Adaptation of the Coniferous Forests to Natural Fire Disturbances in the Altai Mountains, Xinjiang, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Fire Disturbance Year Determination
2.3. Relative Air Humidity Monitoring in Forested Areas
2.4. Bark Properties
2.5. Measurement of Flammability
- (1)
- Determination of ash content
- (2)
- Determination of crude fat content
- (3)
- HHV determination
2.6. Significant Value Calculations
2.7. Redundancy Analysis
3. Results
3.1. Natural Fire Disturbance and Forest Species Composition in Altai Forests
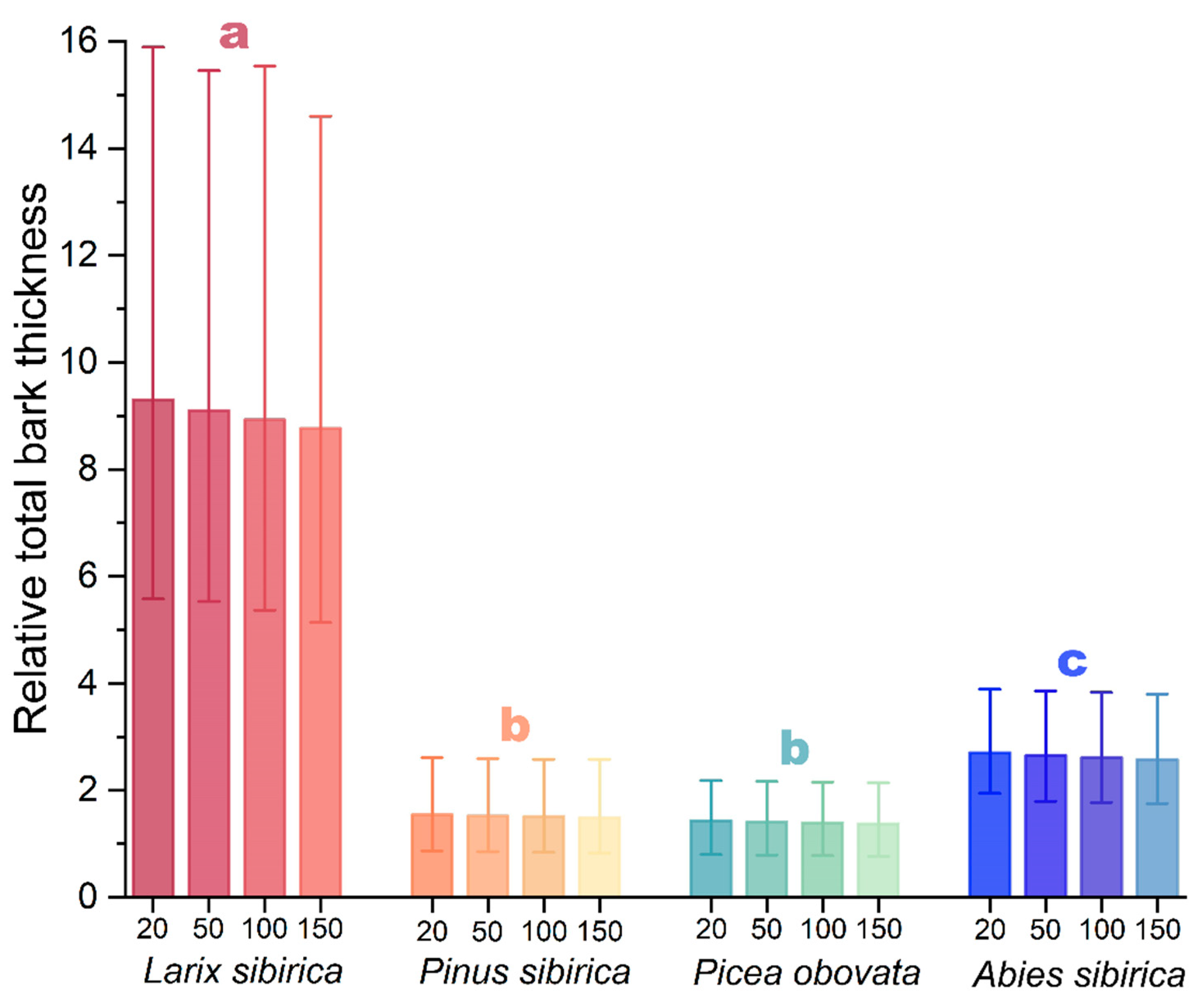
3.2. Bark Thickness Fire Adaptation
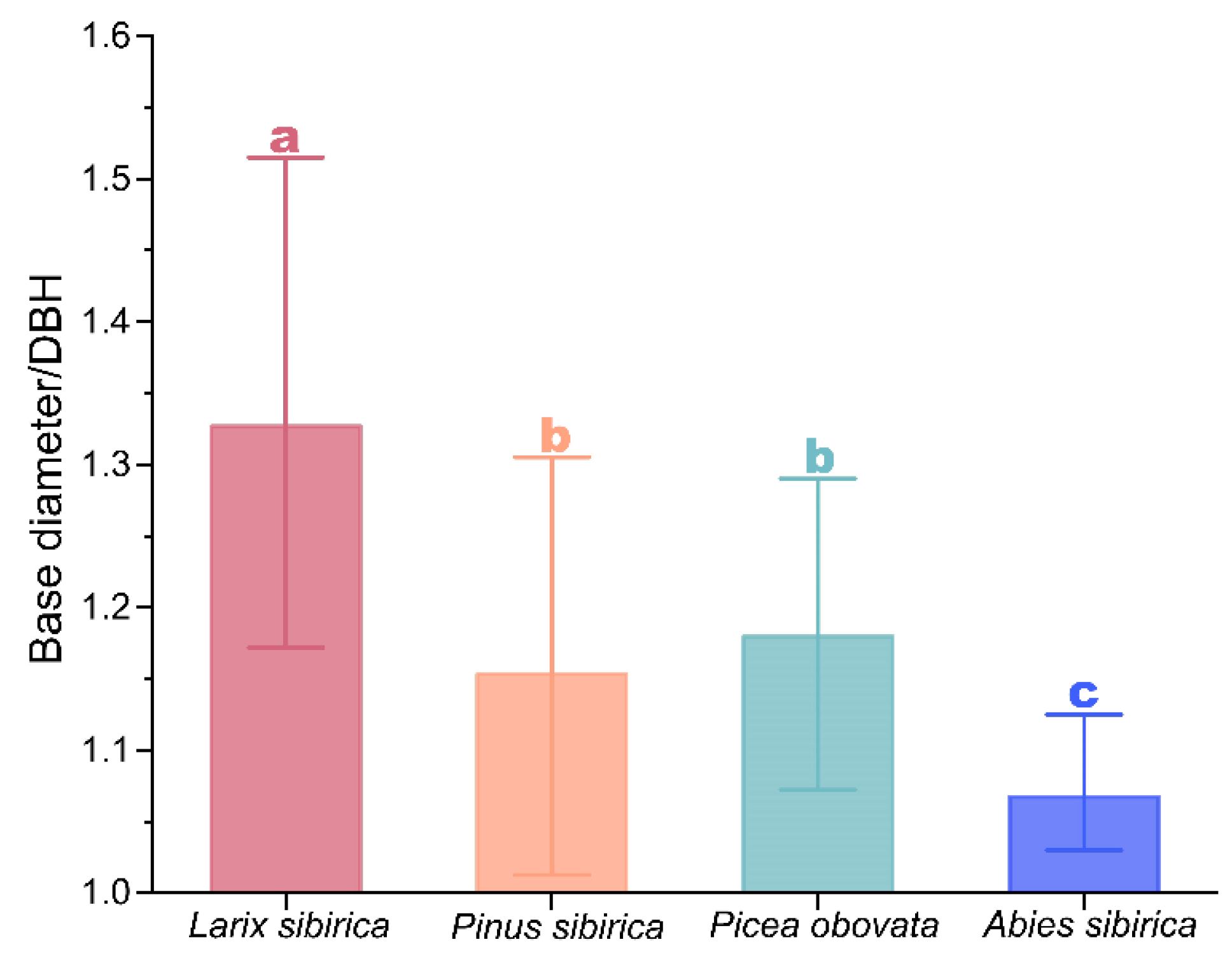
3.3. Differences in Basal Diameter/DBH
3.4. Differences in Larix sibirica Bark in Different Regions
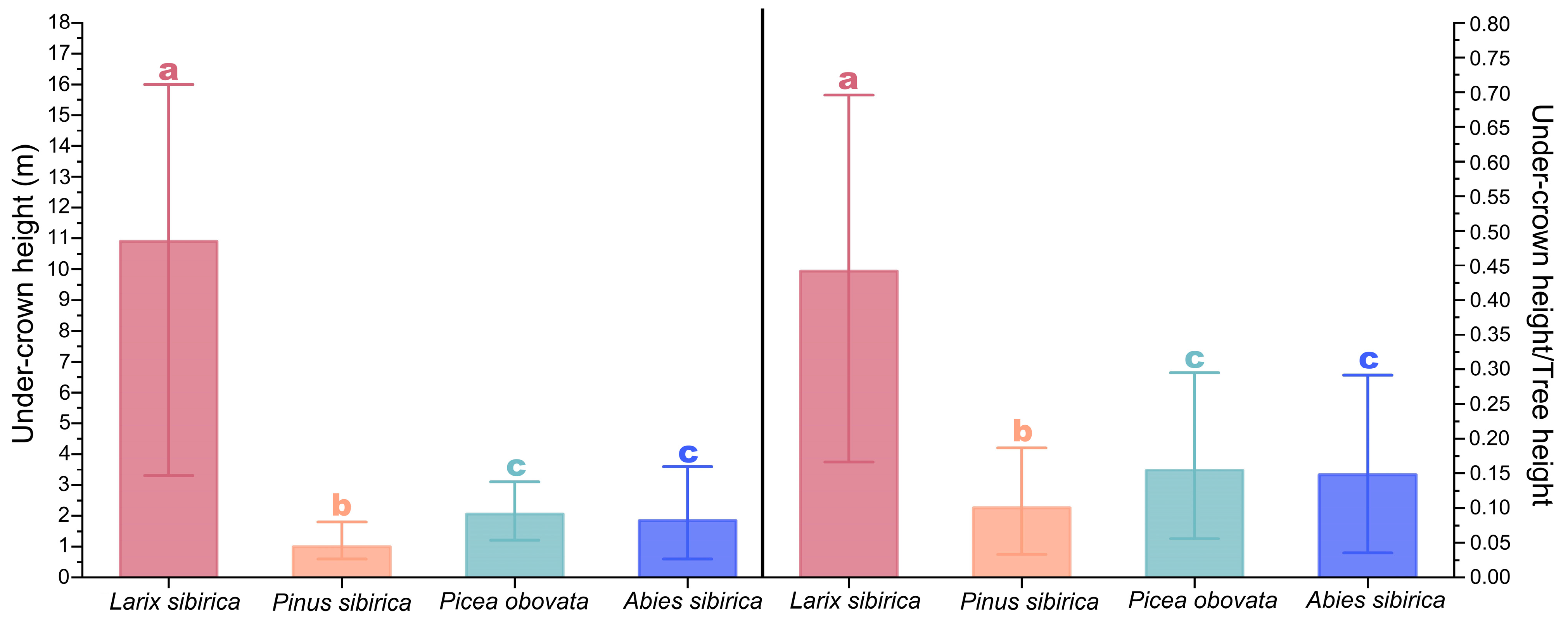
3.5. Differences in Under-Crown Height
3.6. Comparison of Flammability
3.7. Seed Characteristics of Different Conifers
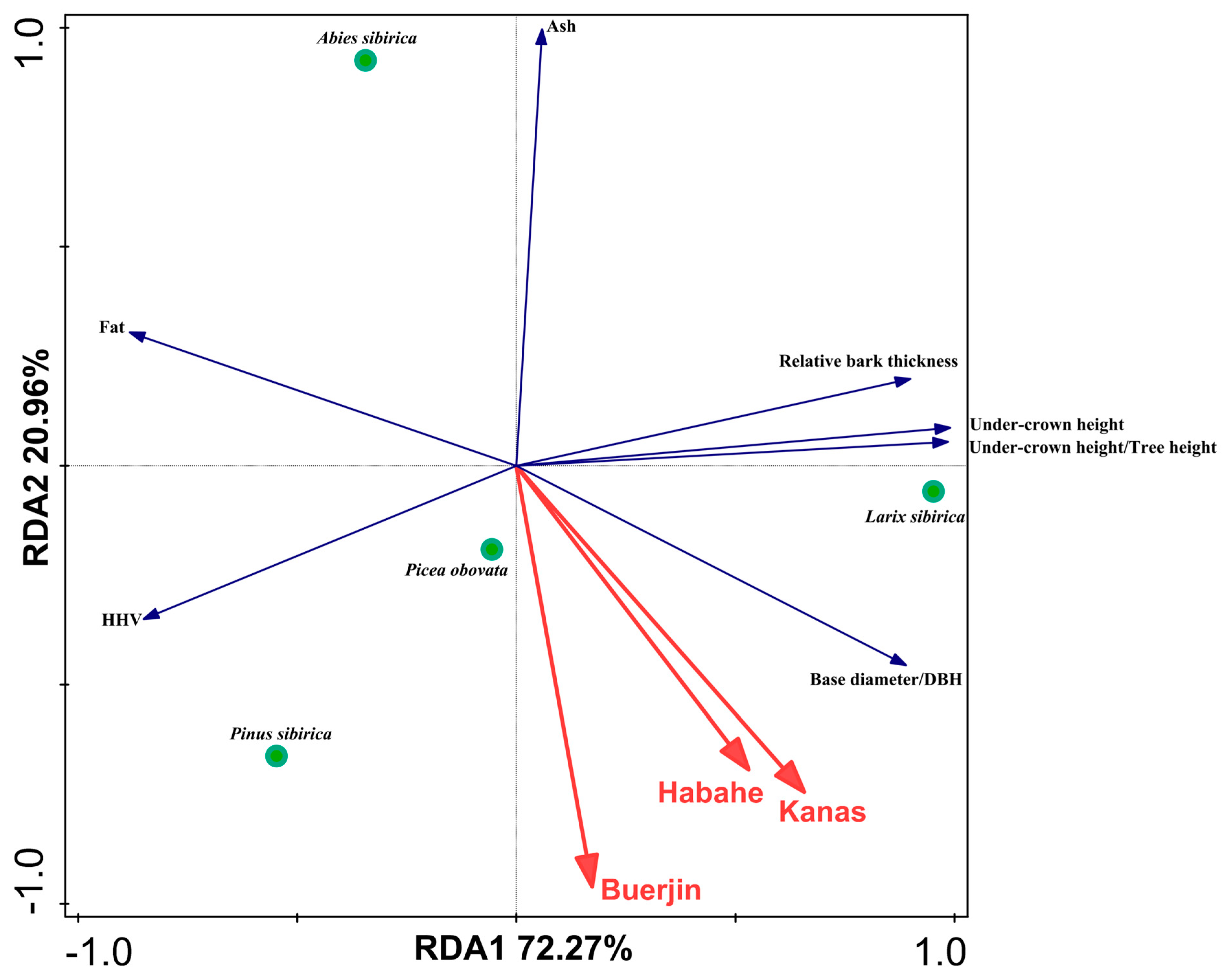
3.8. Association between the Fire Adaptation Traits of Coniferous Trees and the Distribution of Vegetation in Different Forest Areas
3.9. Association between Natural Fire Disturbance Scenarios and the Distribution of Different Conifer Species
4. Discussion
4.1. Characterization of Fire Adaptation of Four Conifer Species within the Altai Forests
4.2. Stress Effects of Larix sibirica Bark from Fire Disturbances
4.3. Ecological Significance of Natural Fire Disturbances in a Coniferous Forest in the Altai Mountains
4.4. Discussion on Forest Fire Management Measures in the Altai Forest Area
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glasspool, I.J.; Edwards, D.; Axe, L. Charcoal in the Silurian as evidence for the earliest wildfire. Geology 2004, 32, 381–384. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the earth system. Science 2009, 324, 481. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. A burning story: The role of fire in the history of life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef]
- Pausas, J.G. Evolutionary fire ecology: Lessons learned from pines. Trends Plant Sci. 2015, 20, 318–324. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Belcher, C.M.; Lamont, B.B.; Lim, S.L. A 350-million-year legacy of fire adaptation among conifers. J. Ecol. 2016, 104, 352–363. [Google Scholar] [CrossRef]
- Crisp, M.D.; Burrows, G.E.; Cook, L.G.; Thornhill, A.H.; Bowman, D.M.J.S. Flammable biomes dominated by eucalypts originated at the Cretaceous–Palaeogene boundary. Nat. Commun. 2011, 2, 193. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lamont, B.B.; Downes, K.S. Banksia born to burn. New Phytol. 2011, 191, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Lamont, B.B.; He, T.; Downes, K.S. Adaptive responses to directional trait selection in the Miocene enabled Cape proteas to colonize the savanna grasslands. Evol. Ecol. 2013, 27, 1099–1115. [Google Scholar] [CrossRef]
- Santini, F.; Climent, J.M.; Voltas, J. Phenotypic integration and life history strategies among populations of Pinus halepensis: An insight through Structural Equation Modeling. Ann. Bot. Lond. 2019, 124, 1161–1171. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Star, G. Ecological forestry in the Southeast: Understanding the ecology of fuels. J. Forest. 2009, 107, 391–397. [Google Scholar] [CrossRef]
- Varner, J.M.; Kane, J.M.; Kreye, J.K.; Engber, E. The flammability of forest and woodland litter: A synthesis. Curr. For. Rep. 2015, 1, 91–99. [Google Scholar] [CrossRef]
- He, T.; Pausas, J.G.; Belcher, C.M.; Schwilk, D.W.; Lamont, B.B. Fire-adapted traits of Pinus arose in the fiery Cretaceous. New Phytol. 2012, 194, 751–759. [Google Scholar] [CrossRef]
- Keeley, J.E. Ecology and evolution of pine life histories. Ann. For. Sci. 2012, 69, 445–453. [Google Scholar] [CrossRef]
- Flanagan, S.A.; Bhotika, S.; Hawley, C.; Starr, G.; Wiesner, S.; Hiers, J.K.; O’Brien, J.J.; Goodrick, S.; Callaham, M.A.; Scheller, R.M.; et al. Quantifying carbon and species dynamics under different fire regimes in a southeastern U.S. pineland. Ecosphere 2019, 10, e02772. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Ackerly, D.D. Flammability and serotiny as strategies: Correlated evolution in pines. Oikos 2001, 94, 326–336. [Google Scholar] [CrossRef]
- Paine, C.E.T.; Stahl, C.; Courtois, E.A.; Patiño, S.; Sarmiento, C.; Baraloto, C. Functional explanations for variation in bark thickness in tropical rain forest trees. Funct. Ecol. 2010, 24, 1202–1210. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Gaetani, M.S.; Poulos, H.M. Oak bark allometry and fire survival strategies in the Chihuahuan Desert Sky Islands, Texas, USA. PLoS ONE 2013, 8, e79285. [Google Scholar] [CrossRef]
- Schafer, J.L.; Breslow, B.P.; Hohmann, M.G.; Hoffmann, W.A. Relative bark thickness is correlated with tree species distributions along a fire frequency gradient. Fire Ecol. 2015, 11, 74–87. [Google Scholar] [CrossRef]
- Pausas, J.G. Bark thickness and fire regime. New Phytol. 2017, 213, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Hengst, G.E.; Dawson, J.O. Bark properties and fire resistance of selected tree species from the central hardwood region of North America. Can. J. For. Res. 1994, 24, 688–696. [Google Scholar] [CrossRef]
- van Mantgem, P.; Schwartz, M. Bark heat resistance of small trees in Californian mixed conifer forests: Testing some model assumptions. For. Ecol. Manag. 2003, 178, 341–352. [Google Scholar] [CrossRef]
- Lawes, M.J.; Richards, A.E.; Dathe, J.; Midgley, J.J. Bark thickness determines fire resistance of selected tree species from fire-prone tropical savanna in north Australia. Plant Ecol. 2011, 212, 2057–2069. [Google Scholar] [CrossRef]
- VanderWeide, B.L.; Hartnett, D.C. Fire resistance of tree species explains historical gallery forest community composition. For. Ecol. Manag. 2011, 261, 1530–1538. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [PubMed]
- Stephens, S.L.; Libby, W.J. Anthropogenic fire and bark thickness in coastal and island pine populations from Alta and Baja California. J. Biogeogr. 2006, 33, 648–652. [Google Scholar] [CrossRef]
- Goubitz, S.; Nathan, R.; Roitemberg, R.; Shmida, A.; Nèman, G. Canopy seed bank structure in relation to: Fire, tree size and density. Plant Ecol. 2004, 173, 191–201. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Paula, S.; Pausas, J.G. Burning seeds: Germinative response to heat treatments in relation to resprouting ability. J. Ecol. 2008, 96, 543–552. [Google Scholar] [CrossRef]
- Gauthier, S.; Bergeron, Y.; Simon, J.P. Effects of fire regime on the serotiny level of jack pine. J. Ecol. 1996, 84, 539–548. [Google Scholar] [CrossRef]
- Vincenzi, S.; Piottisimone, A. Evolution of serotiny in maritime pine (Pinus pinaster) in the light of increasing frequency of fires. Plant Ecol. 2014, 215, 689–701. [Google Scholar] [CrossRef]
- Gómez-González, S.; Torres-Díaz, C.; Bustos-Schindler, C.; Gianoli, E. Anthropogenic fire drives the evolution of seed traits. Proc. Natl. Acad. Sci. USA 2011, 108, 18743–18747. [Google Scholar] [CrossRef]
- Gashaw, M.; Michelsen, A. Influence of heat shock on seed germination of plants from regularly burnt savanna woodlands and grasslands in Ethiopia. Plant Ecol. 2002, 159, 83–93. [Google Scholar] [CrossRef]
- Pausas, J.G.; Moreira, B. Flammability as a biological concept. New Phytol. 2012, 194, 610–613. [Google Scholar] [CrossRef]
- Boydak, M.; Çalışkan, S. Effects of heat shock on seed germination of Turkish red pine (Pinus brutia). Bosque 2016, 37, 327–333. [Google Scholar] [CrossRef]
- Tinker, D.B.; Romme, W.H.; Hargrove, W.W.; Gardner, R.H.; Turner, M.G. Landscape-scale heterogeneity in lodgepole pine serotiny. Can. J. For. Res. 1994, 24, 897–903. [Google Scholar] [CrossRef]
- Daskalakou, E.N.; Thanos, C.A. Aleppo pine (Pinus halepensis) postfire regeneration: The role of canopy and soil seed banks. Int. J. Wildland Fire 1996, 6, 59–66. [Google Scholar] [CrossRef]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of mediterranean pines. Plant Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Parchman, T.L.; Gompert, Z.; Mudge, J.; Schilkey, F.D.; Benkman, C.W.; Buerkle, C.A. Genome-wide association genetics of an adaptive trait in lodgepole pine. Mol. Ecol. 2012, 21, 2991–3005. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Serrano, A.; Verdú, M.; González-Martínez, S.C.; Pausas, J.G. Fire structures pine serotiny at different scales. Am. J. Bot. 2013, 100, 2349–2356. [Google Scholar] [CrossRef]
- Talluto, M.V.; Benkman, C.W. Landscape-scale eco–evolutionary dynamics: Selection by seed predators and fire determine a major reproductive strategy. Ecology 2013, 94, 1307–1316. [Google Scholar] [CrossRef]
- Talluto, M.V.; Benkman, C.W. Conflicting Selection From Fire And Seed Predation Drives Fine-Scaled Phenotypic Variation in a Widespread North American Conifer. Proc. Natl. Acad. Sci. USA 2014, 111, 9543–9548. [Google Scholar] [CrossRef]
- Pausas, J.G.; Su, W.; Luo, C.; Shen, Z. A shrubby resprouting pine with serotinous cones endemic to southwest China. Ecology 2021, 102, e03282. [Google Scholar] [CrossRef]
- Knapp, B.O.; Pile, L.S.; Walker, J.L.; Wang, G.G. Fire effects on a fire-adapted species: Response of grass stage longleaf pine seedlings to experimental burning. Fire Ecol. 2018, 14, 2. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Hiers, J.K.; Callaham, M.A.J.; Mitchell, R.J.; Jack, S.B. Interactions among overstory structure, seedling life-history traits, and fire in frequently burned neotropical pine forests. Ambio 2008, 37, 542–547. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Fonda, R.W.; Varner, J.M. Burning characteristics of needles from eight pine species. Northwest Sci. 2001, 78, 322–333. [Google Scholar] [CrossRef]
- Gagnon, P.R.; Passmore, H.A.; Platt, W.J.; Myers, J.A.; Paine, C.E.T.; Harms, K.E. Does pyrogenicity protect burning plants? Ecology 2010, 91, 3481–3486. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.R.; Belcher, C.M.; Bond, W.J.; Bradstock, R.A.; Daniau, A.-L.; Dexter, K.; Forrestel, E.J.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2017, 13, 033003. [Google Scholar] [CrossRef]
- Maxwell, C.J.; Serra-Diaz, J.M.; Scheller, R.M.; Thompson, J.R. Co-designed management scenarios shape the responses of seasonally dry forests to changing climate and fire regimes. J. Appl. Ecol. 2020, 57, 1328–1340. [Google Scholar] [CrossRef]
- Stephens, S.L.; Agee, J.K.; Fule, P.Z.; North, M.P.; Romme, W.H.; Swetnam, T.W.; Turner, M.G. Managing forests and fire in changing climates. Science 2013, 342, 41–42. [Google Scholar] [CrossRef]
- Knapp, B.O.; Stephan, K.; Hubbart, J.A. Structure and composition of an oak-hickory forest after over 60 years of repeated prescribed burning in Missouri, U.S.A. For. Ecol. Manag. 2015, 344, 95–109. [Google Scholar] [CrossRef]
- Hong, R.; Li, J.; Wang, J.; Zhu, X.; Li, X.; Ma, C.; Cao, H.; Wang, L.; Wang, Q. Effects of prescribed burning on understory Quercus species of Pinus yunnanensis forest. Front. For. Glob. Chang. 2023, 6, 1208682. [Google Scholar] [CrossRef]
- Wang, J.; Hong, R.; Ma, C.; Zhu, X.; Xu, S.; Tang, Y.; Li, X.; Yan, X.; Wang, L.; Wang, Q. Effects of Prescribed Burning on Surface Dead Fuel and Potential Fire Behavior in Pinus yunnanensis in Central Yunnan Province, China. Forests 2023, 14, 1915. [Google Scholar] [CrossRef]
- Capblancq, T.; Luu, K.; Blum, M.G.B.; Bazin, E. Evaluation of redundancy analysis to identify signatures of local adaptation. Mol. Ecol. Resour. 2018, 18, 1223–1233. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Liang, Y.; Shu, L. Altai forest fire danger area. In Forest Fire and Management in Altai, Xinjiang, 2nd ed.; Liang, Y., Shu, L., Eds.; Xinjiang People’s Press: Urumqi, China, 2021; pp. 176–202. [Google Scholar]
- Bond, W.J.; Midgley, J.J. Kill thy neighbour: An individualistic argument for the evolution of flammability. Oikos 1995, 73, 79–85. [Google Scholar] [CrossRef]
- Zhang, Y. Abies sibirica forest. In Forest in China, 2nd ed.; The Editorial Board of Forest in China; China Forestry Pubulishing House: Beijing, China, 1998; Volume 2, pp. 772–773. [Google Scholar]
- Shi, C.; Liang, Y.; Gao, C.; Wang, Q.; Shu, L. Drought-Modulated Boreal Forest Fire Occurrence and Linkage with La Nina Events in Altai Mountains, Northwest China. Atmosphere 2020, 11, 956. [Google Scholar] [CrossRef]
- Liu, X.; Pan, C. Analysis of Fire Severity of Kanas Taiga. Xinjiang Agric. Sci. 2020, 57, 1704–1712. [Google Scholar] [CrossRef]
- Zhang, F.; Pan, C.; Li, G.; Guo, K.; Liu, B.; Zou, Z. Quantity and quality change of edificators regeneration in Kanas Taiga after fire disturbance. Ecol. Environ. Sci. 2019, 28, 455–462. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Kreye, J.K.; Varner, J.M.; Hiers, J.K.; Mola, J. Toward a mechanism for eastern North American forest mesophication: Differential litter drying across 17 species. Ecol. Appl. 2013, 23, 1976–1986. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Abrupt climate-independent fire regime changes. Ecosystems 2014, 17, 1109–1120. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Wildfires as an ecosystem service. Front. Ecol. Environ. 2019, 17, 289–295. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Area | May | June | July | August | September |
|---|---|---|---|---|---|
| Habahe | 30.26% | 32.41% | 35.78% | 43.56% | 40.71% |
| Kanas | 39.67% | 41.57% | 44.34% | 59.62% | 48.87% |
| Buerjin | 43.42% | 55.78% | 63.81% | 67.23% | 52.50% |
| Species | Habahe | Kanas | Buerjin | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RA | RF | RD | IV | RA | RF | RD | IV | RA | RF | RD | IV | |
| Larix sibirica | 0.459 | 1.000 | 0.638 | 0.699 | 0.327 | 1.000 | 0.688 | 0.672 | 0.181 | 1.000 | 0.448 | 0.543 |
| Pinus sibirica | 0.393 | 1.000 | 0.330 | 0.574 | 0.320 | 0.900 | 0.205 | 0.475 | 0.530 | 1.000 | 0.520 | 0.683 |
| Picea obovata | 0.107 | 0.500 | 0.029 | 0.212 | 0.349 | 0.750 | 0.091 | 0.397 | 0.229 | 1.000 | 0.030 | 0.420 |
| Abies sibirica | 0.041 | 0.100 | 0.003 | 0.048 | 0.004 | 0.150 | 0.016 | 0.057 | 0.060 | 0.200 | 0.002 | 0.087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, R.; Liang, Y.; Wang, J.; Ma, C.; Zhu, X.; Xu, S.; Yang, X.; Yeerna, A.; Wang, W.; Wang, L.; et al. Adaptation of the Coniferous Forests to Natural Fire Disturbances in the Altai Mountains, Xinjiang, China. Forests 2024, 15, 296. https://doi.org/10.3390/f15020296
Hong R, Liang Y, Wang J, Ma C, Zhu X, Xu S, Yang X, Yeerna A, Wang W, Wang L, et al. Adaptation of the Coniferous Forests to Natural Fire Disturbances in the Altai Mountains, Xinjiang, China. Forests. 2024; 15(2):296. https://doi.org/10.3390/f15020296
Chicago/Turabian StyleHong, Ruicheng, Ying Liang, Jin Wang, Cheng Ma, Xilong Zhu, Shiying Xu, Xu Yang, Asiwuhan Yeerna, Wendong Wang, Leiguang Wang, and et al. 2024. "Adaptation of the Coniferous Forests to Natural Fire Disturbances in the Altai Mountains, Xinjiang, China" Forests 15, no. 2: 296. https://doi.org/10.3390/f15020296