
Taxonomic Studies on Five Species of Sect. Tuberculata (Camellia L.) Based on Morphology, Pollen Morphology, and Molecular Evidence

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Materials
2.2. Experimental Research Method
2.2.1. Data Statistics
2.2.2. Micromorphological of Pollen
2.2.3. Chloroplast DNA Acquisition and Genome Assembly
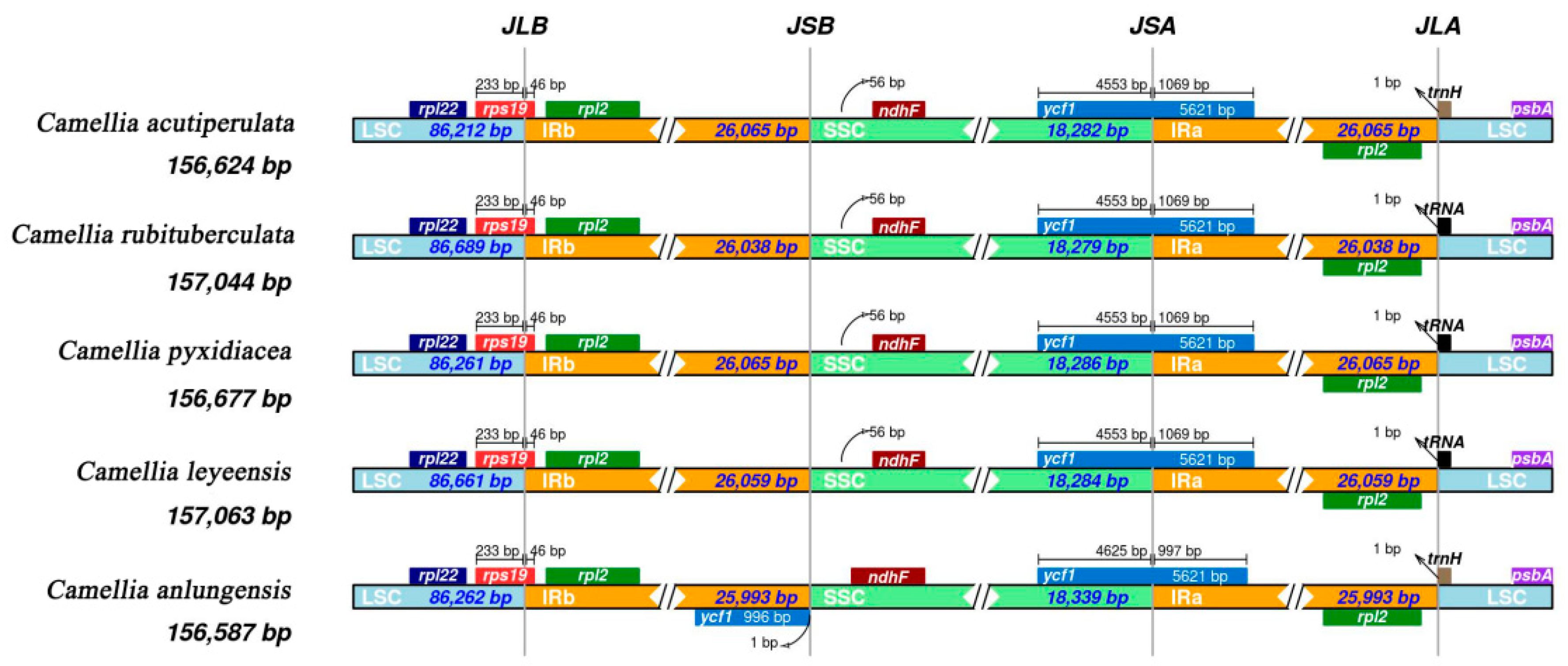
2.2.4. Expansion and Contraction of the IR Boundary
2.2.5. Phylogenetic Tree
3. Results
3.1. Morphological Studies
3.2. Micromorphological Characteristics of Pollen
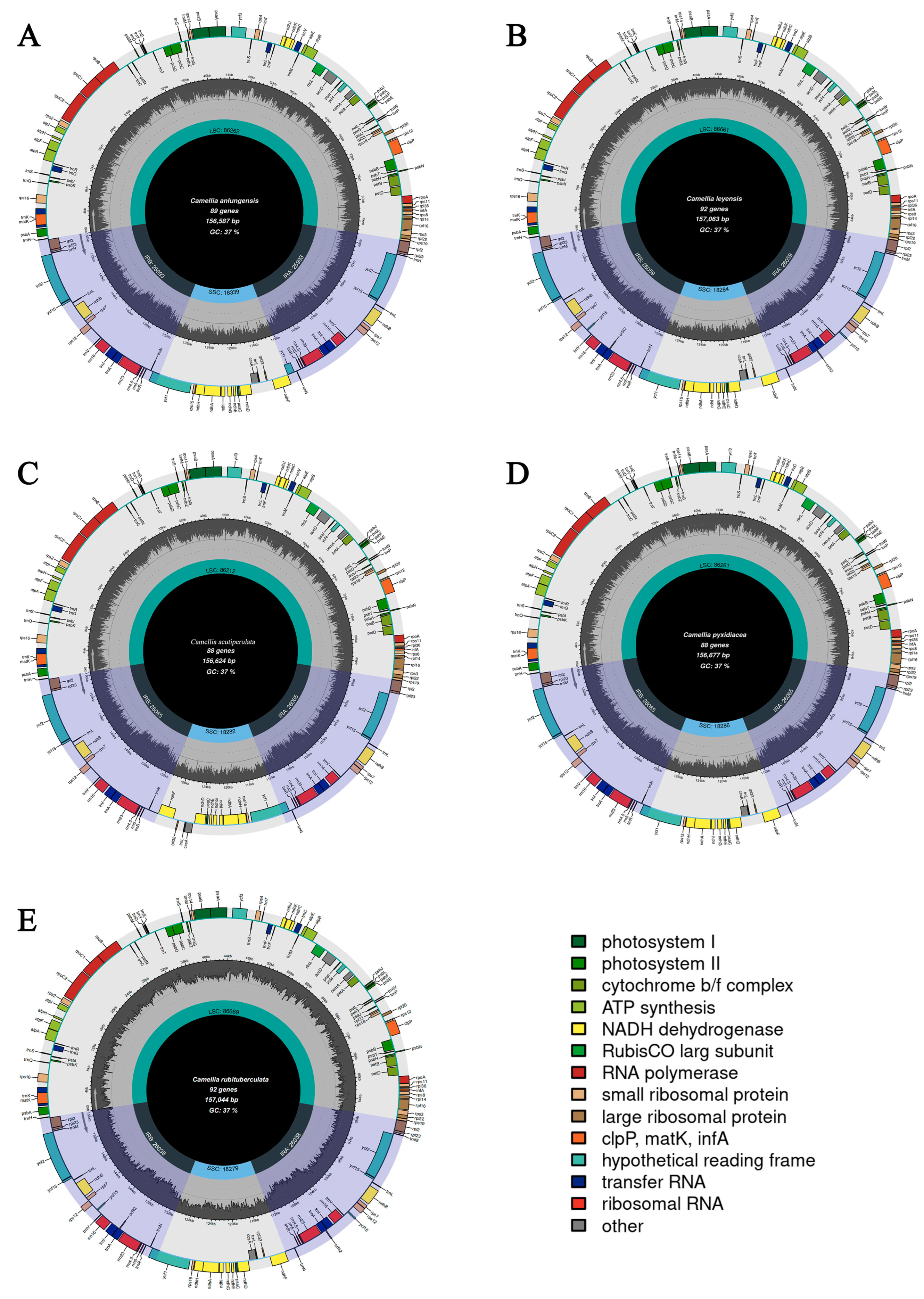
3.3. Features of Chloroplast Genomes
3.4. Phylogenetic Analysis
3.5. Taxonomic Treatment
- Camellia anlungensis Chang
- Camellia autoregulates Chang and C. X. Ye
- Camellia pyxidiacea Xu, F. P. Chen and C. Y. Deng
- Camellia rubituberculata Chang
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chang, H.T. A Taxonomy of the Genus Camellia; Editorial Department of Journal of Sun Yatsen University: Guangzhou, China, 1981; pp. 47–52. [Google Scholar]
- Chien, S.S. Four new ligneous plants of Szechuan. Contrib. Biol. Lab. Sci. Soc. China Bot. 1939, 12, 89–100. [Google Scholar]
- Min, T.L.; Zhong, Y.C. A revision of genus Camellia sect. Tuberculata. Acta Bot. Yunnanica 1993, 15, 123–130. [Google Scholar]
- Min, T.L. A systematic synopsis of the genus Camellia. Acta Bot. Yunnanica 1999, 21, 149–159. [Google Scholar]
- Chang, H.T.; Ren, S.X. Diagnosis on the systematic development of Camellia VI. revised on sect. Tuberculata of Camellia. Suppl. J. Sun Yatsen Univ. 1996, 2, 55–60. [Google Scholar]
- Chang, H.T. New record of Camellia from South China. Acta Sci. Nat. Univ. Sun Yatseni 1984, 23, 77–82. [Google Scholar]
- Sealy, J.R. A Revision of the Genus Camellia; The Royal Horticulture Society: London, UK, 1958; pp. 1–239. [Google Scholar]
- Jiang, B.; Peng, Q.F.; Shen, Z.G.; Moller, M.; Pi, E.X.; Lu, H.F. Taxonomic treatments of Camellia (Theaceae) species with secretory structures based on integrated leaf characters. Plant Syst. Evol. 2010, 290, 1–20. [Google Scholar] [CrossRef]
- Jiang, Z.D. Preliminary Study of Molecular Phylogenetics and Biogeography of the Genus Camellia L. Based on Chloroplast DNA. Ph.D. Thesis, Zhejiang Sci-Tech University, Hangzhou, China, 2017. [Google Scholar]
- Wu, Q.; Tong, W.; Zhao, H.J.; Ge, R.H.; Li, R.P.; Huang, J.; Li, F.D. Comparative transcriptomic analysis unveils the deep phylogeny and secondary metabolite evolution of 116 Camellia plants. Plant J. 2022, 111, 406–421. [Google Scholar] [CrossRef]
- Xiao, X. Revision of the Classification and Comprehensive Evaluation of the Ornamental Value of Plants in the Camellia L. Sect. Tuberculate. Master’s Thesis, Guizhou University, Guiyang, China, 2023. [Google Scholar]
- Ran, Z.H.; Li, Z.; Xiao, X.; An, M.T.; Yan, C. Complete chloroplast genomes of 13 species of sect. Tuberculata Chang (Camellia L.): Genomic features, comparative analysis, and phylogenetic relationships. BMC Genom. 2024, 25, 108. [Google Scholar] [CrossRef]
- Lin, M.J.; Lu, Q.M. New record of Guizhou Camellia. Act. Sci. Nat. Univ. Sunyats. 1984, 2, 81–83. [Google Scholar]
- Xu, Z.R.; Chen, F.; Deng, C.Y. A new species of sect. Tuberculata. Guihaia 1987, 19–21. [Google Scholar]
- Chang, H.T.; Ren, S.X. A classification on the section Tuberculata of Camellia. Act. Sci. Nat. Univ. Sunyats. 1991, 30, 86–91. [Google Scholar]
- Erdtman, G. Handbook of Palynology; Science Press: Beijing, China, 1978. [Google Scholar]
- Wang, K.F.; Wang, X.Z. Introduction to Sporology; Peking University Press: Beijing, China, 1983. [Google Scholar]
- Halbritter, D.D.; Ulrich, D.S.; Grímsson, D.F.; Weber, P.D.; Zetter, P.D.; Hesse, P.D.; Buchner, D.R.; Svojtka, M.M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Cambridge International Law Journal: Cambridge, UK, 2018. [Google Scholar]
- Li, J.L.; Wang, S.; Yu, J.; Wang, L.; Zhou, S.L. A modified CTAB protocol for plant DNA extraction. Chin. Bull. Bot. 2013, 48, 72–78. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. Organellar Genome DRAW (OGDRAW): A tool for the easy generation of highquality custom graphical maps of plastid and mitochondrial genome. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Amyiryousefi, A.; Hyvonen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT: Iterative refinement and additional methods. Methods Mol. Biol. 2014, 1079, 131–146. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQTREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5, an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Efroni, I.; Eshed, Y.; Lifschitz, E. Morphogenesis of simple and compound leaves: A critical review. Plant Cell 2010, 22, 1019–1032. [Google Scholar] [CrossRef]
- Wei, Z.X.; Zavada, M.S.; Min, T.L. Pollen morphology of Camellia (Theaceae) and ITS taxonomic significance. Acta Bot. Yunnanica. 1992, 14, 275–282. [Google Scholar]
- Hu, Z.M.; Zhao, C.H.; Zhao, Y.Y.; Liu, J.X. Pollen morphology of Liliaceae and its systematic significance. Palynology 2021, 45, 531–568. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Location | Specimen Number | Elevation (m) | Longitude and Latitude |
|---|---|---|---|---|
| C. anlungensis | Wangmo County, Guizhou, China | GZAC, LZ20211204 | 449 | 106°16′40.69″ E, 25°2′51.65″ N |
| C. leyeensis | Leye County, Guangxi, China | GZAC, LZ20210413 | 684 | 106°17′31.48″ E, 24°51′29.98″ N |
| C. acutiperulata | Longlin County, Guangxi, China | GZAC, LZ20221103 | 851 | 104°52′39.66″ E, 24°39′31.59″ N |
| C. pyxidiacea | Xingyi City, Guizhou, China | GZAC, LZ20211204 | 837 | 104°32′18.18″ E, 24°47′53.12″ N |
| C. rubituberculata | Xingren County, Guizhou, China | GZAC, LZ20210411 | 1297 | 105°21′08.10″ E, 25°43′50.08″ N |
| Trait | C. anlungensis | C. leyeensis | C. acutiperulata | C. pyxidiacea | C. rubituberculata |
|---|---|---|---|---|---|
| Leaf type | Leaves thin, leathery, obovate | Leaves thin, leathery, obovate | Leaves thin, coriaceous, elliptic | Leaves thin, coriaceous, elliptic | Leaves thin coriaceous, elliptic |
| Length (cm) | 9.14–12.54 | 9.95–12.31 | 10.28–13.69 | 8.14–13.08 | 10.08–11.43 |
| Width (cm) | 2.78–4.45 | 2.79–3.85 | 3.32–4.73 | 3.94–4.88 | 3.51–4.82 |
| Flower type | Obovate | Obovate | Elliptic | Elliptic | Elliptic |
| Color | White | White | White | Pink petal apexes | Red |
| Length (cm) | 2.08–2.95 | 2.27–3.10 | 3.44–4.01 | 2.43–2.88 | 2.79–3.25 |
| Width (cm) | 0.97–1.54 | 0.93–1.63 | 1.35–1.94 | 0.95–1.58 | 2.15–3.24 |
| Number of petals | 11–13 | 10–12 | 9–12 | 6–9 | 6–8 |
| Sepal | Ovate | Ovate | Ovate, apex pointed, margin membranous | Ovate | Semicircular |
| Number of calyces | 5–7 | 5–8 | 5–7 | 6–8 | 7–9 |
| Fruit shape | Capsule verrucose, subglobose, epidermis with verrucose projections | Capsule verrucose, subglobose, epidermis with verrucose projections | Capsule verrucose, subglobose, testa transversely lobed | Fruit-compressed globose, skin with verrucose protuberance and obvious parting | Capsule verrucose, globose, seed coat shallowly undulated |
| Diameter (cm) | 1.4–2.04 | 1.35–1.9 | 1.3–2.1 | 2.65–3.81 | 4.84–5.18 |
| Shell thickness (mm) | 6.3–7.7 | 5.8–7.3 | 3.1–4.3 | 3.2–0.67 | 12.2–14.1 |
| Species | Pollen Shape | Polar View | Equatorial View | Polar Axis (P)/μm | Equatorial Axis E/μm | Germination Pores | Pollen Aperture | Pollen Wall Ornamentation | Ridge | |
|---|---|---|---|---|---|---|---|---|---|---|
| Long/μm | Wide/μm | |||||||||
| C. anlungensis | oblate spherical | Triangular | Oval | 30.21–32.27 | 36.09–37.88 | 26.83–27.75 | 7.14–7.75 | Colpus | Uneven | Wavy |
| C. leyeensis | oblate spherical | Triangular | Oval | 29.41–31.67 | 34.74–36.51 | 26.94–27.73 | 6.73–7.42 | Colpus | Uneven | Wavy |
| C. acutiperulata | oblate spherical | Triangular | Oval | 27.54–28.51 | 33.66–35.47 | 24.54–25.12 | 6.71–7.13 | Colpus | Uneven | Wavy |
| C. pyxidiacea | oblate spherical | Triangular | Oval | 30.91–31.82 | 31.87–33.21 | 22.11–23.17 | 6.59–6.97 | Colpus | Uneven | Wavy |
| C. rubituberculata | oblate spherical | Oval | Oval | 28.27–29.84 | 32.69–34.57 | 26.85–27.35 | 8.28–8.74 | Colpus | Uneven | Wavy |
| C. acutiperulata | C. pyxidiacea | C. anlungensis | C. rubituberculata | C. leyeensis | |
|---|---|---|---|---|---|
| Genome size (bp) | 156,624 | 156,677 | 156,587 | 157,044 | 157,063 |
| GC (%) | 37.33 | 37.33 | 37.33 | 37.31 | 37.30 |
| LSC size (bp) | 86,212 | 86,261 | 86,262 | 86,689 | 86,661 |
| SSC size (bp) | 18,286 | 18,283 | 18,281 | 18,279 | 18,276 |
| IR size (bp) | 52,130 | 52,130 | 51,986 | 52,076 | 52,118 |
| GC in LSC (%) | 35.36 | 35.35 | 35.31 | 35.31 | 35.31 |
| GC in SSC (%) | 30.59 | 30.62 | 30.60 | 30.62 | 30.60 |
| GC in IR (%) | 42.95 | 42.95 | 42.96 | 42.98 | 42.96 |
| GC in CDS (%) | 37.61 | 37.53 | 37.54 | 37.65 | 37.55 |
| 1st position GC (%) | 45.37 | 45.19 | 45.24 | 45.42 | 45.26 |
| 2nd position GC (%) | 38.04 | 37.94 | 37.97 | 38.00 | 37.97 |
| 3rd position GC (%) | 29.43 | 29.43 | 29.40 | 29.53 | 29.40 |
| Length of CDS | 79,500 | 79,767 | 79,671 | 80,155 | 80,175 |
| Number of genes | 130 | 132 | 134 | 136 | 136 |
| Number of CDS | 87 | 87 | 89 | 91 | 91 |
| Number of tRNAs | 35 | 37 | 37 | 37 | 37 |
| Number of rRNAs | 8 | 8 | 8 | 8 | 8 |
| GenBank ID | OQ556869 | OP058659 | NC_050354 | MZ766253 | OK046127 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Li, Z.; Ran, Z.; Yan, C.; Tang, M.; Huang, L. Taxonomic Studies on Five Species of Sect. Tuberculata (Camellia L.) Based on Morphology, Pollen Morphology, and Molecular Evidence. Forests 2024, 15, 1718. https://doi.org/10.3390/f15101718
Xiao X, Li Z, Ran Z, Yan C, Tang M, Huang L. Taxonomic Studies on Five Species of Sect. Tuberculata (Camellia L.) Based on Morphology, Pollen Morphology, and Molecular Evidence. Forests. 2024; 15(10):1718. https://doi.org/10.3390/f15101718
Chicago/Turabian StyleXiao, Xu, Zhi Li, Zhaohui Ran, Chao Yan, Ming Tang, and Lang Huang. 2024. "Taxonomic Studies on Five Species of Sect. Tuberculata (Camellia L.) Based on Morphology, Pollen Morphology, and Molecular Evidence" Forests 15, no. 10: 1718. https://doi.org/10.3390/f15101718