
Evaluation of Fine Root Morphology and Rhizosphere Environmental Characteristics of the Dioecious Idesia polycarpa Maxim
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Test Area
2.2. Experimental Materials
2.3. Fine Root Morphology Measurement
2.4. Soil Nutrient Determination and Analysis
2.5. Genome Sequencing
2.6. Bioinformatics Analysis
2.7. Determination of Rhizosphere Soil Metabolites
2.8. Metabolite Data Analysis
3. Results
3.1. Fine Root Characteristics of Female and Male Idesia polycarpa
3.2. Nutrient Characteristics of Rhizosphere Soil of Male and Female Plants
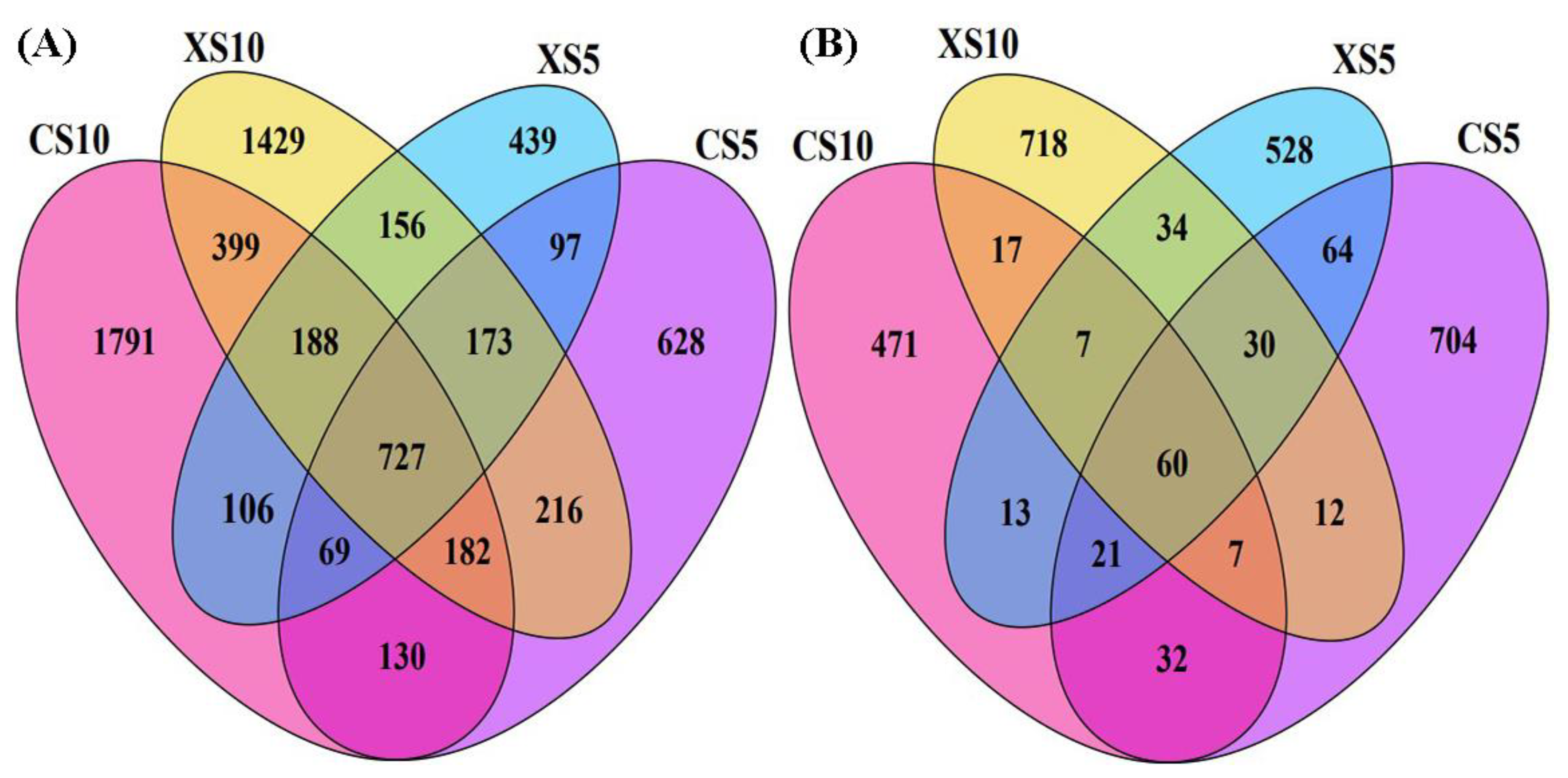
3.3. Microbial Diversity in Rhizosphere Soil of I. polycarpa in Different Stages
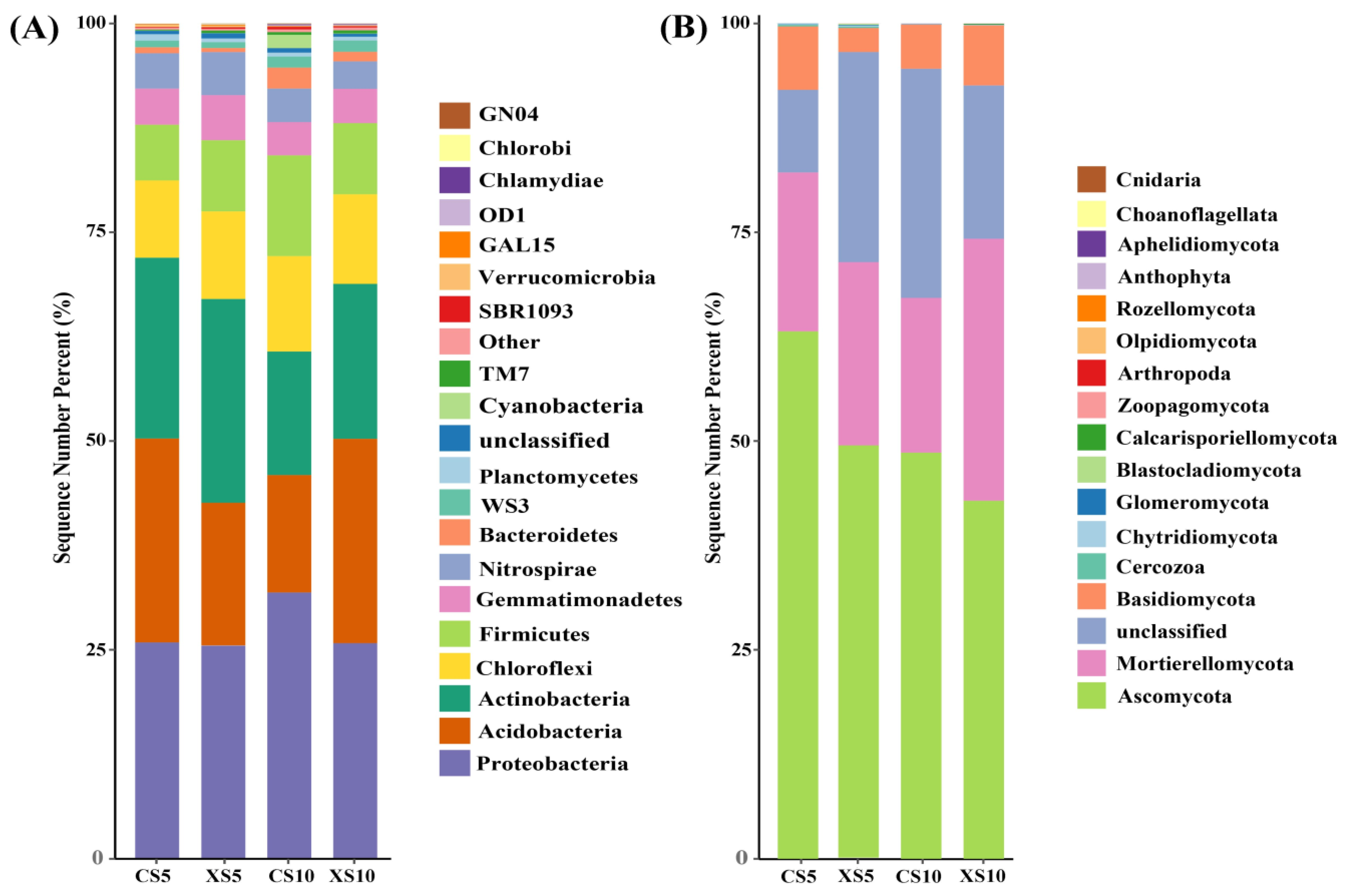
3.4. Microbial Community Composition in Rhizosphere Soil of Male and Female Plants
3.5. Principal Component Analysis of Metabolites in Rhizosphere Soil of Male and Female Plants
3.6. Screening of Differential Metabolites in Rhizosphere Soil of I. polycarpa
3.7. KEGG Enrichment Analysis
3.8. Effects of Rhizosphere Environment on Root Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, Y.B.; Jiang, Y.L.; Chen, K.; Duan, B.L.; Zhang, S.; Korpelaiinen, H.; Nijinemets, Ü.; Li, C.Y. Reproductive investments driven by sex and altitude in sympatric Populus and Salix trees. Tree Physiol. 2017, 37, 1503–1514. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Guo, D.L. Root Ecology. Chin. J. Plant Ecol. 2008, 32, 1213–1216. [Google Scholar] [CrossRef]
- Vega-Frutis, R.; Guevara, R. Different arbuscular mycorrhizal interactions in male and female plants of wild Carica papaya L. Plant Soil 2009, 322, 165–176. [Google Scholar] [CrossRef]
- Chen, J.; Duan, B.; Wang, M.; Korpelainen, H.; Li, C. Intra- and inter-sexual competition of Populus cathayana under different watering regimes. Funct. Ecol. 2014, 28, 124–136. [Google Scholar] [CrossRef]
- Wen, X.M.; Xian, T.; Liu, J.Y. Effects of defoliation on growth and non-structural carbohydrates in female and male Populus cathayana chin. J. Ecol. 2021, 40, 41–48. [Google Scholar] [CrossRef]
- Zhang, L.M.; Chen, J.; Shen, W.T.; Duan, X.L.; Ge, Y.L.; Dong, Y.F.; Xu, X. Different morphological and physiological responses of Populus cathayana to drought under male and female combination mode. J. China West Norm. Univ. 2019, 40, 325–331. [Google Scholar] [CrossRef]
- Chen, G.T.; Tu, L.H.; Peng, Y.; Hu, H.L.; Hu, T.X.; Xu, Z.F.; Liu, L.; Tang, Y. Effect of nitrogen additions on root morphology and chemistry in a subtropical bamboo forest. Plant Soil 2017, 412, 441–451. [Google Scholar] [CrossRef]
- Zhang, J.R.; Yan, X.J.; Jia, L.Q.; Fan, A.L.; Wang, X.; Chen, T.T.; Chen, G.S. Morphology and C and N stoichiometry traits of fine Roots of nine understory shrubs in subtropical natural evergreen broad-leaved forest. Acta Ecol. Sin. 2022, 42, 3716–3726. [Google Scholar] [CrossRef]
- Fan, B.; Ma, Z.; Gao, P.; Lu, J.; Ding, N.; Sun, K. Functional traits of male and female leaves of Hippophae tibetana on the eastern edge of the Tibetan Plateau and their altitudinal variability. Plants 2022, 11, 2484. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Z. Effects of Competition Modes on Growth and Physiological Characteristics of Populus yunnanensis in Different Light Environments; Sichuan Agricultural University: Sichuan, China, 2018. [Google Scholar] [CrossRef]
- Song, H.F.; Cai, Z.Y.; Liao, J.; Tang, D.T.; Zhang, S. Sexually differential gene expressions in Poplar roots in response to nitrogen deficiency. Tree Physiol. 2019, 39, 1614–1629. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Ren, Y.; Chen, Z.X.; Shi, Y.L.; Chen, C.; Sun, G.Q.; Zhou, Y.Q.; Li, Q. Nutrient differences between male and female Zanthoxylum shinifolium sieb. et zucc. and their effects on soil nutrient. Non Wood For. Res. 2022, 40, 192–199. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, X.X.; Zhang, D.; Dong, Z.; Ji, H.; Li, H. Localized nutrient supply promotes Maize growth and nutrient acquisition by shaping root morphology and physiology and mycorrhizal symbiosis. Soil Tillage Res. 2023, 25, 105550. [Google Scholar] [CrossRef]
- Yang, F.; Jin, C.; Wang, S.; Wang, Y.J.; Wei, L.; Zheng, L.H.; Gu, H.P.; Lam, S.S.; Naushad, M.; Li, C.; et al. Bamboo-based magnetic activated carbon for efficient removal of sulfadiazine: Application and adsorption mechanism. Chemosphere 2023, 323, 138245. [Google Scholar] [CrossRef]
- Yu, L.; Huang, Z.; Li, Z.; Korpelainen, H.; Li, C. Sex-specific strategies of nutrient resorption associated with leaf economics in Populus euphratica. J. Ecol. 2022, 110, 2062–2073. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, H.; Fu, S.; Yao, Q. Variation in soil microbial community structure associated with different Legume species is greater than that associated with different Grass species. Front. Microbiol. 2017, 8, 1007. [Google Scholar] [CrossRef]
- Xiao, M.; Zang, H.D.; Liu, S.L.; Ye, R.Z.; Zhu, Z.K.; Su, Y.R.; Wu, J.S.; Ge, T.D. Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice–soil systems: A 13C-CO2 pulse labeling study. Plant Soil 2019, 445, 101–112. [Google Scholar] [CrossRef]
- Guo, Q.X.; Yan, L.J.; Korpelainen, H.; Niinemets, Ü.; Li, C.Y. Plant-plant interactions and N fertilization shape soil bacterial and fungal communities. Soil Biol. Biochem. 2019, 128, 127–138. [Google Scholar] [CrossRef]
- Gu, Y.J.; Liu, Y.H.; Yu, X.W.; Feng, Y.; Yang, X.Y.; Lam, S.S.; Naushad, M.; Li, C.; Sonne, C. Autochthonous bioaugmentation accelerates phenanthrene degradation in acclimated soil. Environ. Res. 2023, 224, 115543. [Google Scholar] [CrossRef] [PubMed]
- Du, S.N.; Pan, H.Y.; Xing, Q.N.; Zhang, A.L.; Wang, Q.H. Effects of nitrogen application on amino acid content in rhizosphere of Beet at different development stages. Chin. Agric. Sci. Bull. 2022, 38, 81–88. [Google Scholar] [CrossRef]
- Miller, S.B.; Heuberger, A.L.; Broeckling, C.D.; Jahn, C.E. Non-targeted metabolomics reveals Sorghum rhizosphere-associated exudates are influenced by the belowground interaction of substrate and Sorghum genotype. Int. J. Mol. Sci. 2019, 20, 431. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; He, Y.; Yu, L.; Li, Z.; Korpelainen, H.; Li, C. Revealing interactions between root phenolic metabolomes and rhizosphere bacterial communities in Populus euphratica plantations. Biol. Fertil. Soils 2021, 57, 421–434. [Google Scholar] [CrossRef]
- Xue, X.Y.; Liu, Z.; Wang, Y.M.; Geng, X.D.; Cai, Q.F.; Li, Z. Analysis on the development advantages and countermeasures of I. polycarpa industry in Henan province. Hubei Agric. Sci. 2022, 61, 115–120. [Google Scholar] [CrossRef]
- Jiang, H.Y.; Pang, G.Y.; Kang, T.R. Purification technology and antioxidant activity of I. polycarpa oil in vitro. Cereals Oils 2022, 35, 59–64. [Google Scholar]
- Xu, J.J.; Chen, J.T.; Xie, T.; Li, F.; Geng, Y.Y.; Zhu, Y.Y.; Xu, J. Analysis and comprehensive evaluation of fruit phenotypic diversity in natural populations of I. polycarpa. Seed 2021, 40, 64–71. [Google Scholar] [CrossRef]
- Fang, L.S.; Xu, Z.H.; Liu, Z.; Li, Z.; Geng, X.D.; Cai, Q.F.; Zhai, W.J.; Zhou, H.Q.; Wang, Y.M. Changes of contents, endogenous hormones and photosynthetic characteristics of I. polycarpa fruit at the different developmental stages. Sci. Silvae Sin. 2020, 56, 41–52. [Google Scholar] [CrossRef]
- Zhang, X.X.; Liu, Z.; Wang, X.J.; Li, Z.; Cai, Q.F.; Geng, X.D.; Wang, Y.M.; Wu, H.Y. Comparison on the growth performance of 2-Year Old I. polycarpa seedlings from different provenances. J. Northwest For. Univ. 2021, 36, 118–123. [Google Scholar] [CrossRef]
- Rana, S. Study on the Influence Mechanism of Water and Fertilizer Coupling on the Growth and Development of Idesia polycarpa Seedlings. Ph.D. Thesis, College of Forestry, Henan Agricultural University, Henan, China, 2024. [Google Scholar]
- Wu, H.B.; OuYang, S.X.; Liang, J. Research on soil quality characteristics of green areas in Zhengzhou International Tourism Resort Area. Landsc. Gard. 2021, 38, 18–23. [Google Scholar]
- Johnson, M.G.; Tingey, D.T.; Philips, D.L.; Storm, M.J. Advancing fine root research with minirhizotrons. Environ. Exp. Bot. 2001, 45, 263–289. [Google Scholar] [CrossRef]
- Chen, X.Y.; Eamus, D.; Hutley, L.B. Seasonal patterns of fine-root productivity and turnover in a tropical savanna of northern Australia. J. Trop. Ecol. 2004, 20, 221–224. [Google Scholar] [CrossRef]
- Shibata, H.; Hiura, T.; Tanaka, Y.; Takagi, K.; Koike, T. Carbon cycling and budget in a forested basin of southwestern Hokkaido, northern Japan. Ecol. Res. 2005, 20, 325–331. [Google Scholar] [CrossRef]
- Zhao, L.; Shang, S.; Shi, D.L.; Xu, H.; Wang, J. Composition and structural characteristics of rhizosphere microorganisms of Polygonum sibiricum (Laxm.) Tzvelev in the Yellow River Delta. Diversity 2022, 14, 965. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 2000; pp. 263–270. ISBN 7-109-06644-4. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Roybal, C.M.; Butterfield, B.J. Species-specific trait-environment relationships among populations of widespread grass species. Oecologia 2019, 189, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, Y.F.; Liu, X.Y.; Yao, Y.F.; Qi, W. Stump height after regenerative cutting of sea-buckthorn (Hippophae rhamnoides) affects fine root architecture and rhizosphere soil stoichiometric properties. Rhizosphere 2022, 24, 100602. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.L.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Shively, T.J.; Cook, R.; Maier, C.A.; Garcia, K.; Albaugh, T.J.; Campoe, O.; Leggett, Z. Readily available resources across sites and genotypes result in greater aboveground growth and reduced fine root production in Pinus taeda. For. Ecol. Manag. 2022, 521, 120431. [Google Scholar] [CrossRef]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Teitel, Z.; Pickup, M.; Field, D.L.; Barrett, S.C. The dynamics of resource allocation and costs of reproduction in a sexually dimorphic, wind-pollinated dioecious plant. Plant Biol. 2016, 18, 98–103. [Google Scholar] [CrossRef]
- Rana, S.; Jemim, R.S.; Li, Z.; Geng, X.D.; Wang, Y.M.; Cai, Q.F.; Liu, Z. Study on the pattern of reproductive allocation and fruit development in young dioecious trees of I. polycarpa. S. Afr. J. Bot. 2022, 146, 472–480. [Google Scholar] [CrossRef]
- Hoffmann, A.J.; Alliende, M.C. Interactions in the patterns of vegetative growth and reproduction in woody dioecious plants. Oecologia 1984, 61, 109–114. [Google Scholar] [CrossRef]
- Zhao, S.C.; Yan, T.F.; Wang, F.H. Root growth and development law and its correlation with aboveground growth of Xinyang Peach in May. Jiangsu Agric. Sci. 2016, 44, 212–213. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Zhang, P.F.; Trivedi, P.; Riera, N.; Wang, Y.Y.; Liu, X.; Fan, G.Y.; Tang, J.L.; Coletta-Filho, H.D.; et al. The structure and function of the global Citrus rhizosphere microbiome. Nat. Commun. 2018, 9, 4894. [Google Scholar] [CrossRef] [PubMed]
- Samad, A.; Trognitz, F.; Compant, S.; Antonielli, L.; Sessitsch, A. Shared and host-specific microbiome diversity and functioning of grapevine and accompanying weed plants: Microbial communities associated with grapevine and vineyard weeds. Environ. Microbiol. 2017, 19, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Liu, J.; Yu, L.; Korpelainen, H.; Li, C.Y. Different sexual impacts of dioecious Populus euphratica on microbial communities and nitrogen cycle processes in natural forests. For. Ecol. Manag. 2021, 496, 119403. [Google Scholar] [CrossRef]
- Wu, G.; Yu, F.; Yuan, M.; Wang, J.B.; Liu, C.; He, W.Z.; Ge, Z.H.; Sun, Y.X.; Liu, Y. Responses of rhizosphere bacterial and fungal communities to the long-term continuous monoculture of Water oat. Microorganisms 2022, 10, 2174. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Qin, L.Y.; Sun, X.Y.; Zhao, S.W.; Yu, L.; Chen, S.B.; Wang, M. Salt stress-induced changes in soil metabolites promote cadmium transport into Triticum aestivum tissues. Environ. Sci. 2023, 127, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 2002, 245, 35–47. [Google Scholar] [CrossRef]
- Li, C.W.; Bai, X.F.; Chen, G.Z.; Zhu, P.; Zhang, S.T.; Hou, Y.P.; Zhang, X.X. Differences in soil nutrients and phenolic acid Metabolites contents in American ginseng cultivated soils with different restoration years. Chin. Plant Ecol. 2021, 45, 1263–1274. [Google Scholar] [CrossRef]
- Salcedo-Vite, K.; Sigala, J.-C.; Segura, D.; Gosset, G.; Martinez, A. Acinetobacter baylyi ADP1 growth performance and lipid accumulation on different carbon sources. Appl. Microbiol. Biotechnol. 2019, 103, 6217–6229. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhang, S.M.; Yuan, J.C.; Cheng, S.; Liang, Y.; Liu, S.T.; Ren, J.; Cai, H.G.; Liu, J.Z. Accumulation and transport characteristics of nutrient for spring maize in different cultivation modes. Soil Fert. Sci. China 2022, 153–161. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Du, S.F.; Wang, G.Y.; Zhang, J.C.; Jiang, J. Effects of exogenous (K+) potassium application on plant hormones in the roots of Tamarix ramosissima under NaCl stress. Genes 2022, 13, 1803. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X. Effect of K nutrition on Oryza sativa root growth and nutrient uptake. Acta Pedol. Sin. 1997, 182–188. [Google Scholar] [CrossRef]
- Orcutt, B.N.; Sylvan, J.B.; Knab, N.J.; Edwards, K.J. Microbial ecology of the dark ocean above, at, and below the seafloor. Microbiol. Mol. Biol. Rev. 2011, 75, 361–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.F.; Vivanco, J.M.; Shen, Q.R. The unseen rhizosphere root-soil-microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef]
- Mendes, L.W.; Tsai, S.M.; Navarrete, A.A.; De, H.M.; Van, V.J.A.; Kuramae, E.E. Soil-borne microbiome: Linking diversity to function. Microb. Ecol. 2015, 70, 255–265. [Google Scholar] [CrossRef]
- Wen, T.; Xie, P.; Penton, C.R.; Hale, L.; Thomashow, L.S.; Yang, S.D.; Ding, Z.X.; Su, Y.Q.; Yuan, J.; Shen, Q.R. Specific metabolites drive the deterministic assembly of diseased rhizosphere microbiome through weakening microbial degradation of autotoxin. Microbiome 2022, 10, 177. [Google Scholar] [CrossRef]
- Li, L.; Jiang, J.L.; Dong, Y.X.; Shu, Y. Allelochemical substances screening and their allelopathic effects analysis in rhizosphere soil of continuous cropping Panacis quinquefolii radix. Acta Agric. Boreali-Occident. Sin. 2022, 31, 1046–1057. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Goods Coverage | Shannon | Simpson | ACE | Chao1 |
|---|---|---|---|---|---|
| CS5 | 1 ± 0 a | 6.1502 ± 0.3259 a | 0.9633 ± 0.0066 a | 368.3333 ± 66.3961 a | 368.3333 ± 66.3961 a |
| XS5 | 1 ± 0 a | 5.9245 ± 0.4606 a | 0.9554 ± 0.0113 a | 233.3333 ± 44.7002 a | 233.3333 ± 44.7002 a |
| CS10 | 1 ± 0 a | 5.9357 ± 0.085 a | 0.9588 ± 0.0039 a | 304.6666 ± 32.6615 a | 304.6666 ± 32.6615 a |
| XS10 | 0.9999 ± 0.0001 a | 5.6706 ± 0.2073 a | 0.9439 ± 0.0162 a | 323.5778 ± 20.4549 a | 323.3476 ± 20.2630 a |
| Sample | Goods Coverage | Shannon | Simpson | ACE | Chao1 |
|---|---|---|---|---|---|
| CS5 | 0.9985 ± 0.0001 b | 9.5746 ± 0.0536 b | 0.9980 ± 0.0001 a | 1123.1685 ± 47.0738 c | 1129.1540 ± 48.0329 c |
| XS5 | 0.9995 ± 0 a | 9.8344 ± 0.1153 a | 0.9975 ± 0.0005 a | 1631.3415 ± 28.9380 b | 1633.2107 ± 29.9486 b |
| CS10 | 0.9988 ± 0.0003 b | 9.4560 ± 0.0547 b | 0.9978 ± 0.0002 a | 1039.9429 ± 18.5783 c | 1050.0278 ± 18.1969 c |
| XS10 | 0.9995 ± 0 a | 10.0815 ± 0.0684 a | 0.9982 ± 0.0002 a | 1805.6163 ± 8.4434 a | 1807.9299 ± 9.0967 a |
| Comparison Group | Total Number of Differential Metabolites | The Top Five Metabolites of VIP Value | Log2FC | p-Value | VIP | Metabolite Types | Type |
|---|---|---|---|---|---|---|---|
| XS5 VS CS5 | 19 | Myo-Inositol 3 | 3.02 × 100 | 0.014613 | 1.561137 | Alcohol | down |
| D-(-)-Ribofuranose 2 | 1.57 × 100 | 0.009517 | 1.559431 | Carbohydrate | down | ||
| D-Allose 2 | 1.92 × 100 | 0.013674 | 1.549882 | Carbohydrate | down | ||
| D(+)-Talose | 2.02 × 100 | 0.017956 | 1.54911 | Carbohydrate | down | ||
| Sucrose | 1.62 × 100 | 0.000101 | 1.547348 | Carbohydrate | down | ||
| XS10 VS CS10 | 10 | Glucose 1 | 1.469326 | 3.42 × 10−6 | 1.886262 | Carbohydrate | up |
| 4-(2-Methylbutanoyl)sucrose | 0.796648 | 0.007635 | 1.821765 | Carbohydrate | up | ||
| 2-Phenyl-1,3-oxazol-2-ine | 0.569156 | 0.021172 | 1.737153 | Carbohydrate | up | ||
| N-ethyl-Acetamide | −0.10944 | 0.01822 | 1.736087 | Carbohydrate | down | ||
| D-Arabinitol 2 | 0.483392 | 0.00822 | 1.73337 | Carbohydrate | up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Yang, Y.; Feng, J.; Rana, S.; Wang, S.; Wang, H.; Zhang, T.; Wang, Y.; Guo, G.; Cai, Q.; et al. Evaluation of Fine Root Morphology and Rhizosphere Environmental Characteristics of the Dioecious Idesia polycarpa Maxim. Forests 2024, 15, 234. https://doi.org/10.3390/f15020234
Li Z, Yang Y, Feng J, Rana S, Wang S, Wang H, Zhang T, Wang Y, Guo G, Cai Q, et al. Evaluation of Fine Root Morphology and Rhizosphere Environmental Characteristics of the Dioecious Idesia polycarpa Maxim. Forests. 2024; 15(2):234. https://doi.org/10.3390/f15020234
Chicago/Turabian StyleLi, Zhi, Yi Yang, Jian Feng, Sohel Rana, Shasha Wang, Huimin Wang, Tao Zhang, Yanmei Wang, Gaiping Guo, Qifei Cai, and et al. 2024. "Evaluation of Fine Root Morphology and Rhizosphere Environmental Characteristics of the Dioecious Idesia polycarpa Maxim" Forests 15, no. 2: 234. https://doi.org/10.3390/f15020234