Optimal Selection of Seed-Trees Using the Multi-Objective NSGA-II Algorithm and a Seed Dispersal Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. The Data-Set
2.2. Seed Dispersal Modeling
2.3. Optimization
- The relative number of seed-trees (Objective 1),
- The post-treatment relative seed-crop loss (Objective 2), and
- The post-treatment coefficient of variation in seed arrival across microsites (Objective 3).
2.3.1. Formulating the First Objective
2.3.2. Formulating the Second Objective
2.3.3. Formulating the Third Objective
2.3.4. NSGA-II Implementation
3. Results
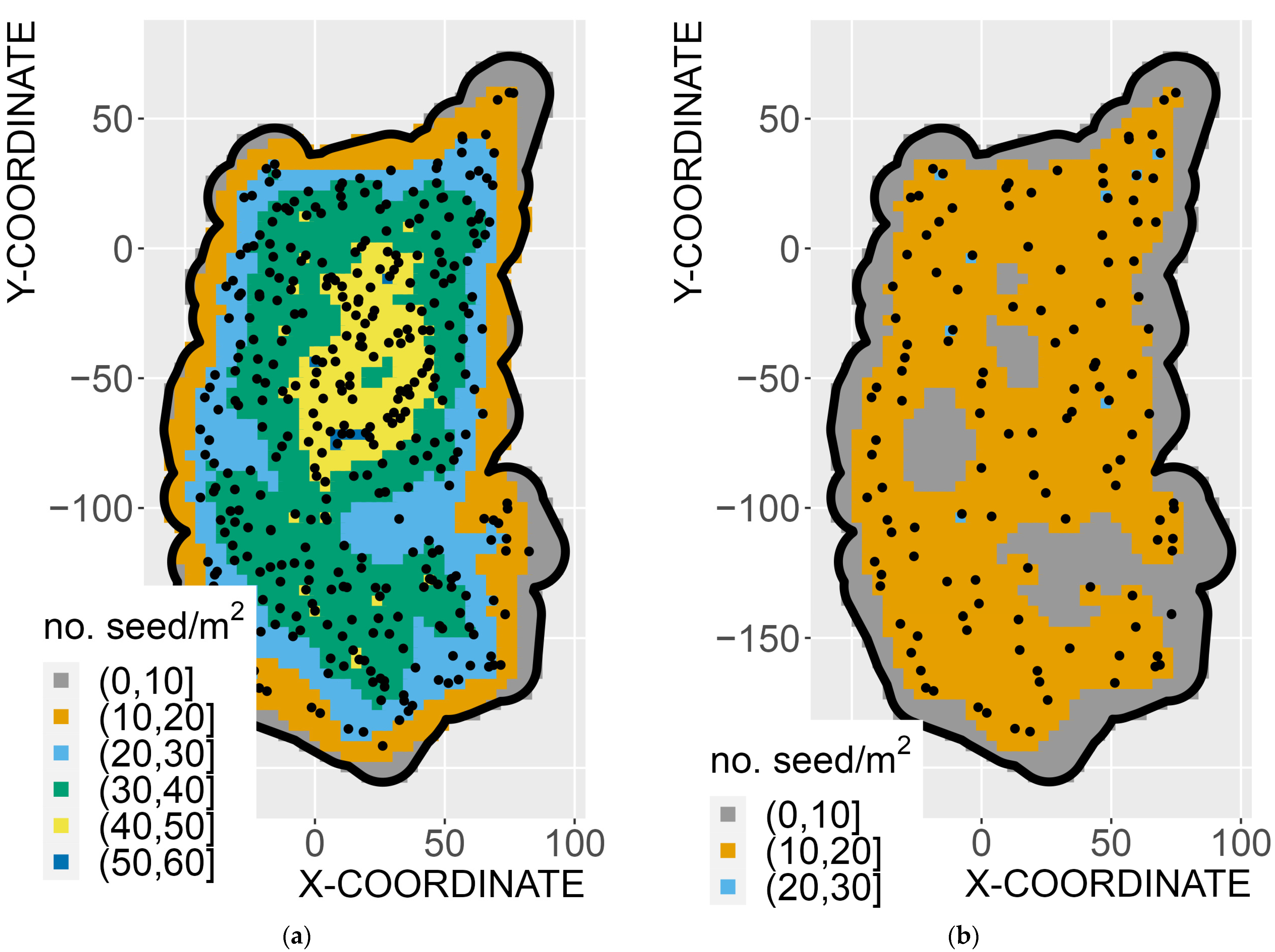
3.1. Seed-Dispersal Modeling
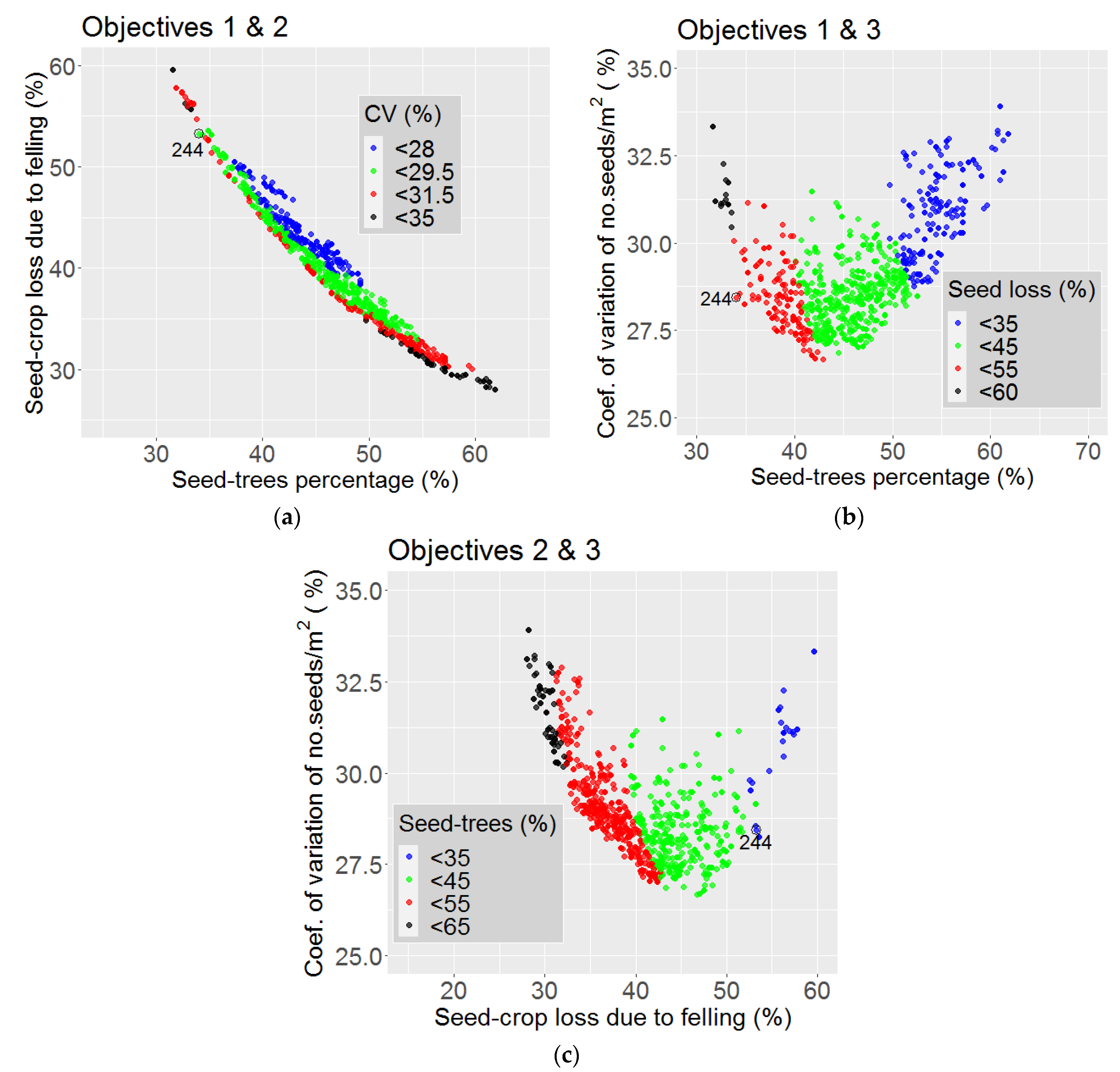
3.2. Optimization
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, K.P. Forest Management: Regulation and Valuation; McGraw-Hill Book Company: New York, NY, USA, 1966. [Google Scholar]
- Charco, J.; Venturas, M.; Gil, L.; Nanos, N. Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus Pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources. Forests 2017, 8, 312. [Google Scholar] [CrossRef]
- Stoehr, M.U. Seed Production of Western Larch in Seed-Tree Systems in the Southern Interior of British Columbia. For. Ecol. Manag. 2000, 130, 7–15. [Google Scholar] [CrossRef]
- Juez, L.; González-Martínez, S.C.; Nanos, N.; de-Lucas, A.I.; Ordóñez, C.; del Peso, C.; Bravo, F. Can Seed Production and Restricted Dispersal Limit Recruitment in Pinus Pinaster Aiton from the Spanish Northern Plateau? For. Ecol. Manag. 2014, 313, 329–339. [Google Scholar] [CrossRef]
- Calama, R.; Manso, R.; Lucas-Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo, F.; Peso, C.D.; Pardos, M. Natural Regeneration in Iberian Pines: A Review of Dynamic Processes and Proposals for Management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Guignabert, A.; Augusto, L.; Delerue, F.; Maugard, F.; Gire, C.; Magnin, C.; Niollet, S.; Gonzalez, M. Combining Partial Cutting and Direct Seeding to Overcome Regeneration Failures in Dune Forests. For. Ecol. Manag. 2020, 476, 118466. [Google Scholar] [CrossRef]
- Ettinger, A.K.; Lee, B.R.; Montgomery, S. Seed Limitation and Lack of Downed Wood, Not Invasive Species, Threaten Conifer Regeneration in an Urban Forest. Urban Ecosyst. 2017, 20, 877–887. [Google Scholar] [CrossRef]
- Lobo, N. Conifer Seed Predation by Terrestrial Small Mammals: A Review of the Patterns, Implications, and Limitations of Top-down and Bottom-up Interactions. For. Ecol. Manag. 2014, 328, 45–54. [Google Scholar] [CrossRef]
- Raymond, P.; Bédard, S.; Roy, V.; Larouche, C.; Tremblay, S. The Irregular Shelterwood System: Review, Classification, and Potential Application to Forests Affected by Partial Disturbances. J. For. 2009, 107, 405–413. [Google Scholar]
- Karlsson, M.; Nilsson, U. The Effects of Scarification and Shelterwood Treatments on Naturally Regenerated Seedlings in Southern Sweden. For. Ecol. Manag. 2005, 205, 183–197. [Google Scholar] [CrossRef]
- Cargill, J.; van Etten, E.J.B.; Stock, W.D. The Influence of Seed Supply and Seedbed on Seedling Recruitment in Shelterwood-Treated Jarrah (Eucalyptus Marginata) Forest. For. Ecol. Manag. 2019, 432, 54–63. [Google Scholar] [CrossRef]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and Management of Natural Regeneration of White Spruce in the Boreal Forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Nyland, R.D. Silviculture: Concepts and Applications, 3rd ed.; Waveland Press: Long Grove, IL, USA, 2016; ISBN 978-1-4786-3376-1. [Google Scholar]
- Zhou, W. Optimal Natural Regeneration of Scots Pine with Seed Trees. J. Environ. Manag. 1998, 53, 263–271. [Google Scholar] [CrossRef]
- Freitas, J.V.d.; Pinard, M.A. Applying Ecological Knowledge to Decisions about Seed Tree Retention in Selective Logging in Tropical Forests. For. Ecol. Manag. 2008, 256, 1434–1442. [Google Scholar] [CrossRef]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; HilleRisLambers, J. Seed Dispersal near and Far: Patterns across Temperate and Tropical Forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Clark, J.S.; Lewis, M.; McLachlan, J.S.; HilleRisLambers, J. Estimating Population Spread: What Can We Forecast and How Well? Ecology 2003, 84, 1979–1988. [Google Scholar] [CrossRef]
- Ribbens, E.; Silander, J.A.; Pacala, S.W. Seedling Recruitment in Forests: Calibrating Models to Predict Patterns of Tree Seedling Dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Condit, R.; Hubbell, S.P. Interspecific Variation in Primary Seed Dispersal in a Tropical Forest. J. Ecol. 2008, 96, 653–667. [Google Scholar] [CrossRef]
- Nanos, N.; Larson, K.; Millerón, M.; Sjöstedt-de Luna, S. Inverse Modeling for Effective Dispersal: Do We Need Tree Size to Estimate Fecundity? Ecol. Model. 2010, 221, 2415–2424. [Google Scholar] [CrossRef]
- Rogers, H.S.; Buhle, E.R.; HilleRisLambers, J.; Fricke, E.C.; Miller, R.H.; Tewksbury, J.J. Effects of an Invasive Predator Cascade to Plants via Mutualism Disruption. Nat. Commun. 2017, 8, 14557. [Google Scholar] [CrossRef]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of Long-Distance Seed Dispersal. Trends Ecol. Evol. 2008, 23, 638–647. [Google Scholar] [CrossRef]
- Kim, M.; Lee, S.; Lee, S.; Yi, K.; Kim, H.-S.; Chung, S.; Chung, J.; Kim, H.S.; Yoon, T.K. Seed Dispersal Models for Natural Regeneration: A Review and Prospects. Forests 2022, 13, 659. [Google Scholar] [CrossRef]
- Diaz-Balteiro, L.; Romero, C. Multiple Criteria Decision-Making in Forest Planning: Recent Results and Current Challenges. In Handbook of Operations Research in Natural Resources; Weintraub, A., Romero, C., Bjørndal, T., Epstein, R., Miranda, J., Eds.; International Series in Operations Research & Mana; Springer: Boston, MA, USA, 2007; pp. 473–488. ISBN 978-0-387-71815-6. [Google Scholar]
- Sun, Y.; Jin, X.; Pukkala, T.; Li, F. Two-Level Optimization Approach to Tree-Level Forest Planning. For. Ecosyst. 2022, 9, 100001. [Google Scholar] [CrossRef]
- Pascual, A. Multi-Objective Forest Planning at Tree-Level Combining Mixed Integer Programming and Airborne Laser Scanning. For. Ecol. Manag. 2021, 483, 118714. [Google Scholar] [CrossRef]
- Fransson, P.; Franklin, O.; Lindroos, O.; Nilsson, U.; Brännström, Å. A Simulation-Based Approach to a near-Optimal Thinning Strategy: Allowing Harvesting Times to Be Determined for Individual Trees. Can. J. For. Res. 2020, 50, 320–331. [Google Scholar] [CrossRef]
- Contreras, M.A.; Chung, W. Developing a Computerized Approach for Optimizing Individual Tree Removal to Efficiently Reduce Crown Fire Potential. For. Ecol. Manag. 2013, 289, 219–233. [Google Scholar] [CrossRef]
- Packalen, P.; Pukkala, T.; Pascual, A. Combining Spatial and Economic Criteria in Tree-Level Harvest Planning. For. Ecosyst. 2020, 7, 18. [Google Scholar] [CrossRef]
- Wing, B.M.; Boston, K.; Ritchie, M.W. A Technique for Implementing Group Selection Treatments with Multiple Objectives Using an Airborne Lidar-Derived Stem Map in a Heuristic Environment. For. Sci. 2019, 65, 211–222. [Google Scholar] [CrossRef]
- Fonseca, C.M.; Fleming, P.J. Genetic Algorithms for Multiobjective Optimization: Formulation, Discussion and Generalization. In Proceedings of the 5th International Conference on Genetic Algorithms, Urbana-Champaign, IL, USA, 1 June 1993; pp. 416–423. [Google Scholar]
- Deb, K.; Pratap, A.; Agarwal, S.; Meyarivan, T. A Fast and Elitist Multiobjective Genetic Algorithm: NSGA-II. IEEE Trans. Evol. Comput. 2002, 6, 182–197. [Google Scholar] [CrossRef]
- Katoch, S.; Chauhan, S.S.; Kumar, V. A Review on Genetic Algorithm: Past, Present, and Future. Multimed. Tools Appl. 2021, 80, 8091–8126. [Google Scholar] [CrossRef]
- Emmerich, M.T.M.; Deutz, A.H. A Tutorial on Multiobjective Optimization: Fundamentals and Evolutionary Methods. Nat. Comput. 2018, 17, 585–609. [Google Scholar] [CrossRef]
- Ducheyne, E.I.; De Wulf, R.R.; De Baets, B. A Spatial Approach to Forest-management Optimization: Linking GIS and Multiple Objective Genetic Algorithms. Int. J. Geogr. Inf. Sci. 2006, 20, 917–928. [Google Scholar] [CrossRef]
- Ducheyne, E.I.; De Wulf, R.R.; De Baets, B. Single versus Multiple Objective Genetic Algorithms for Solving the Even-Flow Forest Management Problem. For. Ecol. Manag. 2004, 201, 259–273. [Google Scholar] [CrossRef]
- Fotakis, D.G.; Sidiropoulos, E.; Myronidis, D.; Ioannou, K. Spatial Genetic Algorithm for Multi-Objective Forest Planning. For. Policy Econ. 2012, 21, 12–19. [Google Scholar] [CrossRef]
- López de Heredia, U.; Nanos, N.; García del Rey, E.; Guzmán, P.; López, R.; Venturas, M.; Gil Muñoz, P.; Gil Sánchez, L. High Seed Dispersal Ability of Pinus Canariensis in Stands of Contrasting Density Inferred from Genotypic Data. For. Syst. 2015, 24, 15. [Google Scholar] [CrossRef]
- Garcia-del-Rey, E.; Nanos, N.; López-de-Heredia, U.; Muñoz, P.G.; Otto, R.; Fernández-Palacios, J.M.; Gil, L. Spatiotemporal Variation of a Pinus Seed Rain Available for an Endemic Finch in an Insular Environment. Eur. J. Wildl. Res. 2011, 57, 337–347. [Google Scholar] [CrossRef]
- Peters, J.; González-Rodríguez, Á.M.; Jiménez, M.S.; Morales, D.; Wieser, G. Influence of Canopy Position, Needle Age and Season on the Foliar Gas Exchange of Pinus Canariensis. Eur. J. For. Res. 2008, 127, 293–299. [Google Scholar] [CrossRef]
- Proença-Ferreira, A.; Borda-de-Água, L.; Porto, M.; Mira, A.; Moreira, F.; Pita, R. Dispfit: An R Package to Estimate Species Dispersal Kernels. Ecol. Inform. 2023, 75, 102018. [Google Scholar] [CrossRef]
- Henningsen, A.; Toomet, O. maxLik: A Package for Maximum Likelihood Estimation in R. Comput. Stat. 2011, 26, 443–458. [Google Scholar] [CrossRef]
- Benitez, F.; Pinto-Roa, D. rmoo: Multi-Objective Optimization in R. R Package Version 0.2.0. 2022. Available online: https://CRAN.R-project.org/package=rmoo (accessed on 5 March 2024).
- Montoro Girona, M.; Lussier, J.-M.; Morin, H.; Thiffault, N. Conifer Regeneration After Experimental Shelterwood and Seed-Tree Treatments in Boreal Forests: Finding Silvicultural Alternatives. Front. Plant Sci. 2018, 9, 1145. [Google Scholar] [CrossRef]
- Bataineh, M.; Kenefic, L.; Weiskittel, A.; Wagner, R.; Brissette, J. Influence of Partial Harvesting and Site Factors on the Abundance and Composition of Natural Regeneration in the Acadian Forest of Maine, USA. For. Ecol. Manag. 2013, 306, 96–106. [Google Scholar] [CrossRef]
- Bose, A.K.; Weiskittel, A.; Wagner, R.G.; Kuehne, C. Assessing the Factors Influencing Natural Regeneration Patterns in the Diverse, Multi-Cohort, and Managed Forests of Maine, USA. J. Veg. Sci. 2016, 27, 1140–1150. [Google Scholar] [CrossRef]
- González-González, J.M.; Vázquez-Méndez, M.E.; Diéguez-Aranda, U. Multi-Objective Models for the Forest Harvest Scheduling Problem in a Continuous-Time Framework. For. Policy Econ. 2022, 136, 102687. [Google Scholar] [CrossRef]
- López, E.; Climent, J.M.; Montero, G. Selvicultura de Pinus Canariensis Sweet. In Compendio de Selvicultura Aplicada en España; INIA: Madrid, Spain, 2008. [Google Scholar]
- Mavrotas, G. Effective Implementation of the ε-Constraint Method in Multi-Objective Mathematical Programming Problems. Appl. Math. Comput. 2009, 213, 455–465. [Google Scholar] [CrossRef]
- Simonsen, R. Optimal Regeneration Method-Planting vs. Natural Regeneration of Scots Pine in Northern Sweden. Silva Fenn. 2013, 47, 928. [Google Scholar] [CrossRef]
- Gonzalez Molina, J.M.; Piqué Nicolau, M.; Grau, P.V. Manual de Ordenación Por Rodales. Gestión Multifuncional de los Espacios Forestales; Organismo Autόnomo de Parques Nacionales: Madrid, Spain, 2011; ISBN 978-84-8014-789-7. [Google Scholar]
- Axer, M.; Martens, S.; Schlicht, R.; Wagner, S. Modelling Natural Regeneration of European Beech in Saxony, Germany: Identifying Factors Influencing the Occurrence and Density of Regeneration. Eur. J. For. Res. 2021, 140, 947–968. [Google Scholar] [CrossRef]
- Kohler, M.; Pyttel, P.; Kuehne, C.; Modrow, T.; Bauhus, J. On the Knowns and Unknowns of Natural Regeneration of Silviculturally Managed Sessile Oak (Quercus petraea (Matt.) Liebl.) Forests—A Literature Review. Ann. For. Sci. 2020, 77, 101. [Google Scholar] [CrossRef]
- Bose, A.K.; Alcalá-Pajares, M.; Kern, C.C.; Montoro-Girona, M.; Thiffault, N. Complex Regeneration Responses of Eight Tree Species to Partial Harvest in Mixedwood Forests of Northeastern North America. For. Ecol. Manag. 2023, 529, 120672. [Google Scholar] [CrossRef]
- Beckman, N.G.; Sullivan, L.L. The Causes and Consequences of Seed Dispersal. Annu. Rev. Ecol. Evol. Syst. 2023, 54, 403–427. [Google Scholar] [CrossRef]
- Vander Wall, S.B. Seed Dispersal in Pines (Pinus). Bot. Rev. 2023, 89, 275–307. [Google Scholar] [CrossRef]
- Ruano, I.; Manso, R.; Fortin, M.; Bravo, F. Extreme Climate Conditions Limit Seed Availability to Successfully Attain Natural Regeneration of Pinus Pinaster in Sandy Areas of Central Spain. Can. J. For. Res. 2015, 45, 1795–1802. [Google Scholar] [CrossRef]
- Millerón, M.; López De Heredia, U.; Lorenzo, Z.; Perea, R.; Dounavi, A.; Alonso, J.; Gil, L.; Nanos, N. Effect of Canopy Closure on Pollen Dispersal in a Wind-Pollinated Species (Fagus sylvatica L.). Plant Ecol. 2012, 213, 1715–1728. [Google Scholar] [CrossRef]
- Schupp, E.W.; Jordano, P.; Gómez, J.M. Seed Dispersal Effectiveness Revisited: A Conceptual Review. New Phytol. 2010, 188, 333–353. [Google Scholar] [CrossRef] [PubMed]
- Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Queinnec, M.; Luther, J.E.; Bolton, D.K.; White, J.C.; Wulder, M.A.; van Lier, O.R.; Hermosilla, T. Modelling Lidar-Derived Estimates of Forest Attributes over Space and Time: A Review of Approaches and Future Trends. Remote Sens. Environ. 2021, 260, 112477. [Google Scholar] [CrossRef]
- Kwak, D.-A.; Lee, W.-K.; Lee, J.-H.; Biging, G.S.; Gong, P. Detection of Individual Trees and Estimation of Tree Height Using LiDAR Data. J. For. Res. 2007, 12, 425–434. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Model | Lklhd | AIC | r | Maes | Merr | MSerr | p1 | p2 | p3 |
|---|---|---|---|---|---|---|---|---|---|---|
| 2007 | Lognormal | −344.035 | 694.070 | 0.484 | 4.462 | 0.00065 | 29.293 | 2.94 | 0.87 | 17,113.3 |
| Gamma | −344.129 | 694.259 | 0.486 | 4.460 | 0.00010 | 29.261 | 8.26 | 2.43 | 16,079.5 | |
| 2Dt | −344.289 | 694.578 | 0.486 | 4.461 | −0.00071 | 29.289 | 30.35 | 3.40 | 16,210.1 | |
| Weibull | −344.421 | 694.842 | 0.483 | 4.466 | 0.00011 | 29.317 | 21.52 | 1.87 | 15,952.0 | |
| Wald | −344.480 | 694.960 | 0.466 | 4.481 | −0.00091 | 29.468 | 39.60 | 20.97 | 18,560.5 | |
| Geometric | −344.563 | 695.126 | 0.489 | 4.464 | 0.00019 | 29.350 | 149,690 | 13,106 | 16,458.0 | |
| 2008 | Weibull | −329.581 | 665.162 | 0.467 | 3.676 | 0.00036 | 21.666 | 21.84 | 1.56 | 11,150.5 |
| Gamma | −329.706 | 665.412 | 0.458 | 3.685 | 0.00047 | 21.703 | 10.55 | 1.89 | 11,190.5 | |
| Geometric | −329.748 | 665.497 | 0.457 | 3.689 | −0.00054 | 21.720 | 60,276 | 6269 | 11,125.1 | |
| Lognormal | −330.214 | 666.427 | 0.441 | 3.707 | 0.00089 | 21.835 | 2.82 | 0.93 | 11,719.3 | |
| Wald | −330.894 | 667.788 | 0.428 | 3.755 | −0.00434 | 21.997 | 23.43 | 23.00 | 11,482.0 | |
| 2Dt | −330.990 | 667.980 | 0.485 | 3.695 | 0.00090 | 21.915 | 1,947.01 | 6,597 | 11,147.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanos, N.; Garcia-del-Rey, E.; Gil, L. Optimal Selection of Seed-Trees Using the Multi-Objective NSGA-II Algorithm and a Seed Dispersal Model. Forests 2024, 15, 499. https://doi.org/10.3390/f15030499
Nanos N, Garcia-del-Rey E, Gil L. Optimal Selection of Seed-Trees Using the Multi-Objective NSGA-II Algorithm and a Seed Dispersal Model. Forests. 2024; 15(3):499. https://doi.org/10.3390/f15030499
Chicago/Turabian StyleNanos, Nikos, Eduardo Garcia-del-Rey, and Luis Gil. 2024. "Optimal Selection of Seed-Trees Using the Multi-Objective NSGA-II Algorithm and a Seed Dispersal Model" Forests 15, no. 3: 499. https://doi.org/10.3390/f15030499
APA StyleNanos, N., Garcia-del-Rey, E., & Gil, L. (2024). Optimal Selection of Seed-Trees Using the Multi-Objective NSGA-II Algorithm and a Seed Dispersal Model. Forests, 15(3), 499. https://doi.org/10.3390/f15030499