversity, with 146 vascular plant species identified across various forest types, and are of great importance. Birch forests exhibit higher species diversity than alder forests, with the highest diversity observed in plots with favourable moisture conditions. The study concludes that the hydrological regime is critical in shaping these forest ecosystems’ phytocenotic structure and productivity. These conclusions underline the importance of biodiversity conservation and sustainable forest management in the region, making the implications of this research significant and far reaching.
1. Introduction
Our planet’s forest ecosystems are unique biological communities with diverse flora and fauna, and they play an essential role in maintaining the ecological balance. Forests, the most critical environmental framework of the biosphere, stabilise many natural processes, regulate the gas composition of the atmosphere, preserve the purity of water and the air basin, and protect the earth from water and wind erosion. They are solving the problem of global climate change, which is a necessary condition for the sustainable development of the world community [1,2,3,4,5].
In recent decades, the area of world forests has decreased by an average of 10–15 million hectares per year [6]. Half of the tropical and subtropical forests have disappeared, and the area of forests in Europe, America, and Asia has decreased by 2–3 times, accompanied by a sharp decrease in biodiversity. The quality of forests in some border countries has already been undermined [7,8,9,10]. Due to poor forest management, large-scale replacement of economically valuable species with low-value ones continues, and the system of state forest management has been destroyed.
Birch forests play a crucial role in maintaining the region’s ecological balance, with their functions differing between boreal and temperate zones. In boreal zones, they protect the land from erosion, regulate water regimes, and preserve biodiversity. In temperate zones, they act as primary forests in succession processes following disturbances, such as fires or logging, helping to restore ecosystems. These forests thrive in diverse environments, ranging from elevated areas with limited resources to low-lying wetlands, providing a variety of habitats that support vegetation development [11].
Birch forests are changing worldwide and are subject to anthropogenic threats—logging, fires, diseases, and the impact of climate change [12]. This allows for identifying vulnerable areas, developing a well-thought-out strategy for managing and protecting forest resources, and ensuring their sustainable development in the future. Birch forests are essential components of a careful attitude. With their help, it is possible not only to preserve a region’s biodiversity but also to ensure the sustainability of the ecosystem for many years to come. However, this requires the joint efforts of society, government, and environmental protection agencies.
The formation and composition of forest communities, the study of the structure of which allows us to identify the main types of forests, their distribution, and dominant plant species, are influenced by abiotic and biotic factors. Analysis of changes in the structure and composition of forest ecosystems over time allows us to assess their development dynamics, the influence of anthropogenic impacts, and natural processes in the states of forests [13].
Assessing the contribution of forest ecosystems to the conservation of biodiversity, maintaining the balance of water and soil resources, and their impact on climate processes is an essential aspect of studying the phytocenotic structure of forests. Identifying threats to forest ecosystems and developing effective measures for their protection and sustainable management also play crucial roles in preserving forests’ value and biological diversity [14,15].
In this paper, we review essential aspects of the birch forest community of the Kostanay region and their importance to the environment. Birch forests of the Kostanay region (Kazakhstan) correspond to the level of species diversity of both woody and non-woody plants. Silver birch (Betula pendula) and downy birch (Betula pubescens) often dominate the tree community, while a variety of grasses, shrubs, and mosses thrive in the understory [16].
The main goal of this study is to study the key features of the phytocenotic structure of birch and alder forest ecosystems, as well as to analyse the influence of environmental factors on their formation and function at the present stage, as well as to study the biological diversity of neighbouring species. Studying these aspects will allow a deeper understanding of the features and significance of forest ecosystems in this region in the context of biodiversity conservation and sustainable development of natural resources of the forests of the Kostanay region of northern Kazakhstan.
2. Materials and Methods
2.1. Study Area
According to the Committee on Ecology, Geology and State of Resources in Kazakhstan, as of 1 January 2022, forests occupy 5% of the country’s territory and cover an area of 13.6 million hectares [17]. Forest ecosystems are the object of special attention and study in the Kostanay region, located in the centre of Northern Kazakhstan. The features of this region’s phytocenotic structure of forest ecosystems are determined by its unique natural conditions, including climatic features, geological structure, relief, soil composition, and hydrological regime. The impact of these factors forms the diversity of forest communities and their composition and structure, making the region a fascinating area for biodiversity conservation and sustainable development [18].
We studied birch and alder (relic) forests of natural origin, classified as small-leaved forests by their specificity. To determine the natural regeneration of the studied forests, 20 survey plots were laid under the forest canopy. Then, on the main test plot, plots of 4 square meters were laid to determine the amount of natural seeding and undergrowth in stands untouched by logging, with selective logging, and with clear logging. Selecting the sample size and allocating sites for the birch forest study in the Kostanay region are vital steps to ensure that the study is representative. We determined the sample size based on the soil, climate, and anthropogenic activity. The sites considered differences in birch forest type depending on proximity to water bodies and soil type. Anthropogenic load was considered in protected and unprotected areas, which allowed us to assess the impact of human activity on birch forests. Covering various conditions provides more accurate recommendations for birch forest resource management. The study areas are shown in Figure 1.
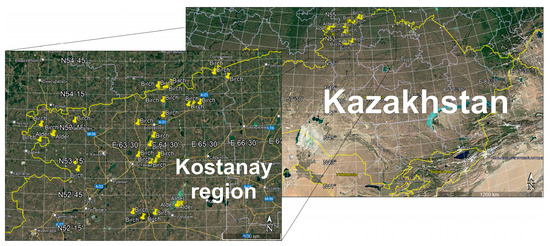
Figure 1.
Study areas of birch and alder forests in the Kostanay region.
The outstanding scientist P. G. Pugachev [19] contributed significantly to the study and protection of forests in Kazakhstan, including the Kostanay region, from 1932 to 2012. His work covers a wide range of aspects of forestry, from floristic studies, the ecological state of forests, forest management, and forest protection measures to sustainable forest management issues. His recommendations for improving forest resource management and environmental protection are still being implemented. For a complete understanding of the phytocenotic structure of forest ecosystems in the Kostanay region, it is necessary to consider the species diversity of vegetation. Studying the diversity of the flora of forest ecosystems allows us to determine this region’s main plant species characteristics and assess their contribution to the management of biological diversity [20]. In general, the study of the phytocenotic structure and biological diversity of associated species of forest ecosystems in the Kostanay region is essential for developing strategies for the sustainable use and protection of forest resources in this region, as well as for understanding their role in maintaining ecological balance at local and global levels [21].
Forest ecosystems of the Kostanay region are characterised by various phytocenoses—complexes of plant communities—each of which has its own type and species composition. For example, typical pine forests in this area can have different age classes and include various types of herbs, shrubs, and vines. Birch forests are considered as more diverse communities—Alder—relic communities. Despite the natural diversity, forest ecosystems of the Kostanay region are developing anthropogenic sunlight. Illegal logging, deforestation for agricultural agriculture and industrial development, and environmental pollution, necessitate protection from the harmful effects of air pollution in forest ecosystems. Changes in forest management and natural disasters, such as fires and droughts, also put pressure on ecosystems [22].
Forests in the Kostanay region do not represent the dominant type of vegetation. According to the forest management data of 2022, the forest fund in this territory is 447.1 thousand hectares, the area covered by forest is 215.2 thousand hectares, the formation of birch forests accounts for 103.9 thousand hectares, aspen—21.1 thousand hectares, pine—74.1 thousand hectares, willow forests and shrubs make up 8.4 thousand hectares, and only about 200 hectares are occupied by alder forests [23].
A characteristic feature of the forests of the Kostanay region is their heterogeneity in the composition of tree species. This is due to a complex of ecological and geographical factors determined by the border position of the territory with the forest-steppe subzone of the West Siberian Lowland in the northeast, the steppe of the Ubagan-Ishim interfluvial territory in the east, the low-mountain province of the Trans-Urals steppe in the west, and the northern border of the semi-desert in the south of Turgay [24,25].
Birch forests are among the most characteristic and widespread in the Kostanay region. The family Betulaceae, or Birch, includes about 120 species of trees and shrubs distributed mainly in the temperate climate of the Northern Hemisphere. The most famous representatives of these forests include birches (Betula), alders (Alnus), hornbeams (Carpinus), hazels (Corylus), and others. Betulaceae are found in many habitats, from low swamps to mountain slopes. In Russia and Kazakhstan, these plants are often found in forest and forest-steppe zones, forming mixed and pure stands. Representatives of Betulaceae play a significant role in ecosystems, participating in forest ecosystems and preserving the soil. They are often pioneer counters, populating disturbed areas and contributing to the restoration of the ecosystem. Birches, for example, are often the first to colonise clearings and burnt areas due to their ability to increase and resilience. Their uniqueness, richness of biodiversity, and compliance with the ecological balance make them the object of special attention for scientific research and conservationists [26].
The Kostanay region has specific ecological features that influence the selection of sites for birch research. First, this moderately sharp continental climate with cold winters and warm summers creates biotopes for Betula pubescens and Betula pendula. Soil features and their diversity (chernozem, sandy loam, peat, and solonetzic soil) affect the distribution and health of birches. The presence of rivers and lakes, as well as the relief, including hills and plains, create different microclimates and conditions conducive to the growth of birches. Anthropogenic factors, such as agriculture and logging, contribute to the change in natural ecosystems and the deterioration of the living conditions of birch forests. The sites for the study of the birch family were selected in such a way that they represent the entire spectrum of these ecological features. For example, selecting areas near water bodies, on different types of soil, and with varying landscape characteristics will allow us to draw conclusions about the influence of these conditions on the diversity and condition of birch trees in the region as a whole and develop recommendations for their conservation and restoration.
2.2. Floristic and Ecological Methods of Analysis
The main key areas were distributed in the Auliekol, Kostanay, Altynsarin, Karabalyk, Uzunkol, and Mendykarin districts of the Kostanay region. The flora analysis material is the collection of herbarium materials stored in the Herbarium of Kostanay Regional University, named after A. Baitursynov. The work process also studied literary sources and studies of the area. Studies of the main birch and alder communities in the field adhere to the generally accepted methodologies of V.N. Sukachev, V.S. Zon, and G.P. Motovilov [27] and V.N. Sukachev and N.V. Dylisa [28]. Geobotany was assessed using forest stand taxation methods [29]. When characterising the soil conditions of the study areas, generally accepted soil taxation and classification methods were used based on these works. Additionally, the recommendations of the “Diagnostic Horizons of Soils in the World” were used to classify and describe soils, considering the characteristics of the soil cover of the territory of Kazakhstan [30].
Geobotanical descriptions were compiled using a specially developed form, which noted the geographical location of the sample area, including the site’s location in the relief, environment, and microrelief. More than 180 geobotanical descriptions of phytocenoses were conducted. To compile the definition of the complex features, at least 3–6 sample areas of 0.25–0.5 hectares were laid out. Taxation of the ancient standard for forest elements and canopies, the timber stock, corresponded to the tables of standard density and stocks of regular stands, the bonnet was determined visually, and the composition of the forest stand was determined by the stem species in the sample area. The species composition, height, and general conditions were considered when describing the shrub layer. The species were verified by the evidence of POWO [31]. The types of composition, relief, and cover (in %) were noted when describing plant communities. The selected species’ height (in cm or meters), size, and vegetation type were distinguished. The kinds of cultural use were identified—“Flora of Kazakhstan” [32], “Flora of the USSR” [33], “Flora of Siberia” [34], and “Identifier of Plants of Central Asia” [35]. The number of species was considered using the Braun–Blanquet cover-abundance scale [36]. During field research, they moved objects in various ecosystems of the Kostanay region. Samples were carefully documented with an indication of their location and conditions. Based on the identification of species, taxonomic categories were determined. The status of species in nine-fold rarity and inclusion in the Red Book were determined. This allowed us to assess the degree of threat to individual species and their status in the ecosystem. The phytocenotic composition of the forest ecosystems was studied, allowing us to determine the species’ ratio, diversity, and habitats. This is important for understanding the sustainability of the ecosystem and its ability to recover. All data were recorded and processed for further use in scientific research and developing measures to restore forest cover and preserve plant diversity.
2.3. Methods for Analysing Biological Diversity
The leading indices for assessing biological diversity were used as the method for studying biological diversity. The data included various biodiversity metrics for the birch and alder sites. Here are the key metrics and their values for each site: Taxa S: Number of taxa (species) in the site. Individuals: Total number of individuals. Dominance D: Dominance index. Simpson 1-D: Simpson index. Shannon H: Shannon index. Evenness eH/S: Evenness index. Brillouin: Brillouin index. Menhinick: Menhinick index. Margalef: Margalef index. Equitability J: Equivalence index. Fisher alpha: Fisher’s alpha. Berger–Parker: Berger–Parker index. Chao-1: Estimation of species richness by the Chao1 index.
Table 1 analyses the Braun–Blanquet and the Drude scales as a systematic way to assess the frequency of occurrence and projective cover of plant species in an ecosystem. The frequency of occurrence indicates how often a particular species can be found in the study area. A high frequency of occurrence indicates a species that has adapted to a given ecosystem environment and occupies a large area.

Table 1.
Braun–Blanquet and Drude scales.
Coverage reflects the degree of dominance of a species in a plant community. A species with high coverage has a significant impact on the structure and functions of an ecosystem. The Braun–Blanquet and Drude scales provide additional classification systems for assessing plant communities. These scales help scientists and ecologists to compare data between different studies and regions by providing a standardised description of plant communities. Using the scales in a study helps to identify species that are key to maintaining ecosystem structure and rare and threatened species that require conservation and protection.
3. Results
3.1. Phytocenotic Analysis
According to the features of the phytocenotic structure of forest stands in the studied region, forest formations are distinguished and represented by deciduous species, which include birch and alder forests. Birch forests are the most widespread in the studied area, which, by their specificity, belong to small-leaved forests of the Siberian type. Indigenous communities of birch forests are confined mainly to depressions of lake basins and to groves of deep depressions with moist soils. Betula pendula dominates in birch forests of dry and wet habitats of the studied phytocenoses; Betula pubescens and Populus tremula often act as codominants. Alder forests in the region occupy small areas. The main areas grow near Lake Kushmurun, at the exit of springs near the village of Chilik-Kuduk and north of the town of Borovskoye in the Mendykarinsky district, and along the bank of the Toguzak River in the vicinity of the rural districts Mikhailovka and Nadezhdinka in the Karabalyk district. Below is a description of the studied plant communities of forest ecosystems of the family Betulaceae of the Kostanay region in the studied areas:
Plot 1 (birch forest): The plot is located in the Mendykarinsky district’s forest-steppe zone in the villages of Teniz, Uzynagash, and Lake Borovskoye; in the Uzynkolsky district, in the vicinity of the towns of Ershovka, Otynagash, and Uzynkol; and also in the Karabalyksky district, in the area of the villages of Prirechnoye, Mikhailovka, and Boskol. The soils in the depressions are moist and humus. In the territory of the Borovskoye forest, there is sod-pine sandy soil and medium humus. Birch grows in the steppe plain, forming birch groves in saucer-shaped hollows and hilly hills. The horsetail-forb birch forest community has been studied and described in this plot—community of horsetail-forb birch forest (Betula pendula—Equisetum sylvaticum—herbosum). The tree layer is formed by Betula pendula, sometimes with an admixture of B. pubescens 8 B.p. 1 B.p. Birch trees are of average height—9 m, diameter—17 cm, age—30–40 years, Bonitet II, and crown density is 0.7. Wood stock per 1 ha is 140 m3. Birch regeneration is weak. The species composition of this community is represented by 51 species from 44 genera and 27 families. The undergrowth contains Cerasus fruticosa, Rosa canina, R. acicularis, Frangula alnus, Ribes nigrum, Salix rosmarinifolia, S. caprea, and S. sinerea. Equisetum sylvaticum dominates in the grass layer; Equisetum hyemale and E. fluviatile act as codominants; and Glycyrrhiza uralensis, Heracleum sibiricum, Fragaria viridis, Veronica spuria, Viola epipsila, Rumex acetosella, Scutellaria galericulata, Carex elongata, C. riparia, Urtica dioica, Galium palustris, Geum rivale, Scirpus sylvaticus, Filipendula ulmaria, Epilobium hirsutum, and Artemisia vulgaris are often found. Rubus saxatilis, Solanum dulcamara, and others are found in saucer-shaped depressions of birch groves. The average height of the grass stand is 65 cm, and the total projective cover is 60%–70%.
Plot 2 (birch forest): The plot is in the Uzunkolsky district near the villages of Presnogorkovka, Troebratsky, and Rassolnoye Lake; the Mendykarinsky district near Kamensk-Uralsk village; and the Kamennoye Lake tract—Kostanay district. Soils are soddy-sandy and humus. Birch grows on hilly elevations and the slopes of lakes and rivers. The community of marsh-fern birch forest was studied and described in this plot—marsh-fern birch forest community (Betula pubescens—Thelypteris palustris). The tree layer comprises Betula pubescens, B. pendula, and Pinus sylvestris. The composition of the tree stand is 6 B.p., 2 B.p., and 2 S.o. Crown density is 0.7–0.8. The average height of downy birch trees is 11 m, and the average diameter is 18 cm. The average age is 45 years. Bonitet II. The timber stock is 80–90 m3. Weak undergrowth of downy birch is noted. Regeneration is weak. The species composition is represented by 44 species, including 36 genera and 19 families. The undergrowth is poorly developed. The main species are Cerasus fruticosa, Frangula alnus, Rosa canina, Crataegus sanguinea, C. altaica, Salix caprea, S. pentandra, S. sinerea, etc. The herbaceous layer is dense, dominated by Thelypteris palustris; codominants are Matteuccia strutiopteris, Heracleum sibiricum, and Caltha palustris; there are Athyrium filix-femina, Stachys palustris, Lathyrus palustris, Equisetum pratense, E. fluvitile, Solanum dulcamara, Veronica officinalis, Oxalis acetosella, Chelidoniun majus, Angelica sylvestris, Ranunculus repens, Adenophora lilifolia, and Campanula glomerata; and on the edge of birch groves grow Fragaria viridis, Linaria vulgaris, Viola mirabilis, Geum rivale, etc. Of the herbaceous vines, Humulus lupulus, grows abundantly. The average height of the grass stand is 60–80 cm, and the total projective cover is 75%–80%.
Plot 3 (birch forest): The plot is located in the forest-steppe plain zone of the Auliekolsky district in the vicinity of the villages of Lesnoye, Sosnovka, and Kazanbasy. The Kustanai district is in the towns of Vladimirovka and Stepnoye, and the Altynsarinsky district is in the area of the cities of Shcherbakovo, Satai, and Basbek. The soils are chernozem and chestnut-humus in places. Birch grows in continuous strips and groves with Scots pine and aspen. In this plot, the rosehip-drupaceous birch forest community was studied and described—rosehip-drupaceous birch forest community (Betula pendula—Rosa acicularis—Rubus saxatilis). The stand consists of Betula pendula with an admixture of Pinus sylvestris and rarely Populus tremula 6 B.p., Bonitet II. Crown density is 0.6. Trunk height is 12–14 (18) m, average diameter is 20 (30) cm, age is 40–50 (70) years. Wood stock is 120–130 m3. The species composition is represented by 43 species, including 37 genera and 17 families. Rosa acicularis dominates in the undergrowth; codominants are Rosa canina, Cerasus fruticosa, Padus avium, Crataegus sanguinea, Salix sinerae, S. Caprea, Frangula alnus, and others. Rubus saxatilis dominates in the herbaceous layer. Codominants are Urtica dioica, Glycyrrhiza uralensis, Solanum dulcamara, Filipendula ulmaria, Ranunculus repens, Lycopus europacus, etc. Rare, relic, and boreal species are found: Heracleum sibiricum, Equisetum hyemale, E. palustris, Gymnocarpium dryopteris, etc. The average height of the grass stand is 80 cm, and the total projective cover is 80%–90%.
Plot 4 (birch forest): The plot is located in the forest-steppe zone of the Karabalyk district in the vicinity of the villages of Boskol, Yesenkol, and Slavyanka, and the Mendykarinsky district in the area of the towns of Tenizovskoye, Tulengut, and Baykozha. Birches grow on hilly elevations and, in some places, are confined to saucer-shaped depressions. Chernozem soils. They form communities of cherry-forb birch forests and stone birch forests—a community of cherry-forb birch forests (Betula pendula—Cerasus fruticosa—herbs). The tree layer consists of Betula pendula with an admixture of Populus tremula. The forest stand is 8 B.p. 2 Os., Bonitet III. The average height is 15 m, the trunk diameter is 22 cm. The crown density is 0.8. Age is 45–50 years. The undergrowth of birch and aspen is average. The timber stock per 1 ha is 280–312 m3. The species composition is represented by 48 species, including 43 genera and 23 families. The undergrowth is dominated by Cerasus fruticosa, the average height of which is 1.5–2 m; also growing are Ribes nigrum, Rosa cinamomea, Frangula alnus, Salix caprea, S. pentandra, and others. In the herbaceous layer, the characteristic species are Glycyrrhiza uralensis, Chelidonium majus, Solanum dulcomara, Stachys palustris, Lythrum virgatum, Phragmites communis, Equisetum palustre, Orchis maculate, Solanum dulcomarum, Geranium sylvaticum, Myosotis palustris, Plantago major, and others. Shrubs and trees are entwined with liana—Humulus lupulus. The average height of the grass stand is 80–90 cm, and the total projective cover is 75%—birch-stone community (Betula pendula—Rubus saxatilis). The tree layer consists of Betula pendula with an admixture of Populus tremula and Crataegus sanguine. The forest stand is 7 B.p. 2 Os. 1 B.k., Bonitet II. The average height is 12 m, the trunk diameter is 19 cm. The crown density is 0.7. Age is 40–45 years. The undergrowth of birch and aspen is weak. The timber stock per 1 ha is 100–120 m3. The species composition is represented by 48 species, including 41 genera and 03 families. The dominant species in the undergrowth is Crataegus sanguine, codominants are C. altaica, Lonicera tatarica, and Cerasus fruticosa, and the characteristic species of this community are also Ribes nigrum, R. idaeus, Rosa cinamomea, R. canina, Frangula alnus, Salix caprea, S. pentandra, and S. cinerea. The dominant species in the grass layer is Rubus saxatilis, the codominant is Fragaria viridis, and common species are Glycyrrhiza uralensis, Chelidonium majus, Solanum dulcomara, Stachys palustris, Lythrum virgatum, Phragmites communis, Equisetum palustre, Orchis maculate, Solanum dulcomarum, Geranium sylvaticum, Myosotis palustris, Plantago major, etc. The average height of the grass stand is 80–85 cm, and the total projective cover is 80%.
Plot 5 (birch forest): The plot is located on the territory of the Mendykarinsky and Uzunkolkolsky districts in the floodplain of the Tobol River near the village of Uzynagash and the Toguzak River near the towns of Nadezhdinka and Verenka in the Karabalyk district. The soils are chestnut, with additional moisture, in places on the hills, they are stony-loamy. The communities of hawthorn-cereal birch forest and meadowsweet birch forest were studied and described—community hawthorn-cereal birch forest (Betula pendula—Crataegus sanguinea). The tree layer is formed by Betula pendula and Crataegus sanguine. Its composition is 7 B.p. 3 B.cr., Bonitet III. The average height of silver birch trees is 11 m, with an average trunk diameter of 19 cm. Age is 35–40 (50). The crown density is 0.6. The undergrowth of Betula pendula is average. The species composition is represented by 46 species, including 40 genera and 21 families. Crataegus sanguinea dominates in the undergrowth; codominants are Crataegus altaica and Cerasus fruicosa (in places, they form dense thickets); Rosa acicularis, R. canina, Lonicera tatarica, Salix cinerea, Frangula alnus, Rhamnus cathartica, and others also grow in patches. The grass stand is well developed. The dominant species in this layer are Poa nemoralis and Agropyron pectinatum, with Bromopsis inermis and Brachypodium pinnatum acting as codominants, and Stipa pinata is rare. Common species in the community include Urtica dioica, Veronica spuria, Trifolium repens, Sanguisorba officinalis, Ranunculus acris, Melandrium album, Geranium sylvaticum, Calamagrostis epigeios, Barbarea vulgaris, Glycyrrhiza uralensis, Lysimachia vulgaris, Galium palustris, Artemisia vulgaris, Myosotis palustris, etc. The community is mainly composed of species that are characterised by high constancy. The average height of the herbaceous layer is 80–90 cm; the total projective cover is 70%—birch-meadowsweet community (Betula pendula—Filipendula ulmaria). Betula pendula forms the tree layer with an admixture of B. pubescens. The composition is 7 B.p. 3 B.p., Bonitet II, the average height of trees is 10 (16) m, and the average diameter is 18 cm. Crown density is 0.7–0.8. Wood stock per 1 ha is 160 m3. Regeneration of birch is average. The species composition is represented by 42 species, including 30 genera and 18 families. The undergrowth is represented by the following species: Frangula alnus, Grataegus altaica, Rosa acicularis, Cerasus fruicosa, Salix caprea, S. pentandra, and S. cinerea. Filipendula ulmaria dominates in the grass cover. The following species are widespread: Urtica dioica and Stellaria holostea. The following are characterised by noticeable constancy: Lysimachia vulgaris, Geum rivale, Calamagrostis canescens, Ranunculus repens, Equisetum pratense, Scutellaria galericulata, Glycyrrhiza uralensis, Caltha palustris, Scirpus silvaticus, Carex riparia, etc. The climbing liana, Humulus lupulus, represents the non-tier vegetation. The average height of the grass stand is 80 cm, and the total projective cover is 70%–75%.
Plot 6 (birch forest): The plot is located in the forest-steppe zone of the Auliekolsky district, in the vicinity of the village of Lesnoye, on the territory of the natural monument “Aspen-birch groves with ordinary potash.” Chestnut soils. In this plot, research on the creation of a viburnum-mixed herb birch forest community was conducted, where rare and relic plant species are present. Community of viburnum-mixed herb birch forest (Betula pendula—Viburnum opulus—herbosum). Betula pendula forms the tree layer with an admixture of Populus tremula. Its composition is 7 B.p. 3 Os., Bonitet II. The average birch height is 10 m, aspen—5 m, and the average trunk diameter is 18 cm. The crown density is 0.6. Timber stock per 1 ha is 110 m3. Birch regeneration is weak. The species composition is represented by 51 species, including 46 genera and 22 families. The undergrowth is dominated by Viburnum opulus, the codominant is Cerasus fruicosa, and there are also thickets of Rosa accularis, R. canina, Frangula alnus, Rhamnus cathartica, Salix caprea, etc. The herbaceous layer is formed by Polygonatum odoratum, Thelypteris palustris, Caltha palustris, Primula longiscapa, Equisetum hyemale, Thalictrum minus, Platanthera bifolia, Pyrola rotundifolia, Pyrola chromantha, Spiranthes sinensis, Glycyrriza uralensis, Comarum palustre, Urtica dioica, Filipendula ulmaria, Galium palustre, Stellaria holostea, Glycyrriza uralensis, Phlomis tuberosa, Sanguisorba officinalis, Inula helenium, Geranium pratense, Veronica longifolia, Hieracium umbellatum, Geranium pratense, Lysimachia vulgaris, and others. The average height of the grass stand is 70 cm. The total projective cover is 70%.
Thus, the birch forests of the Kostanay region are unique ecosystems that play a significant role in preserving biological diversity. For their preservation and restoration, it is necessary to conduct a systematic annual triptych.
Plot 7 (alder forest): The plot is located on dangerous sites in the village of Klyuchevoe, Karasu district, and on the gentle slopes of the northwestern orientation of conservative ravines near Lake Kushmurun. Chestnut, peat-bog soils. Soil moisture due to springs. Black alder is produced partially on the ravine’s slope and populations in its lower part, where a community of marsh-fern alder and marsh-horsetail alder is formed. Community of marsh-fern alder (Alnus glutinosa—Thelypteris palustris). The tree layer is formed by Alnus glutinosa with an admixture of Betula rubescens—9 Ol. 1 B.p. Alder trees now have an average height of 18 m, diameter—20 cm, age of 45–50 years: Bonitet II–III, crown density of 0.8. Timber stock per 1 ha is 200 m3. Regeneration is by coppice. The species composition is represented by 39 species, including 31 genera and 19 families. The undergrowth is formed by Padus avium, Frangula alnus, Rosa canina, Crataegus sanguinea, and Ribes nigrum; willows are often found: Salix caprea, S. Pentandra, and S. sinerea. The grass layer is dominated by Thelypteris palustris, with Scutellaria galericulata, Carex elongata, C. riparia, Urtica dioica, Galium palustris, Geum competitore, Equisetum sylvaticum, E. hyemale, Driopteris filix-mas, Scirpus sylvaticus, Filipendula ulmaria, Phragmites australis, Epilobium hirsutum, Sium latifolium, and Rorippa sylvestris acting as codominants. Rubus saxatilis, Solanum dulcamara, and others are found on the ravine upland. The average height of the grass stand is 85 to 100 cm, and the total projective cover is 80%—community of marsh-horsetail alder (Alnus glutinosa—Equisetum sylvaticum). Alnus glutinosa forms the tree layer with an admixture of Betula pendula. The composition of the stand is 8 Ol. 2 B.p. The crown density is 0.8. The average height of alder trees today is 15 m, and the average diameter is 20 cm. Age is 45–50 years. Bonitet II. Medium-density alder undergrowth (shrub) is noted. Regeneration is average. The species composition is represented by 40 species, including 35 genera and 17 families. The shrub layer is formed by Cerasus fruticosa, Rosa accularis, Ribes nigrum, Salix rosmarinifolia, and Viburnum opulus. The herbage is dominated by Equisetum sylvaticum and Thelypteris palustris, codominants are Calamagrostis canescens, Heracleum sibiricum, Hypericum perforatum, Stachys palustris, Equisetum pratense, E. fluvitile, Veronica spuria, Oxalis acetosella, Angelica sylvestris, Ranunculus repens, Rubus saxatilis, Adenophora lilifolia, Artemisia vulgaris, Campanula glomerata, Fragaria viridis, Linaria vulgaris, Viola mirabilis, Geum competitore, and others. The average height of the herbage layer is 70 cm, and the total projective cover is 70%–75%.
Plot 8 (alder forest): located at the bottom of a deep ravine southeast of Lake Kushmurun. The soils are moist and humus-gley. Black alder trees grow on well-drained slopes and at the bottom of the ravine. Alders form communities of nettle and ostrich alder groves—a community of nettle alder groves (Alnus glutinosa—Urtica dioica). The tree layer is formed by Alnus glutinosa with an admixture of Betula pendula and Populus tremula 7 Ol. 2 B.p. 1 Os., Bonitet II. Crown density is 0.7. Trunk height is 18–20 m, average diameter is 25–30 (50) cm, age is 80–90 (100) years. Regeneration is satisfactory. Timber stock is 300–310 m3. The species composition is represented by 45 species, including 41 genera and 20 families. The undergrowth is represented by Cerasus fruticosa, Ribes nigrum, Rubus idaeus, Rosa acicularis, Cotoneaster melanocarpus, Crataegus sanguinea, Salix caprea, Frangula alnus, Viburnum opulus, etc. Urtica dioica dominates in the herbaceous layer, Gymnocarpium dryopteris is a codominant, and characteristic species of the community are Solanum nigrum, Filipendula ulmaria, Ranunculus polejnchemus, Solanum dulcomara, Lycopus europacus, etc. Boreal species are noted in this community: Angelica sylvestris, Heracleum sibiricum, Equisetum hyemale, and E. palustris. The average height of the herbaceous layer is 70–85 cm, and the total projective cover is 60%–70%—a community of black alder (Alnus glutinosa—Matteuccio struthiopteris). The tree layer is formed by Alnus glutinosa, with an admixture of Betula pubescens. The forest stand is 7 Ol. 3 B.p., Bonitet III. The height of black alder trees is, on average, 17 m, with a trunk diameter of 23 cm. Crown density is 0.9. Age is 60–65 years. Regeneration is coppice. Wood stock per 1 ha is 300–320 m3. The species composition is represented by 47 species, including 40 genera and 23 families. The undergrowth is represented by the following species: Viburnum opulus, Rosa cinamomea, Crataegus altaica, Ribes nigrum, Frangula alnus, Salix caprea, and S. cinerea. The dominant species in the herbaceous layer is Matteuccio struthiopteris, and the codominant species is Thelypteris palustris. The following species are found in this community: Dryopteris cristata, Chelidonium majus, Solanum nigrum, Stachys palustris, Lythrum virgatum, Phragmites australis, Equisetum palustre, Orchis maculate, Geranium palustris, Myosotis palustris, Veronica longifolia, Plantago major, etc. The woody and shrubby vegetation is twined by Humulus lupulus. The average height of the herbaceous layer reaches 100 cm, and the total projective cover is 70%–75%.
Plot 9 (alder forest): The plot is 5 hectares in the southeastern part of Lake Kushmurun on the slope of deep ravines, where springs (aquifers) break through. Peat-bog and gley soils. Abundant moisture, flowing. The black alder grove has been studied and described as a black alder grove community (Alnus glutinosa—Athyrium filix-femina). The tree layer is formed by Alnus glutinosa with an admixture of Populus tremula. Composition is 8 Ol. 2 Os. Bonitet II. The average height of black alder trees is 20 m, with an average trunk diameter of 24 cm. Age is 50–55. Crown density is 0.7. Middle undergrowth of Alnus glutinosa and weak Populus tremula are noted. The species composition is represented by 44 species, including 38 genera and 19 families. In the undergrowth, Ribes nigrum dominates, and Rosa acicularis acts as a codominant; characteristic species of the community are Rosa canina, Crataegus sanguinea, Frangula alnus, Rhamnus cathartica, Viburnum opulus, etc. In the herbaceous layer, which is well expressed, Athyrium filix-femina dominates; Filipendula ulmaria and Dryopteris filix-mas act as codominants; and Urtica dioica, Thelypteris palustris, Phragmitis communis, Lysimachia vulgaris, Carex elongata, Galium palustris, and Viola epipsila are often found. The community is formed mainly by moisture-loving species characterised by high constancy. The average height of the grass stand is 70–85 cm, and the projective cover is 60%–70%.
Plot 10 (alder forest): The plot is located near the village of Mikhailovka, along the bank of the Toguzak River and the terrace floodplain. The soils are marsh-meadow and loamy—flowing moisture. The community of the reed-meadow alder grove was studied and described—a community of reed-meadow alder grove (Alnus glutinosa—Scirpus sylvaticus—Filipendula ulmaria). The tree layer is represented by Alnus glutinosa, with an admixture of Betula pendula. Composition is 6 Ol. 4 B.p. The average height of Alnus glutinosa is 16 m, and the average diameter is 21 cm. Crown density is 0.5–0.6. Bonitet I–II. Regeneration of black alder is weak. Wood volume per 1 ha is 180–200 m3. The species composition of this community is represented by 49 species, including 43 genera and 21 families. The undergrowth is represented by Ribes nigrum, Viburnum opulus, Salix caprea, S. cinerea, Padus racemosa, and Rosa canina. The herbaceous layer is dominated by Scirpus sylvaticus and Filipendula ulmaria. The codominants are Lysimachia vulgaris and Athyrium filix-femina; and the characteristic species of the community are Urtica dioica, Galium palustre, Achillea millefolium, Solanum dulcamara, Fragaria viridis, Phragmites australis, Urtica dioica, Equisetum palustre, Geum urbanum, Carex riparia, and C. leporina. The herbaceous layer is quite dense and is represented by the off-layer vegetation, liana—Humulus lupulus. The average height of the grass stand is 100–110 cm, and the total projective cover is 70%–80%.
Characterising birch forest communities by forest type, it should be noted that communities differ in location and growing conditions. In populations of open elevated areas of the steppe zone, shorter trees dominate than in the lower reaches of depressions and basins and the vicinity of rivers. The average diameter is lower on more prosperous and humid soils, where more individuals grow. In each studied area, depending on the environmental conditions, such as the terrain and soil type, the height of the trees is different. A more detailed comparative analysis of all sites is presented in Table 2.
Table 2.
Comparative table of the floristic diversity of the studied plots.
Birch forest areas (1–6) have high species diversity, especially in the herb layer. The main dominant species include Equisetum sylvaticum, Glycyrrhiza uralensis, and Rubus saxatilis. Alder forest areas (7–10) are also characterised by significant species diversity, especially in the understory and herb layer. Dominant species include Thelypteris palustris, Scirpus sylvaticus, and Filipendula ulmaria. Birch forests are characterised by various dominant species depending on the habitat type: horsetail-forb, bog-fern, rosehip-drupe, etc. Alder forests include bog-fern, bog-horsetail, nettle, and ostrich communities, reflecting their preference for more humid conditions. The average height of the grass stand varies from 60 to 110 cm in different areas, indicating differences in microclimatic and soil conditions. The total projective cover also varies from 60% to 90%, indicating the density and structure of plant communities. Soil conditions and the presence of additional moisture play a vital role in the formation of plant communities. For example, soddy-sandy soils contribute to the development of bog-fern communities, while chestnut soils support various herbaceous communities. This comparative floristic analysis allows us to highlight the key features of different areas of birch and alder forests and their role in maintaining biological diversity in the Kostanay region.
3.2. Analysis of Biological Diversity
Biodiversity is one of the critical indicators of the ecological health of ecosystems. Assessment and analysis of biodiversity allow us to identify the ecosystem’s current state and predict possible future changes. This study examines the leading biodiversity indices for ten study sites divided into two categories: birch forest (sites 1–6) and olshan (sites 7–10). The indices used include the total number of taxa (Taxa S), the Dominance index (Dominance D), Simpson index (Simpson 1-D), Shannon index (Shannon H), Brillouin index, Menhinick index, Margalef index, Berger–Parker index, and Chao1 species richness assessment. These indices allow us to obtain a comprehensive picture of the distribution of species and their level of dominance in each site. Table 3 shows the central values.
Table 3.
Leading indicators of biological diversity indices for the studied areas.
The main observations can be presented as follows. Total taxa (Taxa S): the maximum number of taxa is Birch Forest 5 (121), the minimum number of taxa: Birch Forest 1 (28). Dominance index (Dominance D): the highest dominance value is Birch Forest plot 1 (0.03571), the lowest dominance value: Birch Forests 2 and 3 (0.009434). Simpson index (Simpson 1-D): maximum value is Birch Forest 5 (0.9917), minimum value is Alder Forest 9 (0.9821). Shannon index (H): maximum value is Birch Forest 5 (4.796), minimum value is Birch Forest 1 (3.332). Margalef index: maximum is Birch Forest 5 (25.02), minimum is Birch Forest 1 (8.103). Species richness assessment according to the Chao1 index: maximum is Birch Forest 5 (7381), minimum is Birch Forest 1 (406).
Based on the presented data, the following conclusions can be made: Birch forests have a significantly higher number of taxa and individuals than Alder forests. The highest abundance is observed in the Birch Forest 5 plot (121 taxa and 121 individuals), while the lowest is at Alder Forest 9 (56 taxa and 56 individuals). The Dominance index (Dominance D) is low in all plots, indicating the absence of solid dominance of any species. Birch Forest 5 has the lowest dominance (0.008264), indicating the highest level of evenness. The Simpson index (Simpson 1-D) is high in all plots, confirming the high biodiversity. The highest index is observed in Birch Forest 5 (0.9917). The Shannon index (Shannon H) is also high in all plots, especially in birch forests, with the highest value in Birch Forest 5 (4.796). The Brillouin index is high in all birch forests, especially in Birch Forest 5 (3.823), indicating high diversity and thus a rich and varied ecosystem. The Menhinick index shows significant diversity in all plots, especially in Birch Forest 5 (11.00). The Margalef index confirms the high level of species diversity in the birch forest sites, especially in the Birch Forest 5 site (25.02). The birch forest sites generally show higher biodiversity values than the alder forests, reflected in all indices and quantitative indicators. The most diverse and uniform site is Birch Forest 5. The alder forests have lower species diversity but still show significant biodiversity. The parameters of the number of taxa are presented as a graph in Figure 2.
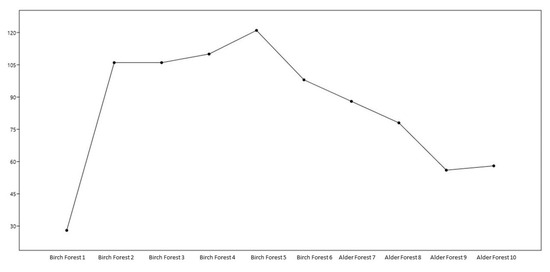
Figure 2.
Changes in the number of taxa in different areas.
The graph in Figure 2 shows the changes in the number of taxa in different plots: Birch plot 1: the lowest number of taxa (28). Birch plots 2, 3, and 4: the number of taxa increases and remains stable at 106–110. Birch plot 5: the maximum number of taxa (121). Birch plot 6: the number of taxa decreases to 98. Olsha plots: The number of taxa continues to decline from 88 to 56. The analysis shows significant differences in biodiversity between the plots. Birch plot 5 demonstrates the highest number of taxa and individuals and the high values of the Shannon and Margalef indices, indicating high biodiversity. These data can be helpful for further ecological research and the conservation of biodiversity (Table 4).
Table 4.
Assessment of species richness.
The analysis and visualisation of species richness estimates confirm that the observed number of species (196) is likely lower than the actual number of species present in the ecosystem. All estimation methods indicate higher numbers of species, with the Jackknife 2 method showing the highest potential species richness (238,278). The low standard deviations for the Chao 2 and Jackknife 1 methods confirm their reliability (Table 5).
Table 5.
Global beta diversities.
Whittaker’s Beta Diversity measures the number of species unique to each habitat, and its value is 1.3086, indicating moderate variation in species composition between habitats. Harrison’s Beta Diversity estimates the proportion of species that occur in more than one site, and its value is 0.1454, indicating low species overlap between habitats. Cody’s Beta Diversity measures the overall change in species counts between habitats, and its value is 253, indicating significant variations in species composition between habitats. Routledge’s Beta Diversity considers the total number of species and the unique species for each habitat, and its value is 0.25364, indicating moderate diversity between habitats. Wilson–Shmida Beta Diversity measures the number of species unique to each habitat compared to their distribution; its value is 2.98, indicating high beta diversity, meaning there is significant variation in species composition between habitats. Mourelle’s Beta Diversity measures the proportion of species unique to each habitat; its value is 0.33111, indicating moderate beta diversity. Harrison 2 Beta Diversity is a modified version of Harrison’s Beta Diversity. Its value is 0.068871, indicating low species overlap between habitats. Williams’ Beta Diversity measures the uniqueness of species for each habitat; its value is 0.38265, indicating moderate beta diversity. An analysis of global beta diversity metrics shows that the most significant species diversity between habitats is observed for the Wilson–Shmida metric (2.98), indicating substantial variation in species composition. The Cody (253) and Whittaker (1.3086) metrics also confirm considerable habitat diversity. The Harrison (0.1454) and Harrison 2 (0.068871) metrics show low species overlap, confirming the uniqueness of species in each habitat. Other metrics such as Routledge (0.25364), Mourelle (0.33111), and Williams (0.38265) indicate moderate beta diversity.
An analysis of global beta diversity metrics shows that the most significant species diversity between habitats is observed for the Wilson–Shmida metric (2.98), indicating substantial differences in species composition. The Cody (253) and Whittaker (1.3086) metrics also confirm significant habitat differences. The Harrison (0.1454) and Harrison 2 (0.068871) metrics show a low level of species overlap, confirming the uniqueness of species in each habitat. Other metrics such as Routledge (0.25364), Mourelle (0.33111), and Williams (0.38265) indicate moderate beta diversity.
Table 6 and Table 7 is a pairwise distance matrix showing the distances between plots labelled “Birch” and “Alder.” The values on the diagonal of the matrix are equal, which is logical since the distance from a plot to itself appears equal. Within-group distances, for example, between birch plots, are generally lower than between-group distances between birch and alder plots. This indicates that the birch and alder plots provide two cluster expansions with increasing distances within groups and rising between them. The smallest distance between birch plots is 0.14151, between plots “Birch 2” and “Birch 3,” while the smallest distance between alder plots is 0.3494, between plots “Alder 1” and “Alder 2.” The most incredible distances are observed between plots of different species, for example, between “Birch 1” and “Alder 2”—0.81132. This indicates a clear separation of birch and alder plots, probably due to differences in the ecological or average conditions that define these species.
Table 6.
Analysis of the pairwise distance matrix.
Table 7.
Number of species and standard error.
The increase in species richness is observed in the steady rise in species (Taxa) with increasing sample number. The maximum number of species is observed in the 10th sample (196 species). The standard error (Std. err) varies from 4.0966 to 4.9139. The highest error is observed in the 3rd sample (4.9139) and the lowest is observed in the 1st sample (4.0966). The analysis shows that the number of species steadily increases with increasing sample number. The standard error remains relatively low, indicating the stability of the data (Figure 3).
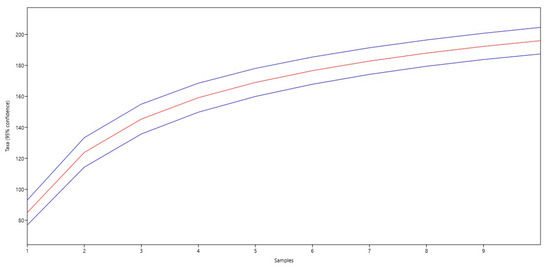
Figure 3.
Graph of the development of the number of species (the central red line shows the underlying trend, and the blue lines above and below indicate confidence intervals).
The graph shows a steady increase in species with an increasing sample number, indicating increasing biodiversity. The confidence intervals provide an idea of the possible variations in the number of species, which helps to better understand the data’s reliability. The data also represent the alpha diversity at different sites depending on the value of the alpha parameter (Figure 4).
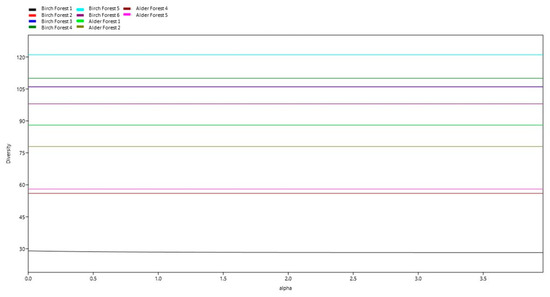
Figure 4.
Alpha diversity.
The graph displays the alpha diversity at different sites depending on the alpha parameter. The graph shows that the number of species at all sites, except Birch Site 1 remains constant with changing alpha. Diversity stability: at the sites Birch Site 2, Birch Site 3, Birch Site 4, Birch Site 5, Birch Site 6, Alder Site 1, Alder Site 2, Alder Site 3, and Alder Site 4, the number of species remains stable with changing alpha. Diversity change at Birch Site 1: at Birch Site 1, there is a slight decrease in the number of species with increasing alpha, which indicates a decline in species diversity with increasing alpha.
4. Discussion
The results of this study revealed significant diversity and specificity in the phytocenotic structure of forest communities dominated by Betula pendula, Betula pubescens, and Alnus glutinosa in Northern Turgai, which indicates the high ecological and biological significance of these ecosystems for the conservation of biodiversity under the conditions of the sharply continental climate of Kazakhstan. The study allows us to examine the factors influencing these communities’ formation and compare them with similar studies in other world regions [37].
One critical factor determining the phytocoenotic structure of the studied communities is the hydrological regime. Our data showed that the alder forests of Northern Turgai, formed in conditions of increased moisture, are characterised by high productivity (Bonitet I–II). By contrast, communities in less moist areas show lower productivity indicators. This fact is consistent with the findings of studies conducted in forest ecosystems of Northern Europe and Siberia, where a similar hydrological regime also significantly impacts the productivity and composition of forest stands [38].
Soil conditions also play an essential role in shaping the structure of forest communities. Our study found that chernozem and meadow-bog soils support the formation of diverse herbaceous layers dominated by species such as Urtica dioica, Filipendula ulmaria, Equisetum sylvaticum, and others. These species perform critical ecological functions, such as moisture retention, soil stabilisation, and participation in nutrient cycling, which is confirmed by similar research results in temperate and subarctic climates.
The floristic composition study showed the presence of 146 species of vascular plants in the grass layer of the studied communities, among which boreal relic species were identified. This biodiversity indicator is an essential indicator of environmental sustainability and indicates the significant biological diversity of the forest ecosystems of Northern Turgai. Similar data were obtained in studies of the forests of Finland and Norway, where boreal relics also play an essential role in maintaining the stability of ecosystems in a changing climate [39].
The role of Betula pendula and Alnus glutinosa as pioneer species capable of restoring ecosystems in disturbed areas should be noted separately. Their ability to vegetatively reproduce and form dense stands helps protect soils from erosion and quickly restore biological diversity. These aspects have also been mentioned in forest restoration work in Central and Eastern Europe [40].
Despite the standard features of other boreal forests, our data revealed several unique features inherent to the phytocenoses of Northern Turgay. For example, a high admixture of Betula pubescens in willow-forb alder forests (up to 40%) indicates specific environmental conditions characteristic of this region. These communities are formed in areas with stagnant water. They are distinguished by significant undergrowth and grass-layer development, which is not typical for similar communities in other boreal regions. These features are likely associated with the impact of the region’s unique microclimate and hydrological conditions, which require further study. The limitations of this study include its local nature and the time frame of data collection. To obtain a more complete picture of phytocenose dynamics, long-term observations covering different climatic conditions and seasons are needed. In the future, it is necessary to expand the geographical scope of studies and include other types of forest communities in the region in the analysis, as well as study the impact of climate change on forest dynamics and biodiversity.
Black alder forms pure stands and stands with an admixture of downy birch, silver birch, and aspen, and their role in different communities is not the same. The highest admixture of Betula pubescens is in willow-forb alder forests (up to 40%) and sedge forests (30%). In these communities, water stagnation is the highest. An admixture of silver birch (B. pendula) is typical for marsh-horsetail and lady fern alder forests, where it makes up (20%). Aspen (Populus tremula) is not a significant admixture in alder forests but only in ostrich and nettle forests (10%–20%). Willows are a common component of the undergrowth of all plant communities but are most typical for willow-forb and marsh-fern alder forests. The undergrowth of the studied communities is formed by the following species: Salix cinerea, Ribes nigrum, Spiraea crenata, Padus avium, Rosa acicularis, R. cinnamomea, R. canina, Cotoneaster melanocarpus, Frangula alnus, Rhamnus cathartica, and Lonicera tatarica. The richest undergrowth is that of the lady alder and nettle [21].
Grass cover is an essential indicator for identifying plant communities. Almost all plants are found in all studied phytocenoses. Still, for each of them, the most characteristic species achieve optimal distribution and abundance in a particular community, thus being a good indicator. Plants that are good indicators of each plant community include Matteuccia struthiopteris, Athyrium filix-femina, Thelypteris palustris, Urtica dioica, Filipendula ulmaria, Comarum palustre, Carex, Equisetum, etc. The indicator value of other species constantly found in alder forests is insignificant; despite their abundant distribution, they cannot form separate communities. The following species belong to this category: Caltha palustris, Lycopus europeaus, Solanum dulcamara, Galium palustre, Lysimachia vulgaris, Angelica sylvestris, Scutellaria galericulata, Stachys palustris, Glechoma hederacea, Ranunculus repens, and others, which are abundant in communities—sol., sp.
Black alder has seed and shoot origins in the studied phytocenoses. According to observations, seed renewal prevails in nettle-marsh-horsetail alder forests, and shoot renewal is typical for all phytocenoses. The limiting factor that characterises different productivity (bonitet) in alder forests is the hydrological regime of the soil. Black alder reaches the highest productivity in nettle and lady’s foot alder forests (Bonitet I); in meadowsweet, sedge, marsh-fern, and marsh-horsetail, the productivity of tree stands is Bonitet II. In willow-forb and ostrich phytocenoses, it is Bonitet III. Many grass cover plants have different degrees of distribution and development in various plant communities, which is a good indicator for them. Along with the forest stand quality, the composition of the tree layer, and the water regime indicators, these plants form phytocenoses [41].
The survival of Alnus glutinosa, together with other accompanying boreal species, in the conditions of the arid, sharply continental climate of the steppe part of Northern Turgai is facilitated by deep lowland ravines, with outlets to the surface of springs and river valleys. These habitats are refuges for preserving the relic species Alnus glutinosa with companion species. In characterising the plant communities of the alder forests of Northern Turgai, it was revealed that each has phytocenotic features [42]. Black alder forms pure stands and stands with an admixture of downy birch, silver birch, and aspen, and its role in different communities is not the same. The highest admixture of Betula pubescens is in willow-forb alder forests (up to 40%) and sedge (30%). These communities have the highest water stagnation. Admixture of B. pendula is typical for marsh-horsetail and yarrow alder forests, where it makes up (20%). Aspen (Populus tremula) is not a significant admixture in alder forests but only in ostrich and nettle (10%–20%). Willows are a common component of the undergrowth of all plant communities but are most typical for willow-forb and marsh-fern alder forests.
The undergrowth of the studied communities is formed by the following species: Salix cinerea, Ribes nigrum, Spiraea crenata, Padus avium, Rosa acicularis, R. cinnamomea, R. canina, Cotoneastr melanocarpus, Frangula alnus, Rhamnus cathatica, Lonicera tatrica, etc. The richest undergrowth is alder, scaly-leaved, and nettle. Grass cover is an essential indicator for identifying plant communities. Almost all plants are found in all the studied phytocenoses, but for each of them, the most characteristic species achieve optimal distribution and abundance in a particular community, thus being a good indicator. Plants that identify each plant community well include Matteuccia struthiopteris, Athyrium filix-femina, Thelypteris palustris, Urtica dioica, Filipendula ulmaria, Comarum palustre, Carex, Equusum, etc. [43].
The indicator value of other species constantly found in alder forests is insignificant; despite their abundant distribution, they cannot form separate communities. The following species belong to this category: Caltha palustris, Lycopus europeaus, Solanum dulcamara, Galium palustre, Lysimachia vulgaris, Angelica sylvestris, Scutellaria galericulata, Stachys palustris, Glechoma hede-racea, Ranunculus repens, and others, the abundance of which in communities is sol, sp.
Black alder has seed and shoot origins in the studied phytocenoses. According to observations, seed renewal prevails in nettle-marsh-horsetail alder forests, and shoot renewal is typical for all phytocenoses. The limiting factor that characterises different productivity (bonitet) in alder forests is the hydrological regime of the soil. Black alder achieves the highest productivity in nettle and lady’s foot alder forests (Bonitet I); in meadowsweet, sedge, marsh-fern, and marsh-horsetail, the productivity of tree stands is Bonitet II. In willow-forb and ostrich phytocenoses, it is Bonitet III.
Many grass cover plants have different degrees of distribution and development in various plant communities, which is a good indicator for them. Along with the bonitet of the forest stand, the composition of the tree layer, and the indicators of the water regime, these plants clearly distinguish phytocenoses. The diagnosis of the characteristics of each community we have identified includes an indication of its distribution, features of the habitat conditions, floristic composition, structure of vegetation layers, and taxation indicators of forest stands. Thus, the richness of the soil and favourable moisture regime contribute to the rapid development of the grass cover in the black alder forests of Northern Turgai. In the studied alder forest communities, 146 species of vascular plants were identified, among which boreal relic species were identified. More often, they occupy a subordinate position in the plant community of black alder forests, and Alnus glutinosa is dominant. The survival of Alnus glutinosa, together with other accompanying boreal species, in the conditions of the arid, sharply continental climate of the steppe part of Northern Turgai is facilitated by deep lowland ravines with outlets to the surface of springs and river valleys. These habitats are refuges for preserving relic species with their companion species.
Birch-alder-stone forest (Betuletum-Alnetum saxatilirubosum). It is found in small areas in the floodplain of the Toguzak River and the middle part of the slopes of the Duan Ravine in the vicinity of Lake Kushmurun. The soils under the forest are marsh-meadow, groundwater at a depth of 30 cm. The tree stand is formed of silver birch with an admixture of black alder and an insignificant participation of aspen. The composition of the tree stand is Bonitet II, the average height of black alder is 12 m, the maximum height is 18 m, the average diameter is 17 cm, the height of birch and aspen does not exceed 11 m, and the average diameter is 15 m. The crown density is 0.7. The wood stock per 1 ha is 128 m3. Regeneration of all species is satisfactory. The main components of the undergrowth are Rosa cinamomea, Ribes nigrum, Cerasus fruticosa, and Salix caprea. The grass cover is dense (projective cover 70%–80%). Rocky bramble dominates—a significant admixture in it is Fragaria viridis, Equisetum silvaticum, Galium boreali, Vicia sepium, Solidago virgaure, Poa pratense, Antennaria dioica, Gallatela punctata, and others. Moss-lichen cover is not noted, only in small areas. The extra-tier vegetation is represented by liana—Humulus lupulus.
The hydrological effect creates conditions for forming distinctive forest ecosystems depending on the area’s altitude. At higher elevations, the influence of the microclimate can be observed, which can affect the availability of water and, therefore, the structure of the forest. For example, the combination of good hydrology and soils rich in organic matter will promote a higher and healthier tree cover. It is essential to consider this interaction when developing strategies for protecting and restoring forest ecosystems such as alder forests, especially under changing climate conditions and the management of water consequences [44].
Climate change, especially in recent years, has significantly impacted the growth and development of birch forests in the Kostanay region. The average annual air temperature increases annually, which affects the phenology of birch, which can disrupt the harmony of its life cycle and that of associated organisms, and also leads to a decrease in the water content of rivers and lakes and, consequently, to a limitation of the hydrological regime, namely, its shallowing. An increase in the duration of hot periods leads to thermal stress and the instability of birches, which affects forest productivity. Due to succession, soil degradation occurs in some study areas. Inconsistent emissions lead to droughts that negatively affect birch growth, especially in depleted soils. Heavy rains cause soil erosion to reduce the birch’s root system and slow the spread of water and nutrients. Climate change leads to the poisoning or transfer of species from birch forests, contributing to pests and diseases, as well as danger to birches. Rapid changes make adapting difficult, and low genetic change affects the degree of adaptability, making template measures for forest restoration difficult. Changes in ecosystems lead to economic losses in forestry due to resource availability. Climate change poses significant ecosystem, environmental, and social challenges to birch forests in the Kostanay region [45,46].
Thus, in studying the phytocenoses of alder forests, it was noted that each habitat condition corresponds to a specific community of black alders. Each community differs from one another in structure, species, and ecological composition.
5. Conclusions
The results of this study are of great practical importance for developing strategies for sustainable management and protection of forests in Northern Kazakhstan. The features of the phytocenotic structure and biological diversity that we identified can be used in planning activities to conserve and restore forest ecosystems in a changing climate. An important step will be to include the data obtained in environmental monitoring programs and the development of recommendations for forest resource management. As a result of the research, we identified and described ten plant communities dominated by Betula pendula, B. rubescens, and Alnus glutinosa. Populus tremula participates in the stand along with birches. Rich soils and favourable moisture conditions contribute to the sound development of the grass cover in alder forests. In total, 146 species of vascular plants were noted in the grass layer of the studied communities. This study comprehensively assessed the forest ecosystems’ phytocenotic structure and biological diversity in Northern Turgai, particularly forest communities dominated by Betula pendula, Betula pubescens, and Alnus glutinosa. The floristic composition of the studied forest communities includes 146 species of vascular plants, among which boreal relic species were identified. The highest level of species diversity was observed in the birch forest of the fifth site, where 121 taxa were recorded. The minimum diversity was noted in the birch forest of the first site with 28 species. Forests with a predominance of birch are characterised by greater species diversity compared to alder forests. Dominance and evenness of species: the Dominance index (Dominance D) shows a low level of dominance of individual species, which indicates the uniformity of species distribution in communities. The lowest values of the Dominance index are noted in the birch forests of the second and third sites, which confirms the high uniformity of the species composition. The productivity of forest stands varies depending on moisture conditions and soil type. The most productive are nettle and fern alder forests, where the forest stand reaches Bonitet I. In birch forests in areas with drier conditions, productivity is lower, reflected in lower values of wood volume. The study showed that the hydrological regime plays a vital role in forming the phytocenotic structure of forest communities. Alder forests formed in increased moisture conditions are characterised by higher species diversity and productivity than forests in less moist areas. Biodiversity assessment: The highest biological diversity was recorded in the birch forests of the fifth site, which was confirmed by high values of the Shannon (4.796) and Margalef (25.02) indices. This indicates a significant diversity and uniformity of species distribution in this community. Biodiversity conservation and sustainable forest management: The study’s results highlight the need to conserve forest ecosystems of Northern Turgay to maintain ecological balance and biological diversity in the region. It is essential to develop strategies to protect forests from anthropogenic impacts and changing climate conditions to ensure their sustainable development in the future.
These results are consistent with similar studies of forest ecosystems worldwide, where hydrological regimes and soil conditions are crucial in shaping forest composition and biodiversity. For example, studies in boreal regions such as Northern Europe and Siberia also highlight the influence of moisture on species diversity and the resilience of pioneer species such as Betula pendula and Alnus glutinosa to ecosystem recovery after disturbances. Globally, forests with high moisture levels, as in our study, show higher species diversity, especially in sites with favourable moisture conditions, where the highest species diversity was recorded. In addition, identifying boreal relic species that contribute to ecological stability mirrors the results of studies in Scandinavian forests, where relic species help to maintain biodiversity in the face of climate change. Research from different regions of the world shows that the composition of the understory and herbage cover, as seen in birch forests, significantly affects the overall resilience of ecosystems, confirmed when comparing different sites and conditions.
Thus, the study confirmed the high ecological and biological significance of the forest ecosystems of Northern Turgay, which requires continued scientific research and the development of measures for their protection and sustainable use. In general, the results of this study highlight the importance of conservation and sustainable management of forest resources of Northern Turgay to maintain ecological balance and preserve biodiversity in this region.
Based on the data obtained, forestry enterprises in the study area can monitor the condition of birch forests, including assessing their biodiversity and resistance to climate stress, and plan conservation measures for their conservation and restoration. Creating multi-species forest plantations will increase the ecosystem’s resilience to climate change. Estimation of permissible volumes of timber harvesting, considering expected climate change, will prevent overexploitation of forests. Methods can be implemented that will improve soil structure and water retention capacity for birch’s total growth, such as forecasting and risk management. Creating crisis management plans will minimise damage from pests or tree diseases. Legal acts can be adopted to protect and preserve forest ecosystems while considering climate change. An integrated approach to managing birch forests in the context of climate change can contribute to their sustainability and development.
Author Contributions
Conceptualization, S.M.; methodology, S.K.; software, A.O.; validation, A.K.; formal analysis, D.D.; investigation, G.A. All authors have read and agreed to the published version of the manuscript.
Funding
The research was carried out within the framework of the target funding program of Ministry of Science and Higher Education BR18574125 “Study of the current state of species diversity of vascular plants of Kazakhstan using modern methods of botany, molecular genetics and bioinformatics” (2023–2024).
Data Availability Statement
Dataset available on request from the authors. The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zamolodchikov, D.G.; Korovin, G.N.; Utkin, A.I.; Chestnykh, O.V.; Songen, B. Carbon in Forest and Agricultural Lands of Russia; KMK Publication: Moscow, Russia, 2005; 200p. [Google Scholar]
- Pobedinskiy, A.V. Water and Soil Protection Role of Forests; VNIILM Publication: Pushkino, Russia, 2013; 208p. [Google Scholar]
- Federation Council. Decision of the Scientific Expert Council under the Chairman of the Federation Council of the Federal Assembly of the Russian Federation. Economic, Budgetary, Financial and Tax Foundations of Federalism; Federation Council: Moscow, Russia, 2016; pp. 187–201. [Google Scholar]
- MacDicken, K. (Ed.) Changes in Global Forest Resources from 1990 to 2015. In Forest Ecology and Management. Science to Sustain the World’s Forests; Elsevier: Amsterdam, The Netherlands, 2015; Volume 352, pp. 1–146. [Google Scholar]
- United Nations. The European Forest Sector Outlook Study II: 2010–2030; United Nations: Geneva, Switzerland, 2011; 108p. [Google Scholar]
- IVUZ. Bulletin of higher educational institutions. For. J. 2019. [Google Scholar]
- Sokolov, V.N. The problem of loess. Soros Educ. J. 1996, 86–93. [Google Scholar]
- Paluch, J.G.; Bartkowicz, L.E. Spatial interactions between Scots pine (Pinus sylvestris L.), common oak (Quercus robur L.) and silver birch (Betula pendula Roth.) as investigated in stratified stands in mesotrophic site conditions. For. Ecol. Manag. 2004, 192, 229–240. [Google Scholar] [CrossRef]
- Atkinson, M.D. Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. J. Ecol. 1992, 80, 837–870. [Google Scholar] [CrossRef]
- Kubentayev, S.A.; Alibekov, D.T.; Perezhogin, Y.V.; Lazkov, G.A.; Kupriyanov, A.N.; Ebel, A.L.; Izbastina, K.S.; Borodulina, O.V.; Kubentayeva, B.B. Revised checklist of endemic vascular plants of Kazakhstan. PhytoKeys 2024, 238, 241–279. [Google Scholar] [CrossRef] [PubMed]
- Storchak, T.V. Birch Forests of the Turgai Depression. Ph.D. Thesis, Kostanay State University, Kostanay, Kazakhstan, 2001. [Google Scholar]
- Ivaschenko, A.A. Flora of Kazakhstan. Encyclopedia/AA Ivaschenko; Almatykitap: Almatym, Kazakhstan, 2009. [Google Scholar]
- Esseen, P.A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal forests. Ecol. Bull. 1997, 46, 16–47. [Google Scholar]
- Orazov, A.; Yermagambetova, M.; Myrzagaliyeva, A.; Mukhitdinov, N.; Tustubayeva, S.; Turuspekov, Y.; Almerekova, S. Plant height variation and genetic diversity between Prunus ledebouriana (Schlecht.) YY Yao and Prunus tenella Batsch based on using SSR markers in East Kazakhstan. PeerJ 2024, 12, e16735. [Google Scholar] [CrossRef]
- Orazov, A.; Myrzagaliyeva, A.; Mukhitdinov, N.; Tustubayeva, S. Callus induction with 6-BAP and IBA as a way to preserve Prunus ledebouriana (Rosaceae), and endemic plant of Altai and Tarbagatai, East Kazakhstan. Biodiversitas J. Biol. Divers. 2022, 23, 6. [Google Scholar] [CrossRef]
- Aidarkhanova, G.S.; Kubentayev, S.A.; Kukhar, Y.V.; Urazalina, A.S. Current state of populations, resources, and biological activity Pulsatilla patens (Ranunculaceae) in Northern Kazakhstan. Biodiversitas 2022, 23, 2311–2320. [Google Scholar] [CrossRef]
- Genievskaya, Y.; Almerekova, S.; Abugalieva, S.; Abugalieva, A.; Sato, K.; Turuspekov, Y. Identification of SNPs associated with grain quality traits in spring barley collection grown in southeastern Kazakhstan. Agronomy 2023, 13, 1560. [Google Scholar] [CrossRef]
- Ozgeldinova, Z.; Mukayev, Z.; Zhanguzhina, A.; Bektemirova, A.; Ulykpanova, M. Spatial and temporal analysis of landscape dynamics in the Kostanay region under anthropogenic impacts. Arch. Environ. Prot. 2023, 49, 80–94. [Google Scholar]
- Zubairov, B.; Lentschke, J.; Schröder, H. Dendroclimatology in Kazakhstan. Dendrochronologia 2019, 56, 125602. [Google Scholar] [CrossRef]
- Imanbayeva, A.; Duisenova, N.; Orazov, A.; Sagyndykova, M.; Belozerov, I.; Tuyakova, A. Study of the Floristic, Morphological, and Genetic (atpF–atpH, Internal Transcribed Spacer (ITS), matK, psbK–psbI, rbcL, and trnH–psbA) Differences in Crataegus ambigua Populations in Mangistau (Kazakhstan). Plants 2024, 13, 1591. [Google Scholar] [CrossRef] [PubMed]
- Bazargaliyeva, A.; Kurmanbayeva, M.; Admanova, G.; Sarzhigitova, A.; Koblanova, S.; Utaubayeva, A.; Gataulina, G.; Kaisagaliyeva, G.; Bissenov, U.; Alzhanova, B. Geobotanical characteristics of plant communities with participation of rare species Alnus glutinosa (L.) Gaertn. Braz. J. Biol. 2024, 84, e281672. [Google Scholar] [CrossRef] [PubMed]
- Ozgeldinova, J.; Mukayev, J.; Turlybekov, K.; Zhanguzhina, A.; Ramazanova, N.; Ulykpanova, M. Post-fire state and restoration of forests in the territory of Kostanay Region. Bull. KazNU Ecol. Ser. 2024, 79, 52–68. [Google Scholar]
- Perezhogin, Y.V. Formation of vegetation cover of Kostanay region (Northern Kazakhstan). Bull. Orenbg. State Univ. 2006, 257–260. [Google Scholar]
- Perezhogin, Y.V. Relics in the flora of Kostanay region (Northern Kazakhstan). Bull. Orenbg. State Univ. 2008, 130–132. [Google Scholar]
- Myrzagaliyeva, A.; Seilkhan, A.; Takirova, M.; Mukhtubayeva, S.; Zhumagul, M.; Nursafina, A.; Bolgibayeva, A.; Makhambetov, M. Morphological variability of the rare species Linaria cretacea in the conditions of the chalk hills in North-Western Kazakhstan. Casp. J. Environ. Sci. 2023, 21, 1273–1278. [Google Scholar]
- Vesselova, P.; Makhmudova, K.; Kudabayeva, G.; Osmonali, B.; Mikhalev, V. Current Growth Conditions of Populus Diversifolia Schrenk and Populus Pruinosa Schrenk in the Syr-Darya Valley. J. Biological Sci. 2022, 22, 425–438. [Google Scholar]
- Sukachev, V.N.; Zonn, S.V.; Motovilov, G.P. Methodical Instructions for Studying Forest Types; Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1957. [Google Scholar]
- Sukachev, V.N. Forest Biogeocenology as a Theoretical Basis for Forestry and Forest Management; Issues of Forest Science and Forestry; Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1960. [Google Scholar]
- Pott, R. Phytosociology: A modern geobotanical method. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2011, 145 (Suppl. 1), 9–18. [Google Scholar] [CrossRef]
- Kafle, G. Diagnostic Horizons of Soils in the World; Academia Letters: San Francisco, CA, USA, 2022. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2024. Available online: http://www.plantsoftheworldonline.org (accessed on 1 September 2024).
- Pavlov, N.B. Flora of Kazakhstan; Academy of Sciences of the Kazakh: Almaty, Kazakhstan, 1961. [Google Scholar]
- Komarov, V.L. Flora SSSR; Israel Program for Scientific Translations: Jerusalem, Israel, 1941; pp. 522–547. (In Russian) [Google Scholar]
- Malyschev, L.I.; Peschkova, G.A. Flora of Siberia; Nauka: Novosibirsk, Russia, 1997. [Google Scholar]
- Vdovina, T.A.; Isakova, E.A.; Lagus, O.A.; Sumbembayev, A.A. Selection assessment of promising forms of natural Hippophae rhamnoides (Elaeagnaceae) populations and their offspring in the Kazakhstan Altai Mountains. Biodiversitas J. Biol. Divers. 2024, 25, 4. [Google Scholar] [CrossRef]
- Wikum, D.A.; Shanholtzer, G.F. Application of the Braun-Blanquet cover-abundance scale for vegetation analysis in land development studies. Environ. Manag. 1978, 2, 323–329. [Google Scholar] [CrossRef]
- Vetchinnikova, L.V.; Titov, A.F. Genus Betula L.: Species-Specific Population-Genetic Features and Taxonomy Problems. Biol. Bull. Rev. 2023, 13 (Suppl. 3), S377–S391. [Google Scholar] [CrossRef]
- Mesyats, S.; Novozhilova, M.; Volkova, E. Investigation of the forest stage of succession of sown phytocenosis when restoring disturbed lands with the aim to give them back to biosphere foundation. Int. Multidiscip. Sci. GeoConf. SGEM 2016, 3, 469–476. [Google Scholar]
- Gritsenko, V.V.; Shynder, O.I. Flora of the botanical-geographical plot “Steppes of Ukraine” in the MM Gryshko National Botanical Garden of the NAS of Ukraine. Plant Introd. 2022, 95, 96. [Google Scholar] [CrossRef]
- Johansson, T. Site index conversion equations for Picea abies and five broadleaved species in Sweden: Alnus glutinosa, Alnus incana, Betula pendula, Betula pubescens and Populus tremula. Scand. J. For. Res. 2006, 21, 14–19. [Google Scholar] [CrossRef]
- Zhuk, A.; Veinberga, I.; Daugavietis, M.; Rungis, D. Cross-species amplification of Betula pendula Roth. simple sequence repeat markers in Alnus species. Balt. For. 2008, 14, 116–121. [Google Scholar]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef]
- Zatybekov, A.; Yermagambetova, M.; Genievskaya, Y.; Didorenko, S.; Abugalieva, S. Genetic Diversity Analysis of Soybean Collection Using Simple Sequence Repeat Markers. Plants 2023, 12, 3445. [Google Scholar] [CrossRef]
- Zeinullina, A.; Zargar, M.; Dyussibayeva, E.; Orazov, A.; Zhirnova, I.; Yessenbekova, G.; Zotova, L.; Rysbekova, A.; Hu, Y.G. Agro-Morphological Traits and Molecular Diversity of Proso Millet (Panicum miliaceum L.) Affected by Various Colchicine Treatments. Agronomy 2023, 13, 2973. [Google Scholar] [CrossRef]
- Zargar, M.; Dyussibayeva, E.; Orazov, A.; Zeinullina, A.; Zhirnova, I.; Yessenbekova, G.; Rysbekova, A. Microsatellite-based genetic diversity analysis and population structure of Proso Millet (Panicum miliaceum L.) in Kazakhstan. Agronomy 2023, 13, 2514. [Google Scholar] [CrossRef]
- Myrzagaliyeva, A.; Irsaliyev, S.; Tustubayeva Sh Samarkhanov, T.; Orazov, A.; Alemseitova, Z. Natural resources of Rhaponticum carthamoides in the Territory of Tarbagatai State National Nature Park. Diversity. Diversity, 2024; in press. [Google Scholar]
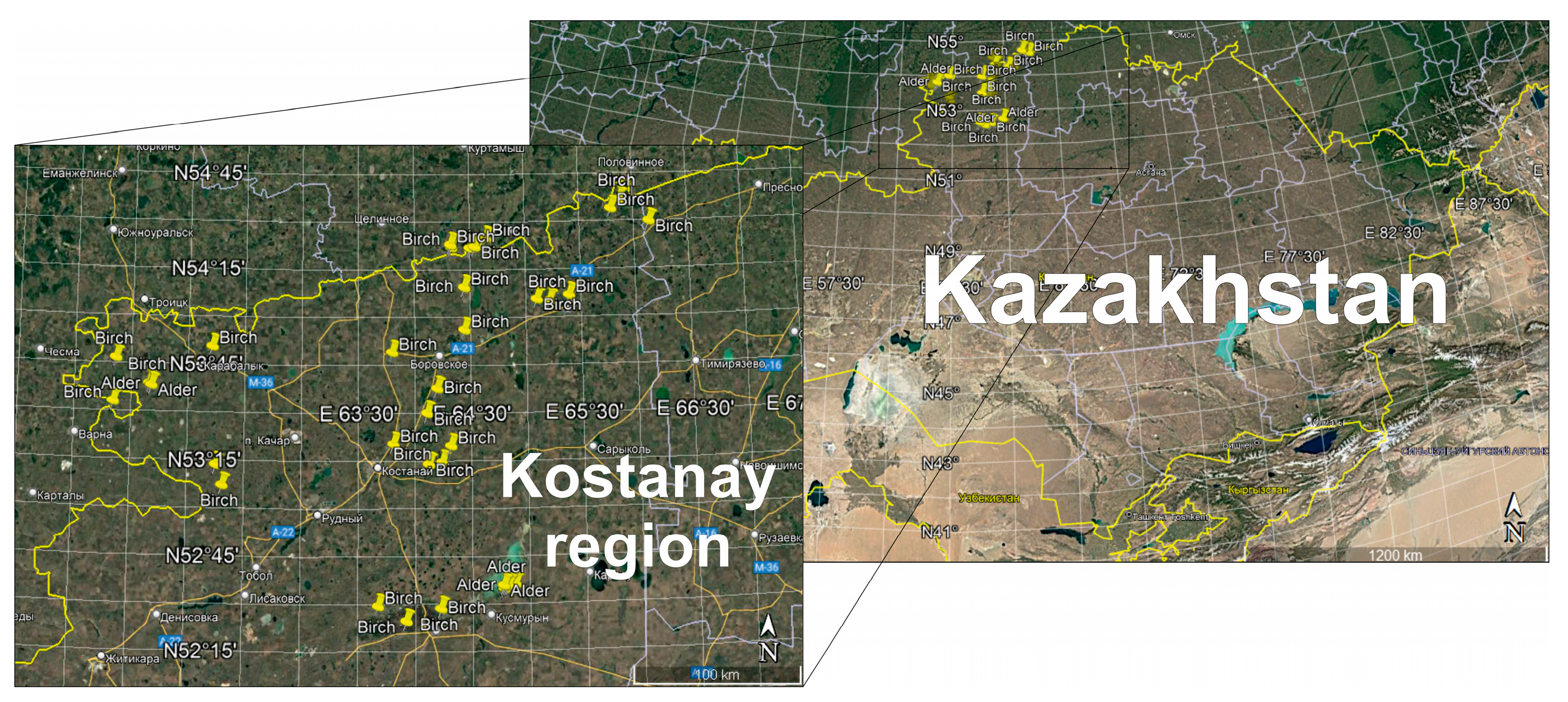
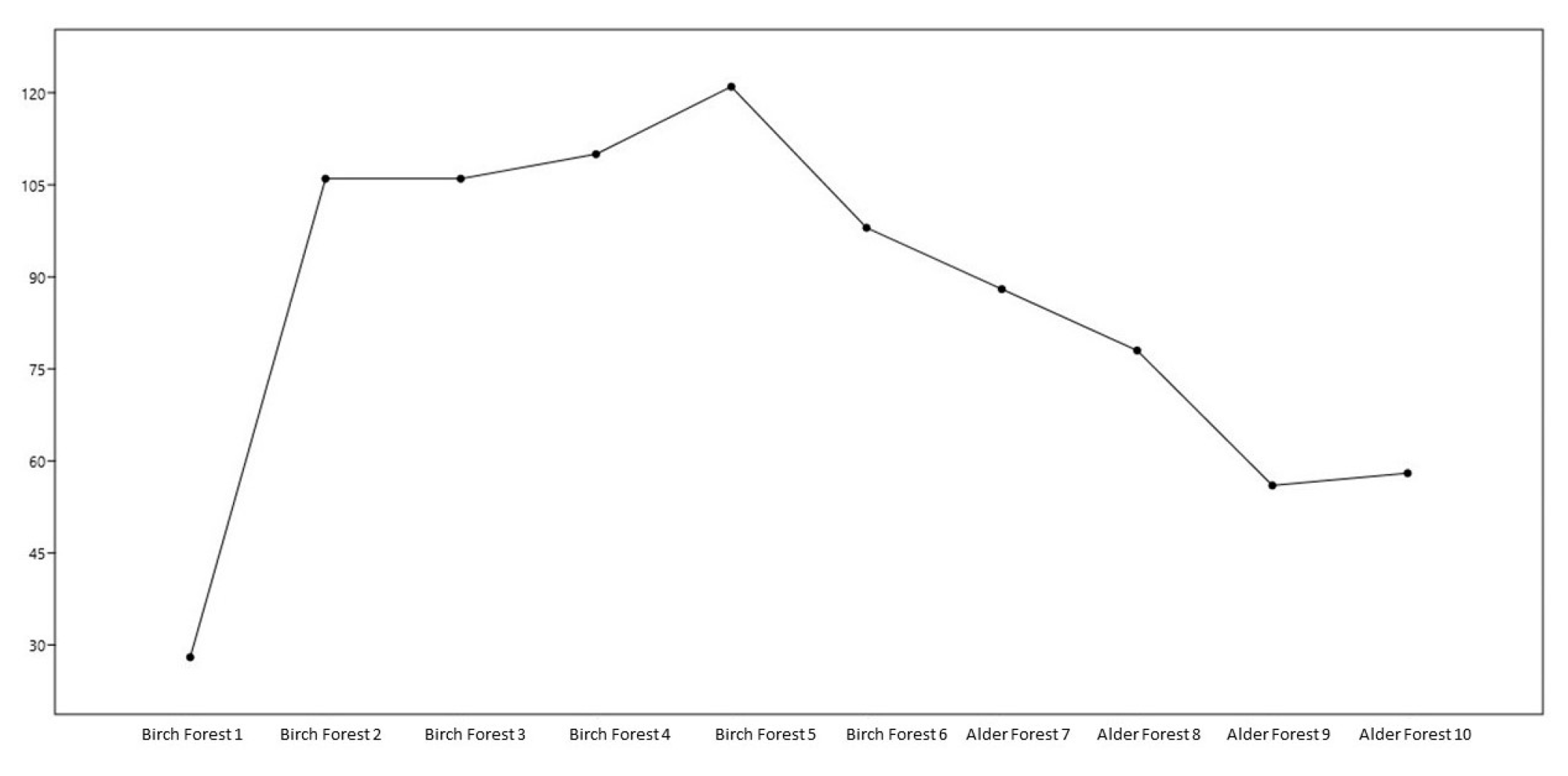
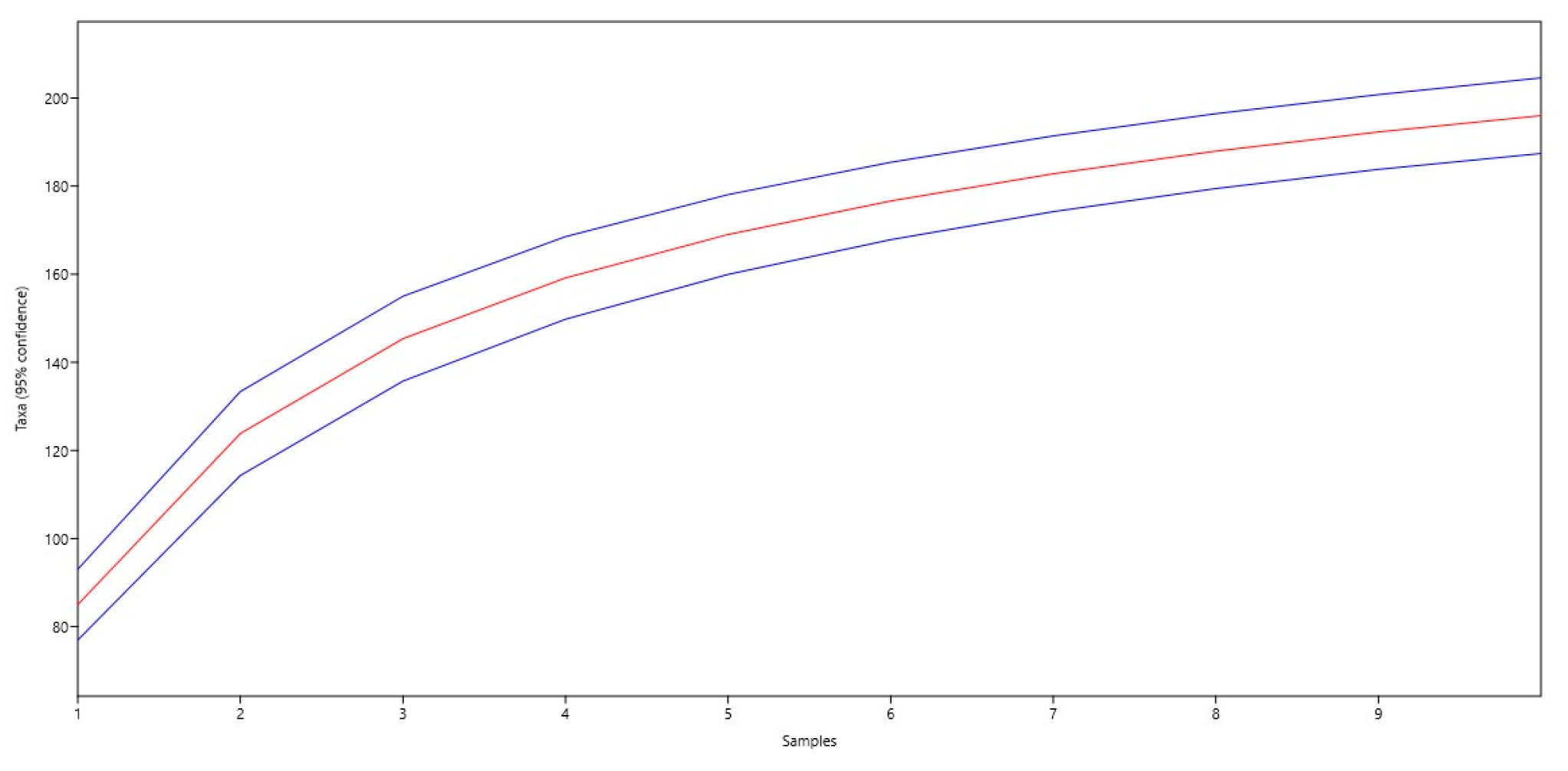
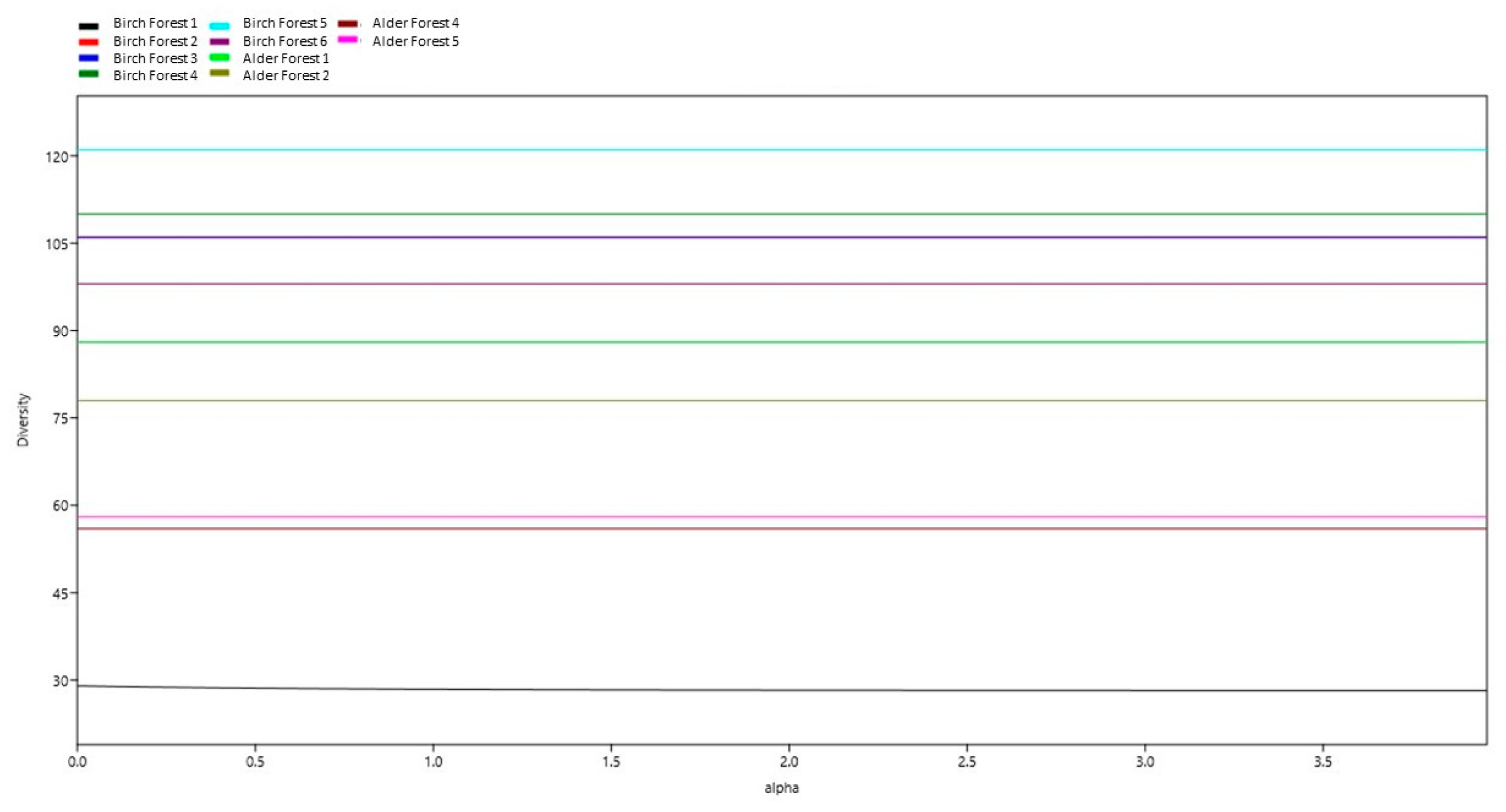
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).