Effects of Root Pruning and Size on Growth Traits of Hybrid Poplar Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seedling Preparation and Selection
2.2. Root Pruning Procedure
2.3. Indicator Measurements
2.3.1. Early Planting Performance and Key Phenology
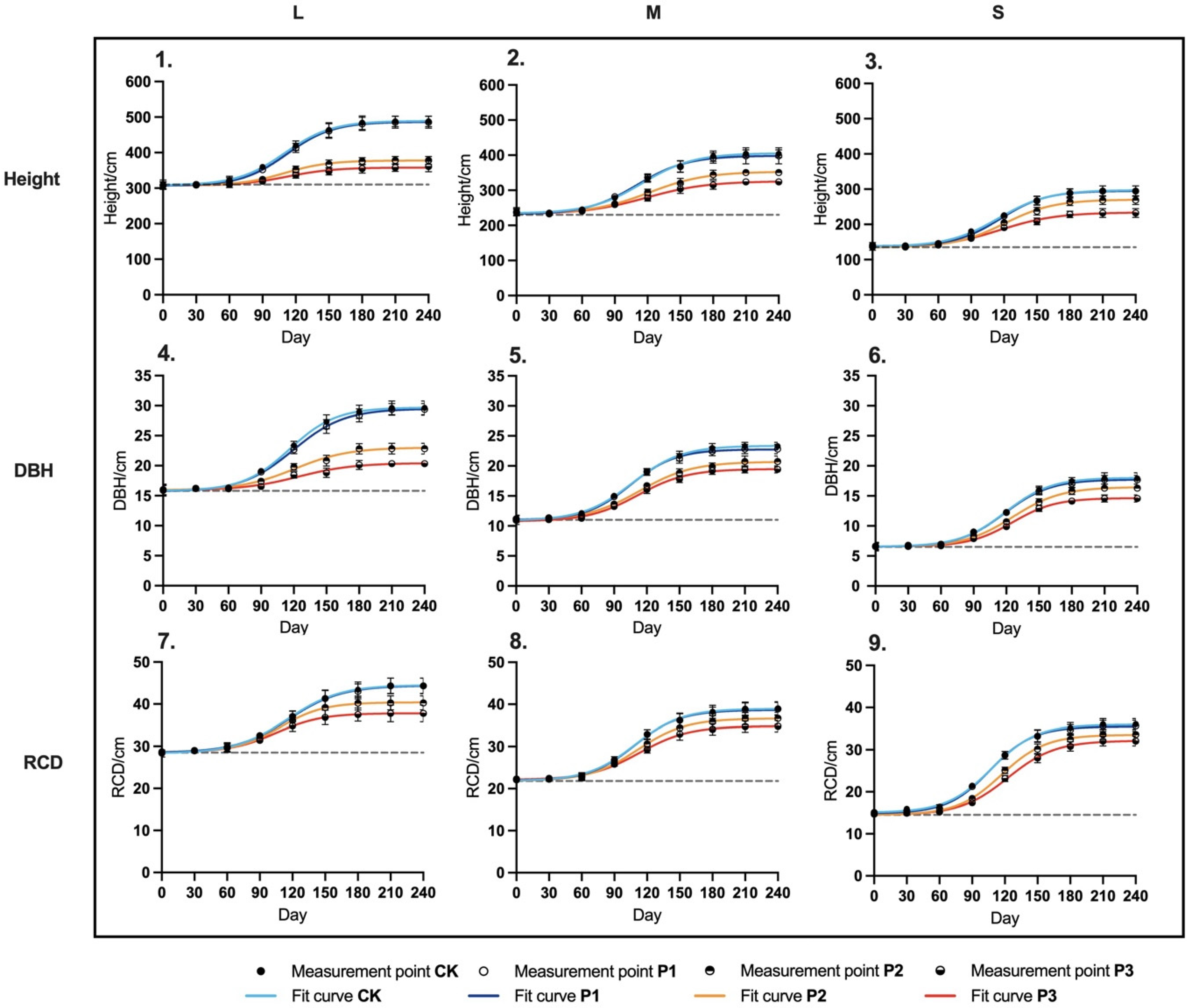
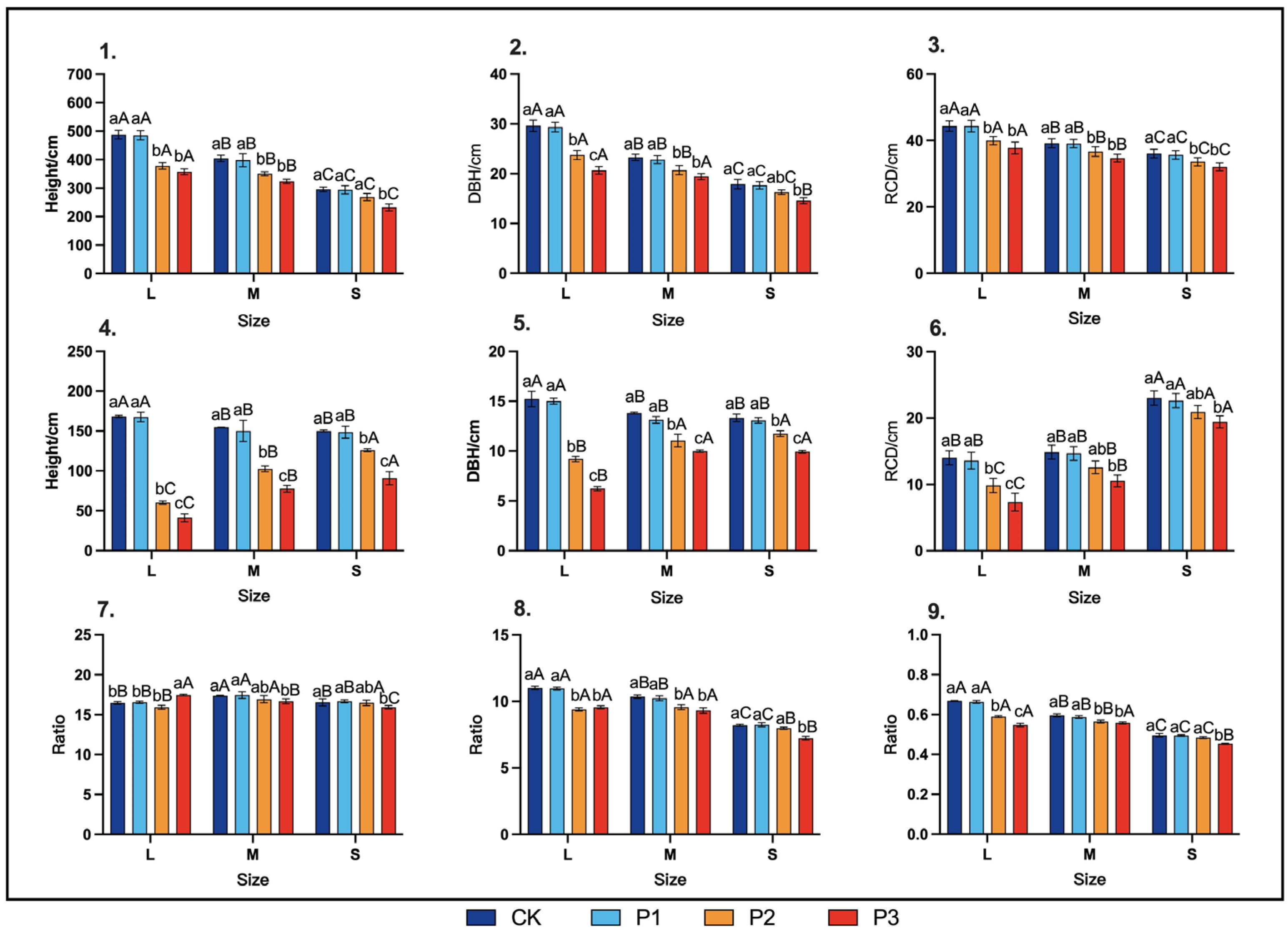
2.3.2. Responses of H, DBH, and RCD
2.4. Statistical Analysis
3. Results
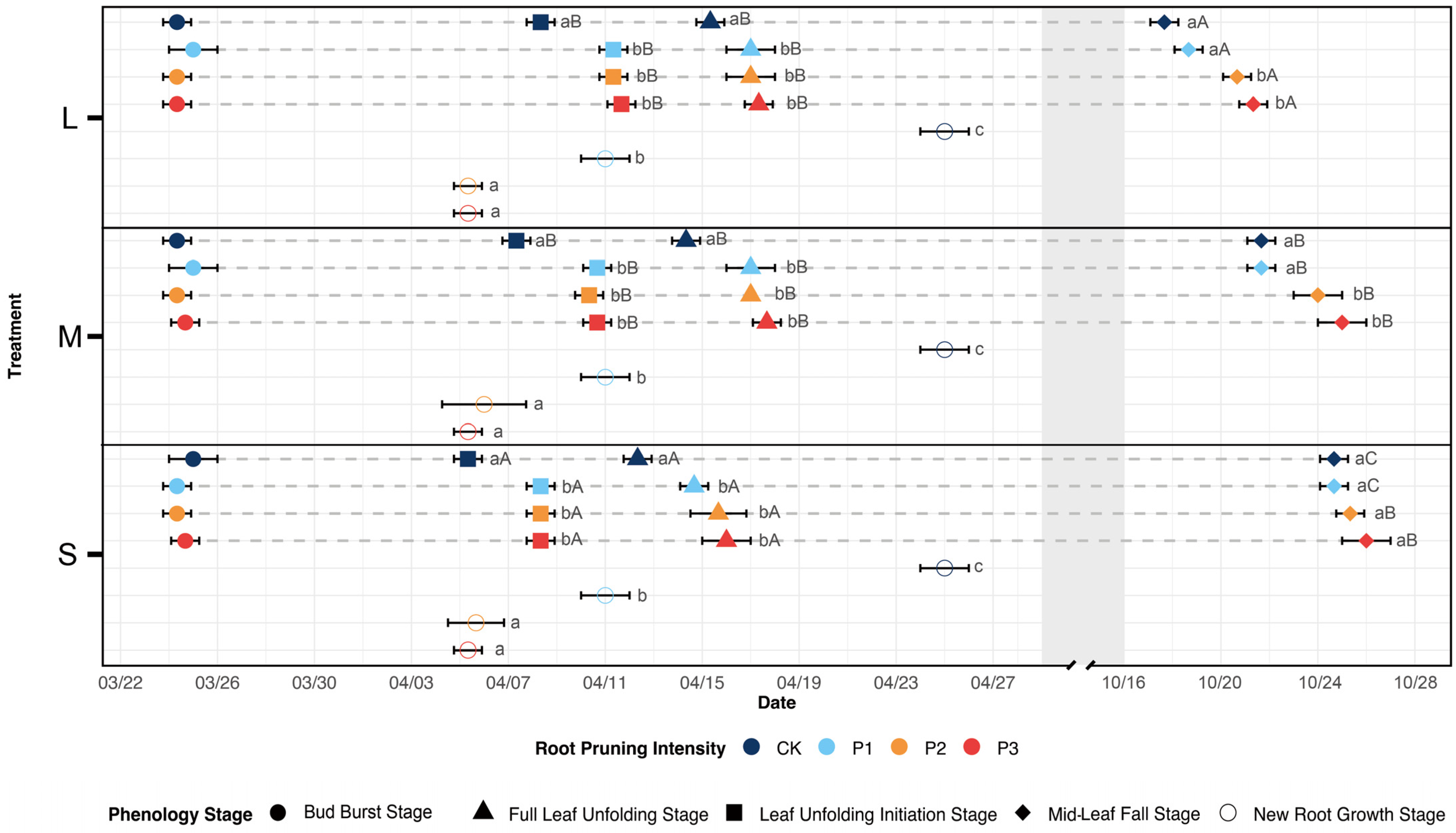
3.1. Key Phenological Traits
3.1.1. Survival Rate and Dieback
3.1.2. Key Leaf Traits and Root Emergence
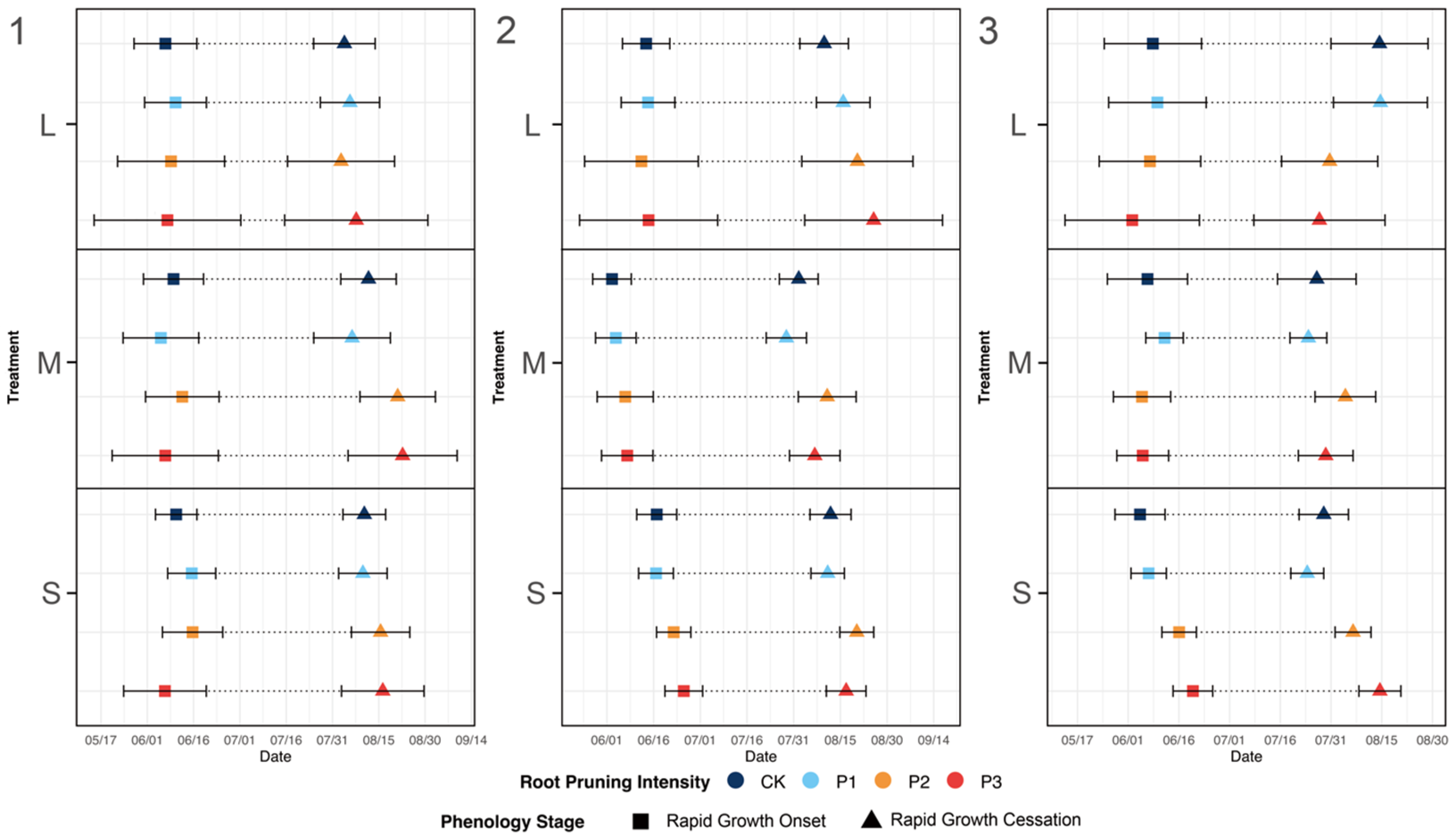
3.1.3. Key Stem Phenological Traits
3.2. Morphological Traits
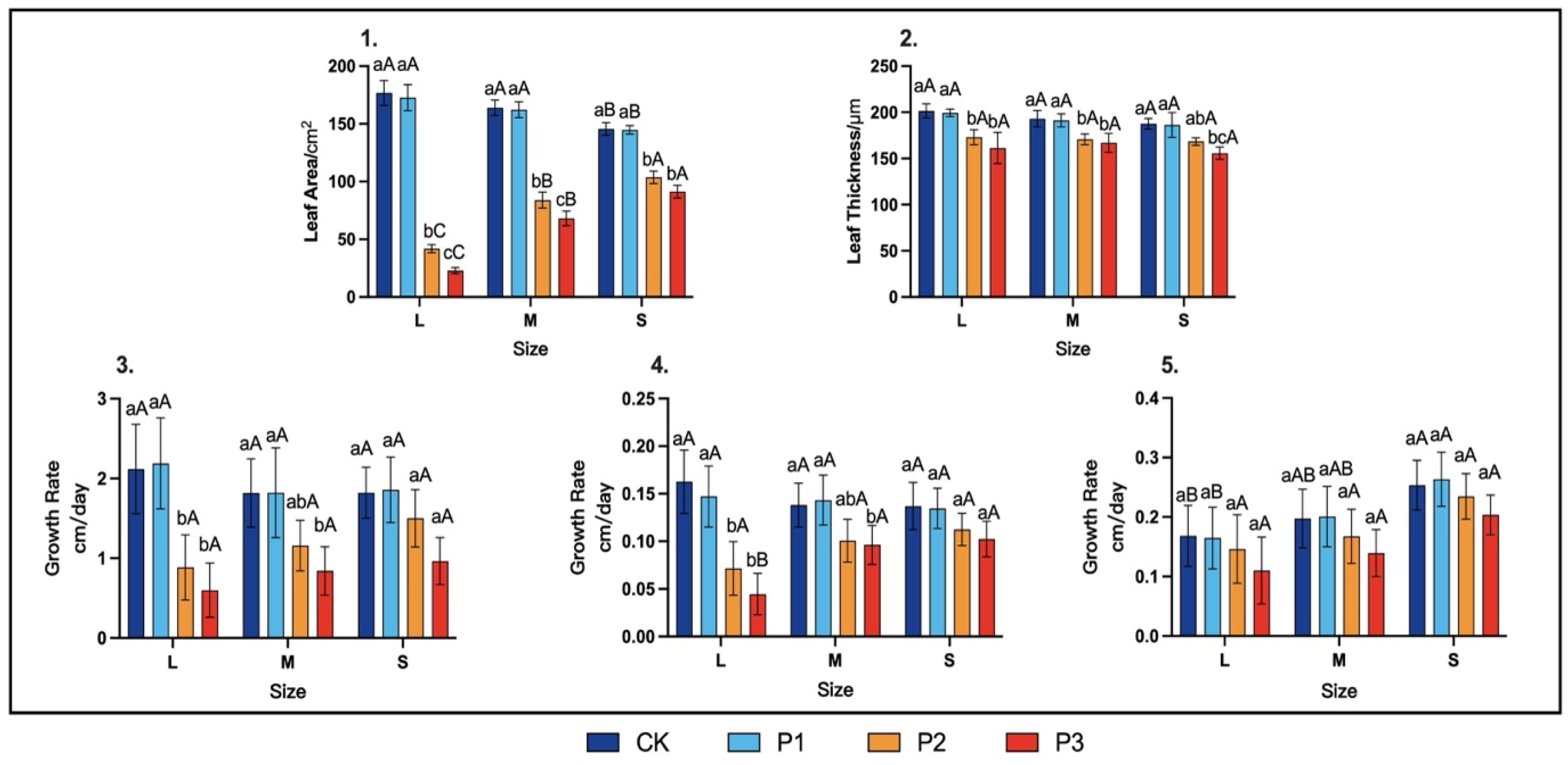
3.2.1. Leaf Traits and Stem Growth Rate during the Rapid Growth Period
3.2.2. End-of-Season Growth Traits
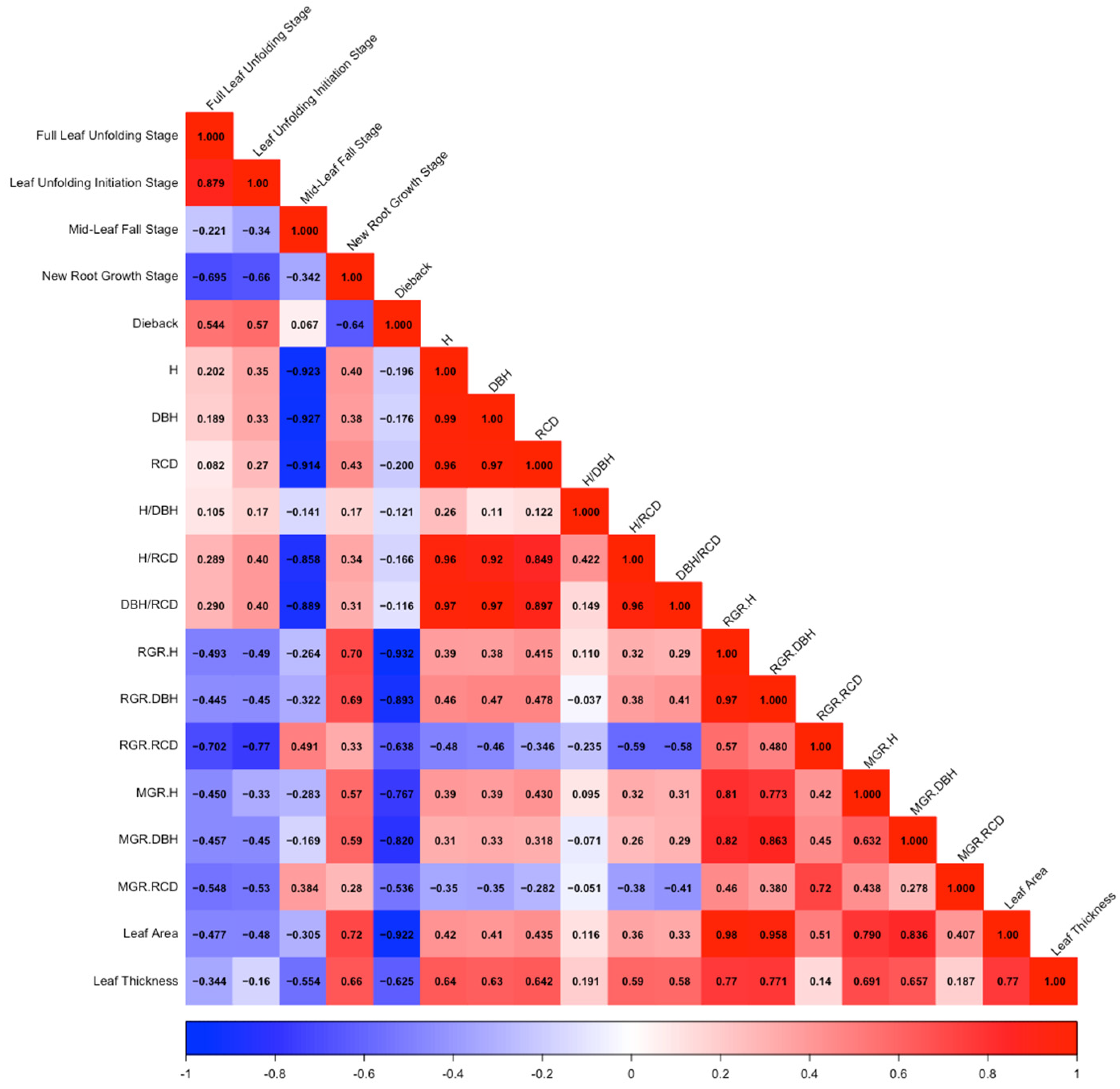
3.3. PCA and Correlation Analysis
4. Discussion
4.1. Effects of Root Pruning Intensity and Seedling Sizes on Key Phenological Traits
4.2. Effects of Root Pruning Intensity and Seedling Sizes on Morphological Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baghalian, K.; Hajirezaei, M.-R.; Lawson, T. Current and Future Perspectives for Controlled Environment Agriculture (CEA) in the 21st Century. Front. Plant Sci. 2023, 14, 1334641. [Google Scholar] [CrossRef] [PubMed]
- Na, S.-J.; Lee, D.-H.; Kim, I.-S. Influence of Initial Seedling Size and Root Pruning Intensity on Growth of Transplanting Seedling of Quercus Acutissma. Korean J. Plant Resour. 2013, 26, 709–717. [Google Scholar] [CrossRef]
- Farmer, J.W.; Pezeshki, S.R. Effects of Periodic Flooding and Root Pruning on Quercus Nuttallii Seedlings. Wetl. Ecol. Manag. 2004, 12, 205–214. [Google Scholar] [CrossRef]
- Toca, A.; Moler, E.; Nelson, A.; Jacobs, D.F. Environmental Conditions in the Nursery Regulate Root System Development and Architecture of Forest Tree Seedlings: A Systematic Review. New For. 2022, 53, 1113–1143. [Google Scholar] [CrossRef]
- Wan, F.; Ross-Davis, A.L.; Davis, A.S.; Song, X.; Chang, X.; Zhang, J.; Liu, Y. Nutrient Retranslocation in Larix Principis-Rupprechtii Mayr Relative to Fertilization and Irrigation. New For. 2021, 52, 69–88. [Google Scholar] [CrossRef]
- Hansen, E.A. Planting Date Affects Survival and Height Growth of Hybrid Poplar. For. Chron. 1986, 62, 164–169. [Google Scholar] [CrossRef]
- Löf, M.; Birkedal, M. Direct Seeding of Quercus robur L. for Reforestation: The Influence of Mechanical Site Preparation and Sowing Date on Early Growth of Seedlings. For. Ecol. Manag. 2009, 258, 704–711. [Google Scholar] [CrossRef]
- Palacios, G.; Navarro Cerrillo, R.M.; del Campo, A.; Toral, M. Site Preparation, Stock Quality and Planting Date Effect on Early Establishment of Holm Oak (Quercus ilex L.) Seedlings. Ecol. Eng. 2009, 35, 38–46. [Google Scholar] [CrossRef]
- Vihera-Aarnio, A.; Hakkinen, R.; Partanen, J.; Luomajoki, A.; Koski, V. Effects of Seed Origin and Sowing Time on Timing of Height Growth Cessation of Betula Pendula Seedlings. Tree Physiol. 2005, 25, 101–108. [Google Scholar] [CrossRef]
- Mechergui, T.; Pardos, M.; Jacobs, D.F. Effect of Acorn Size on Survival and Growth of Quercus suber L. Seedlings under Water Stress. Eur. J. For. Res. 2021, 140, 175–186. [Google Scholar] [CrossRef]
- Cuesta, B.; Villar-Salvador, P.; Puértolas, J.; Jacobs, D.F.; Rey Benayas, J.M. Why Do Large, Nitrogen Rich Seedlings Better Resist Stressful Transplanting Conditions? A Physiological Analysis in Two Functionally Contrasting Mediterranean Forest Species. For. Ecol. Manag. 2010, 260, 71–78. [Google Scholar] [CrossRef]
- Mocko, K.; Jones, C.S. Do Seedlings of Larger Geophytic Species Outperform Smaller Ones When Challenged by Drought? Am. J. Bot. 2021, 108, 320–333. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Wiley, E.T.; Solarik, K.A.; Kulbaba, S.P.; Goeppel, A.E. The Importance of Initial Seedling Characteristics in Controlling Allocation to Growth and Reserves under Different Soil Moisture Conditions. Forests 2023, 14, 796. [Google Scholar] [CrossRef]
- Ward, J.S.; Gent, M.P.N.; Stephens, G.R. Effects of Planting Stock Quality and Browse Protection-Type on Height Growth of Northern Red Oak and Eastern White Pine. For. Ecol. Manag. 2000, 127, 205–216. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Seedling Quality: History, Application, and Plant Attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of Root Growth in Overcoming Planting Stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why Seedlings Grow: Influence of Plant Attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F. Quantifying Root System Quality of Nursery Seedlings and Relationship to Outplanting Performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; Ivetić, V. Root System Development and Field Establishment: Effect of Seedling Quality. New For. 2022, 53, 1021–1067. [Google Scholar] [CrossRef]
- South, D.B.; Starkey, T.E.; Lyons, A. Why Healthy Pine Seedlings Die after They Leave the Nursery. Forests 2023, 14, 645. [Google Scholar] [CrossRef]
- Solfjeld, I.; Hansen, O.B. Post-Transplant Growth of Five Deciduous Ordic Tree Species as Affected by Transplanting Date and Root Pruning. Urban For. Urban Green. 2004, 2, 129–137. [Google Scholar] [CrossRef]
- Andersen, L.; Rasmussen, H.N.; Brander, P.E. Regrowth and Dry Matter Allocation in Quercus robur (L.) Seedlings Root Pruned Prior to Transplanting. New For. 2000, 19, 205–214. [Google Scholar] [CrossRef]
- Łakomy, P.; Kuźmiński, R.; Mucha, J.; Zadworny, M. Effects of Oak Root Pruning in Forest Nurseries on Potential Pathogen Infections. For. Pathol. 2019, 49, e12513. [Google Scholar] [CrossRef]
- Vysotskaya, L.B.; Arkhipova, T.N.; Timergalina, L.N.; Dedov, A.V.; Veselov, S.Y.; Kudoyarova, G.R. Effect of Partial Root Excision on Transpiration, Root Hydraulic Conductance and Leaf Growth in Wheat Seedlings. Plant Physiol. Biochem. 2004, 42, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.R.; Morgenroth, J.; Koeser, A.K. The Effects of Root Pruning on Growth and Physiology of Two Acer Species in New Zealand. Urban For. Urban Green. 2019, 38, 64–73. [Google Scholar] [CrossRef]
- Bigras, F.J. Field Performance of Containerized Black Spruce Seedlings with Root Systems Damaged by Freezing or Pruning. New For. 1998, 15, 1–9. [Google Scholar] [CrossRef]
- Ramirez, J.A.; Vitali, V.; Martínez-Vilalta, J.; Handa, I.T.; Messier, C. Reserve Accumulation Is Prioritized Over Growth Following Single or Combined Injuries in Three Common North American Urban Tree Species. Front. Plant Sci. 2021, 12, 715399. [Google Scholar] [CrossRef]
- Hu, C.; Ding, M.; Qu, C.; Sadras, V.; Yang, X.; Zhang, S. Yield and Water Use Efficiency of Wheat in the Loess Plateau: Responses to Root Pruning and Defoliation. Field Crops Res. 2015, 179, 6–11. [Google Scholar] [CrossRef]
- Dong, T.; Duan, B.; Zhang, S.; Korpelainen, H.; Niinemets, Ü.; Li, C. Growth, Biomass Allocation and Photosynthetic Responses Are Related to Intensity of Root Severance and Soil Moisture Conditions in the Plantation Tree Cunninghamia lanceolata. Tree Physiol. 2016, 36, 807–817. [Google Scholar] [CrossRef]
- Chance, L.M.G.; Arnold, M.A.; Lombardini, L.; Watson, W.T.; Carver, S.T.; King, A.R. Landscape Establishment for Baldcypress, Red Maple, and Chaste Tree Is Delayed for Trees Transplanted from Larger Containers. J. Environ. Hortic. 2017, 35, 43–57. [Google Scholar] [CrossRef]
- Watson, G. Tree Size Affects Root Regeneration and Top Growth after Transplanting. J. Arboric. 1985, 11, 37–40. [Google Scholar] [CrossRef]
- Watson, W.T. Influence of Tree Size on Transplant Establishment and Growth. HortTechnology 2005, 15, 118–122. [Google Scholar] [CrossRef]
- Dostálek, J.; Weber, M.; Matula, S.; Frantík, T. Planting of Different-Sized Tree Transplants on Arable Soil. Open Life Sci. 2009, 4, 574–584. [Google Scholar] [CrossRef]
- Ivetic, V.; Grossnickle, S. Opening Letter. Reforesta 2016. [Google Scholar] [CrossRef]
- Klesse, S.; von Arx, G.; Gossner, M.M.; Hug, C.; Rigling, A.; Queloz, V. Amplifying Feedback Loop between Growth and Wood Anatomical Characteristics of Fraxinus excelsior Explains Size-Related Susceptibility to Ash Dieback. Tree Physiol. 2021, 41, 683–696. [Google Scholar] [CrossRef]
- Camarero, J.J. Within- versus between-Species Size Effects on Drought-Induced Dieback and Mortality. Tree Physiol. 2021, 41, 679–682. [Google Scholar] [CrossRef]
- DesRochers, A.; Tremblay, F. The Effect of Root and Shoot Pruning on Early Growth of Hybrid Poplars. For. Ecol. Manag. 2009, 258, 2062–2067. [Google Scholar] [CrossRef]
- Jing, D.-W.; Liu, F.-C.; Wang, M.-Y.; Ma, H.-L.; Du, Z.-Y.; Ma, B.-Y.; Dong, Y.-F. Effects of Root Pruning on the Physicochemical Properties and Microbial Activities of Poplar Rhizosphere Soil. PLoS ONE 2017, 12, e0187685. [Google Scholar] [CrossRef]
- Augspurger, C.K.; Bartlett, E.A. Differences in Leaf Phenology between Juvenile and Adult Trees in a Temperate Deciduous Forest. Tree Physiol. 2003, 23, 517–525. [Google Scholar] [CrossRef]
- Osada, N.; Hiura, T. Intraspecific Differences in Spring Leaf Phenology in Relation to Tree Size in Temperate Deciduous Trees. Tree Physiol. 2019, 39, 782–791. [Google Scholar] [CrossRef]
- Savage, J.A.; Chuine, I. Coordination of Spring Vascular and Organ Phenology in Deciduous Angiosperms Growing in Seasonally Cold Climates. New Phytol. 2021, 230, 1700–1715. [Google Scholar] [CrossRef] [PubMed]
- Goffinet, M.C.; Larson, P.R. Xylary union between the new shoot and old stem during terminal bud break in populus deltoides. Am. J. Bot. 1982, 69, 432–446. [Google Scholar] [CrossRef]
- Xie, Z.; Forney, C.F.; Bondada, B. Renewal of Vascular Connections between Grapevine Buds and Canes during Bud Break. Sci. Hortic. 2018, 233, 331–338. [Google Scholar] [CrossRef]
- Dong, S.; Cheng, L.; Scagel, C.F.; Fuchigami, L.H. Root Damage Affects Nitrogen Uptake and Growth of Young Fuji/M.26 Apple Trees. J. Hortic. Sci. Biotechnol. 2003, 78, 410–415. [Google Scholar] [CrossRef]
- Wilson, S.J.; Clark, R.J. Root Exposure Effects on Water Relations of Eucalyptus Nitens Nursery Stock. New For. 2000, 19, 13–25. [Google Scholar] [CrossRef]
- SEIWA, K. Changes in Leaf Phenology Are Dependent on Tree Height inAcer Mono, a Deciduous Broad-Leaved Tree. Ann. Bot. 1999, 83, 355–361. [Google Scholar] [CrossRef]
- Fu, Y.H.; Liu, Y.; De Boeck, H.J.; Menzel, A.; Nijs, I.; Peaucelle, M.; Peñuelas, J.; Piao, S.; Janssens, I.A. Three Times Greater Weight of Daytime than of Night-time Temperature on Leaf Unfolding Phenology in Temperate Trees. New Phytol. 2016, 212, 590–597. [Google Scholar] [CrossRef]
- Philipson, J.J. Root Growth in Sitka Spruce and Douglas-Fir Transplants: Dependence on the Shoot and Stored Carbohydrates. Tree Physiol. 1988, 4, 101–108. [Google Scholar] [CrossRef]
- Klimešová, J.; Herben, T.; Martínková, J. Disturbance Is an Important Factor in the Evolution and Distribution of Root-Sprouting Species. Evol. Ecol. 2017, 31, 387–399. [Google Scholar] [CrossRef]
- Jones, R.H.; Raynal, D.J. Root Sprouting in American Beech (Fagus Grandifolia): Effects of Root Injury, Root Exposure, and Season. For. Ecol. Manag. 1988, 25, 79–90. [Google Scholar] [CrossRef]
- Tromp, J. Nutrient Reserves in Roots of Fruit Trees, in Particular Carbohydrates and Nitrogen. Plant Soil 1983, 71, 401–413. [Google Scholar] [CrossRef]
- Stjernberg, E.I. Mechanical Shock during Transportation: Effects on Seedling Performance. New For. 1997, 13, 401–420. [Google Scholar] [CrossRef]
- Cox, R.M.; Zhu, X.B. Effects of Simulated Thaw on Xylem Cavitation, Residual Embolism, Spring Dieback and Shoot Growth in Yellow Birch. Tree Physiol. 2003, 23, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Colangelo, M.; Camarero, J.J.; Battipaglia, G.; Borghetti, M.; De Micco, V.; Gentilesca, T.; Ripullone, F. A Multi-Proxy Assessment of Dieback Causes in a Mediterranean Oak Species. Tree Physiol. 2017, 37, 617–631. [Google Scholar] [CrossRef]
- Lv, H.; Gangwisch, M.; Saha, S. Crown Die-Back of Peri-Urban Forests after Combined Heatwave and Drought Was Species-Specific, Size-Dependent, and Also Related to Tree Neighbourhood Characteristics. Sci. Total Environ. 2024, 913, 169716. [Google Scholar] [CrossRef]
- Crespo-Antia, J.P.; González de Andrés, E.; Gazol, A.; Camarero, J.J.; Linares, J.C. Tree-Level Climate Sensitivity Reveals Size Effects and Impending Growth Decline in Silver Fir Affected by Dieback. Forests 2024, 15, 999. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puértolas, J.; Dumroese, R.K.; Ivetić, V.; Molina-Venegas, R.; Arellano, E.C.; Li, G.; Ovalle, J.F. Climate and Species Stress Resistance Modulate the Higher Survival of Large Seedlings in Forest Restorations Worldwide. Ecol. Appl. 2021, 31, e02394. [Google Scholar] [CrossRef]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and Boreal Forest Tree Phenology: From Organ-Scale Processes to Terrestrial Ecosystem Models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef]
- Wang, H.; Gao, C.; Ge, Q. Low Temperature and Short Daylength Interact to Affect the Leaf Senescence of Two Temperate Tree Species. Tree Physiol. 2022, 42, 2252–2265. [Google Scholar] [CrossRef]
- Donnelly, A.; Yu, R.; Rehberg, C.; Schwartz, M.D. Variation in the Timing and Duration of Autumn Leaf Phenology among Temperate Deciduous Trees, Native Shrubs and Non-Native Shrubs. Int. J. Biometeorol. 2024, 68, 1663–1673. [Google Scholar] [CrossRef]
- Marchand, L.J.; Dox, I.; Gričar, J.; Prislan, P.; Leys, S.; Van den Bulcke, J.; Fonti, P.; Lange, H.; Matthysen, E.; Peñuelas, J.; et al. Inter-Individual Variability in Spring Phenology of Temperate Deciduous Trees Depends on Species, Tree Size and Previous Year Autumn Phenology. Agric. For. Meteorol. 2020, 290, 108031. [Google Scholar] [CrossRef] [PubMed]
- Lu, E.Y.; Tsai, C.H.; Lin, J.J.; Yang, S.H. Leaf Emergence, Shedding, and Lifespan of Dominant Hardwood Species in Chitou, Central Taiwan. Bot. Stud. 2012, 53, 255–264. [Google Scholar]
- Zani, D.; Crowther, T.W.; Mo, L.; Renner, S.S.; Zohner, C.M. Increased Growing-Season Productivity Drives Earlier Autumn Leaf Senescence in Temperate Trees. Science 2020, 370, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Sha, Y.; Sun, L.; Li, Z. Abiotic Stress-Induced Leaf Senescence: Regulatory Mechanisms and Application. Int. J. Mol. Sci. 2023, 24, 11996. [Google Scholar] [CrossRef]
- Hesse, B.D.; Goisser, M.; Hartmann, H.; Grams, T.E.E. Repeated Summer Drought Delays Sugar Export from the Leaf and Impairs Phloem Transport in Mature Beech. Tree Physiol. 2019, 39, 192–200. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Turcsán, A.; Maes, J.; Duchêne, N.; Meeus, S.; Steppe, K.; Steenackers, M. Repeated Summer Drought and Re-Watering during the First Growing Year of Oak (Quercus petraea) Delay Autumn Senescence and Bud Burst in the Following Spring. Front. Plant Sci. 2016, 7, 419. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, L.; Qi, D. Variation in Leaf Traits at Different Altitudes Reflects the Adaptive Strategy of Plants to Environmental Changes. Ecol. Evol. 2020, 10, 8166–8175. [Google Scholar] [CrossRef]
- Baird, A.S.; Anderegg, L.D.L.; Lacey, M.E.; HilleRisLambers, J.; Van Volkenburgh, E. Comparative Leaf Growth Strategies in Response to Low-Water and Low-Light Availability: Variation in Leaf Physiology Underlies Variation in Leaf Mass per Area in Populus Tremuloides. Tree Physiol. 2017, 37, 1140–1150. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Vitali, V.; Ramirez, J.A.; Perrette, G.; Delagrange, S.; Paquette, A.; Messier, C. Complex Above- and Below-Ground Growth Responses of Two Urban Tree Species Following Root, Stem, and Foliage Damage-An Experimental Approach. Front. Plant Sci. 2019, 10, 1100. [Google Scholar] [CrossRef]
- Jones, D.A.; Harrington, C.A.; Marshall, D. Survival, and Growth Response of Douglas-Fir Trees to Increasing Levels of Bole, Root, and Crown Damage. For. Sci. 2019, 65, 143–155. [Google Scholar] [CrossRef]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water Deficits Uncouple Growth from Photosynthesis, Increase C Content, and Modify the Relationships between C and Growth in Sink Organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.A.; Handa, I.T.; Posada, J.M.; Delagrange, S.; Messier, C. Carbohydrate Dynamics in Roots, Stems, and Branches after Maintenance Pruning in Two Common Urban Tree Species of North America. Urban For. Urban Green. 2018, 30, 24–31. [Google Scholar] [CrossRef]
- Mestre, L.M.; Argañaraz, C.I.; Preiß-Daimler, I.; Fernandez, L.; Turi, L.; Soler, R. Influence of Environmental Conditions and Initial Sapling Size in Nothofagus Survival and Growth: Implications for Restoration of Burnt sub-Antarctic Forests. Austral Ecol. 2024, 49, e13269. [Google Scholar] [CrossRef]
- Liu, M.; Zhu, Y.; Pang, R.; Gao, L. Can Growth Increase of Small Trees after Drought Compensate for Large Trees’ Growth Loss? Forests 2024, 15, 448. [Google Scholar] [CrossRef]
- Ivetić, V.; Devetaković, J.; Maksimović, Z. Initial Height and Diameter Are Equally Related to Survival and Growth of Hardwood Seedlings in First Year after Field Planting. Reforesta 2016, 6–21. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size/Index | H (cm) | DBH (mm) | RCD (mm) | H/DBH | H/RCD |
|---|---|---|---|---|---|
| Large | 308.75 ± 9.66a | 15.94 ± 0.68a | 28.47 ± 0.61a | 17.64 ± 5.73b | 9.85 ± 3.22a |
| Medium | 238.00 ± 7.71b | 11.07 ± 0.49b | 24.20 ± 0.43b | 19.51 ± 6.35a | 8.94 ± 2.91b |
| Small | 138.92 ± 7.18c | 6.59 ± 0.51c | 14.87 ± 0.29c | 19.20 ± 6.14a | 8.52 ± 2.73c |
| Treatment | CK | P1 | P2 | P3 |
|---|---|---|---|---|
| Large | 0.00% ± 0.00%cB | 3.33% ± 2.89%bA | 31.67% ± 5.77%aA | 33.33% ± 2.89%aA |
| Middle | 0.00% ± 0.00%bB | 1.67% ± 1.44%bA | 15.00% ± 5.00%aB | 20.00% ± 5.00%aB |
| Small | 3.33% ± 2.89%bA | 1.67% ± 1.44%bA | 6.67% ± 2.89%aC | 15.00% ± 5.00%aB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Zhang, J.; Wan, F.; Xian, L.; Liu, Y. Effects of Root Pruning and Size on Growth Traits of Hybrid Poplar Seedlings. Forests 2024, 15, 1770. https://doi.org/10.3390/f15101770
Chang X, Zhang J, Wan F, Xian L, Liu Y. Effects of Root Pruning and Size on Growth Traits of Hybrid Poplar Seedlings. Forests. 2024; 15(10):1770. https://doi.org/10.3390/f15101770
Chicago/Turabian StyleChang, Xiaochao, Jin Zhang, Fangfang Wan, Lihong Xian, and Yong Liu. 2024. "Effects of Root Pruning and Size on Growth Traits of Hybrid Poplar Seedlings" Forests 15, no. 10: 1770. https://doi.org/10.3390/f15101770