
Effects of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions and Soil C and N Pools in Chinese Hickory Plantation Forests


Abstract
1. Introduction
2. Materials and Methods
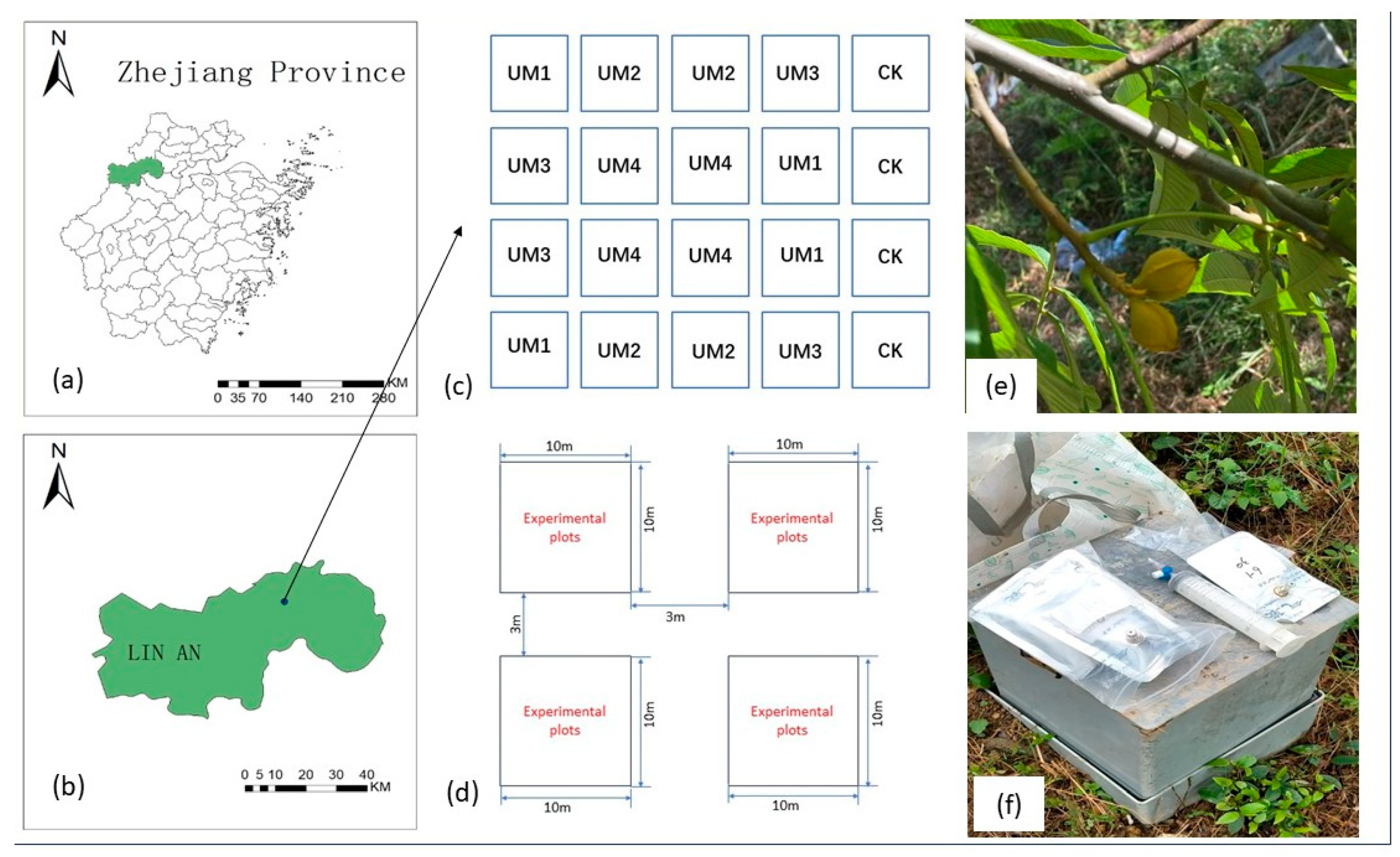
2.1. General Description of the Study Area
2.2. Experimental Design
2.3. Measurement of GHG Emissions from Soils
2.4. Measurement Methods for Soil Physico-Chemical Properties
2.5. Statistics and Analyses
3. Results
3.1. Impact of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions
3.2. Effects of Understory Vegetation Conversion on Soil Environmental Factors
3.3. Relationship between Soil GHG Emissions and Soil Environmental Factors under Forest Vegetation Conversion
4. Discussion
4.1. Impact of Understory Vegetation Conversion on Soil GHG Emissions
4.2. Effects of Understory Vegetation Conversion on Soil C and N Pools
4.3. Influence of Soil Environmental Factors on Soil GHG Emissions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamb, W.F.; Wiedmann, T.; Pongratz, J.; Andrew, R.; Crippa, M.; Olivier, J.G.J.; Wiedenhofer, D.; Mattioli, G.; Khourdajie, A.A.; House, J.; et al. A review of trends and drivers of greenhouse gas emissions by sector from 1990 to 2018. Environ. Res. Lett. 2021, 16, 073005. [Google Scholar] [CrossRef]
- Rajeev, J.; Hukum, S. Carbon sequestration potential of disturbed and non-disturbed forest ecosystem: A tool for mitigating climate change. Afr. J. Environ. Sci. Technol. 2020, 14, 385–393. [Google Scholar] [CrossRef]
- Qureshi, A.; Kumar, A.; Lal Nag, J. Precision Agriculture and Carbon Sequestration—Need of an Hour in Current Climate Changing Scenario. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1668–1673. [Google Scholar] [CrossRef]
- Graves, R.A.; Haugo, R.D.; Holz, A.; Nielsen-Pincus, M.; Jones, A.; Kellogg, B.; Macdonald, C.; Popper, K.; Schindel, M. Potential greenhouse gas reductions from Natural Climate Solutions in Oregon, USA. PLoS ONE 2020, 15, e0230424. [Google Scholar] [CrossRef] [PubMed]
- Mäkipää, R.; Abramoff, R.; Adamczyk, B.; Baldy, V.; Biryol, C.; Bosela, M.; Casals, P.; Curiel Yuste, J.; Dondini, M.; Filipek, S.; et al. How does management affect soil C sequestration and greenhouse gas fluxes in boreal and temperate forests?—A review. For. Ecol. Manag. 2023, 529, 120637. [Google Scholar] [CrossRef]
- Xu, L.; Deng, X.; Ying, J.; Zhou, G.; Shi, Y. Silicate fertilizer application reduces soil greenhouse gas emissions in a Moso bamboo forest. Sci. Total Environ. 2020, 747, 141380. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, P.; Wang, H.; Zhou, G.; Wu, J.; Yang, F.; Qian, X. Seasonal soil CO2 efflux dynamics after land use change from a natural forest to Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2011, 262, 1131–1137. [Google Scholar] [CrossRef]
- McIntosh, A.C.; Macdonald, S.E.; Quideau, S.A. Understory Plant Community Composition Is Associated with Fine-Scale Above- and Below-Ground Resource Heterogeneity in Mature Lodgepole Pine (Pinus contorta) Forests. PLoS ONE 2016, 11, e0151436. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Xiao, R.; Cai, T.; Man, X.; Ge, Z.; Gao, M.; Mencuccini, M. Understory species composition mediates soil greenhouse gas fluxes by affecting bacterial community diversity in boreal forests. Front. Microbiol. 2022, 13, 1090169. [Google Scholar] [CrossRef]
- Li, H.-F. Soil CH4 fluxes response to understory removal and N-fixing species addition in four forest plantations in Southern China. J. For. Res. 2010, 21, 301–310. [Google Scholar] [CrossRef]
- Xiang, H.; Zhang, Y.B.; Wei, H.; Zhang, J.-E.; Zhao, B.-L. Soil properties and carbon and nitrogen pools in a young hillside longan orchard after the introduction of leguminous plants and residues. PeerJ 2018, 6, e5536. [Google Scholar] [CrossRef]
- Das Gupta, S.; Pinno, B.D. Drivers of understory species richness in reconstructed boreal ecosystems: A structural equation modeling analysis. Sci. Rep. 2020, 10, 11555. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, K.; Zhang, J.; Li, D.; Zhang, Y.; Xiang, H. Grass cultivation alters soil organic carbon fractions in a subtropical orchard of southern China. Soil Tillage Res. 2018, 181, 110–116. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Chen, G.; Guo, J.; Xie, J.-S.; Wang, X.-G. Soil respiration and carbon balance in a subtropical native forest and two managed plantations. Plant Ecol. 2007, 193, 71–84. [Google Scholar] [CrossRef]
- Jin, J.; Wang, L.; Müller, K.; Wu, J.-S.; Wang, H.; Zhao, K.L.; Berninger, F.; Fu, W. A 10-year monitoring of soil properties dynamics and soil fertility evaluation in Chinese hickory plantation regions of southeastern China. Sci. Rep. 2021, 11, 23531. [Google Scholar] [CrossRef]
- Wu, J.; Lin, H.; Meng, C.; Jiang, P.; Fu, W. Effects of intercropping grasses on soil organic carbon and microbial community functional diversity under Chinese hickory (Carya cathayensis Sarg.) stands. Soil Res. 2014, 52, 575–583. [Google Scholar] [CrossRef]
- Jin, J.; Huang, X.; Wu, J.; Zhao, W.; Fu, W. A 10-year field experiment proves the neutralization of soil pH in Chinese hickory plantation of southeastern China. J. Soils Sediments 2022, 22, 2995–3005. [Google Scholar] [CrossRef]
- Xu, L.; Fang, H.; Deng, X.; Ying, J.; Lv, W.; Shi, Y.; Zhou, G.; Zhou, Y. Biochar application increased ecosystem carbon sequestration capacity in a Moso bamboo forest. For. Ecol. Manag. 2020, 475, 118447. [Google Scholar] [CrossRef]
- Singh, B.P.; Hatton, B.J.; Balwant, S.; Cowie, A.L.; Kathuria, A. Influence of biochars on nitrous oxide emission and nitrogen leaching from two contrasting soils. J. Environ. Qual. 2010, 39, 1224–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Fan, C.H.; Li, Q.L.; Li, B.; Zhu, Y.Y.; Xiong, Z.Q. A 2-yr field assessment of the effects of chemical and biological nitrification inhibitors on nitrous oxide emissions and nitrogen use efficiency in an intensively managed vegetable cropping system. Agric. Ecosyst. Environ. 2015, 201, 43–50. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chang, S.X.; Jiang, P.; Zhou, G.; Liu, J.; Wu, J.; Shen, Z. Understory vegetation management affected greenhouse gas emissions and labile organic carbon pools in an intensively managed Chinese chestnut plantation. Plant Soil 2013, 376, 363–375. [Google Scholar] [CrossRef]
- Rao, X.; Liu, C.-A.; Tang, J.-W.; Nie, Y.; Liang, M.-Y.; Shen, W.-J.; Siddique, K.H.M. Rubber-leguminous shrub systems stimulate soil N2O but reduce CO2 and CH4 emissions. For. Ecol. Manag. 2021, 480, 118665. [Google Scholar] [CrossRef]
- Tang, X.; Fan, S.-h.; Qi, L.; Guan, F.-Y.; Du, M.; Zhang, H. Soil respiration and net ecosystem production in relation to intensive management in Moso bamboo forests. Catena 2016, 137, 219–228. [Google Scholar] [CrossRef]
- Xiao, H.B.; Shi, Z.H.; Li, Z.W.; Chen, J.; Huang, B.; Yue, Z.J.; Zhan, Y.M. The regulatory effects of biotic and abiotic factors on soil respiration under different land-use types. Ecol. Indic. 2021, 127, 107787. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Chen, D.; Huang, G.; Zhou, L.; Fu, S. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biol. Biochem. 2011, 43, 2355–2357. [Google Scholar] [CrossRef]
- Yuze, S.; Li, Y.; Cai, Y.; Fu, S.; Luo, Y.; Wang, H.; Liang, C.; Lin, Z.; Hu, S.; Li, Y.; et al. Biochar decreases soil N2O emissions in Moso bamboo plantations through decreasing labile N concentrations, N-cycling enzyme activities and nitrification/denitrification rates. Geoderma 2019, 348, 135–145. [Google Scholar]
- Toma, Y.; Nufita Sari, N.; Akamatsu, K.; Oomori, S.; Nagata, O.; Nishimura, S.; Purwanto, B.; Ueno, H. Effects of Green Manure Application and Prolonging Mid-Season Drainage on Greenhouse Gas Emission from Paddy Fields in Ehime, Southwestern Japan. Agriculture 2019, 9, 29. [Google Scholar] [CrossRef]
- Hu, S.; Li, Y.; Chang, S.X.; Li, Y.; Yang, W.; Fu, W.; Liu, J.; Jiang, P.; Lin, Z. Soil autotrophic and heterotrophic respiration respond differently to land-use change and variations in environmental factors. Agric. For. Meteorol. 2018, 250–251, 290–298. [Google Scholar] [CrossRef]
- Wardinski, K.M.; Hotchkiss, E.R.; Jones, C.N.; McLaughlin, D.L.; Strahm, B.D.; Scott, D.T. Water-Soluble Organic Matter From Soils at the Terrestrial-Aquatic Interface in Wetland-Dominated Landscapes. J. Geophys. Res. Biogeosci. 2022, 127, e2022JG006994. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chang, S.X.; Qin, H.; Fu, S.; Jiang, P. Understory management and fertilization affected soil greenhouse gas emissions and labile organic carbon pools in a Chinese chestnut plantation. For. Ecol. Manag. 2015, 337, 126–134. [Google Scholar] [CrossRef]
- Karavanova, E.I.; Zolovkina, D.F. The Effect of Litter Composition on the Characteristics of Their Water-Soluble Organic Matter. Mosc. Univ. Soil Sci. Bull. 2020, 75, 67–73. [Google Scholar] [CrossRef]
- Wu, Y.-p.; Wang, X.; Hu, R.; Zhao, J.; Jiang, Y. Responses of Soil Microbial Traits to Ground Cover in Citrus Orchards in Central China. Microorganisms 2021, 9, 2507. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Fu, S.; Wang, X.; Wang, M.; Liang, C.; Chen, Y.; Zhu, X. Effects of understory removal and nitrogen fertilization on soil microbial communities in Eucalyptus plantations. For. Ecol. Manag. 2013, 310, 80–86. [Google Scholar] [CrossRef]
- Yuan, B.; Yu, D.; Hu, A.; Wang, Y.; Sun, Y.; Li, C. Effects of green manure intercropping on soil nutrient content and bacterial community structure in litchi orchards in China. Front. Environ. Sci. 2023, 10, 1059800. [Google Scholar] [CrossRef]
- Zhao, Q.; Classen, A.T.; Wang, W.-W.; Zhao, X.-R.; Mao, B.; Zeng, D.-H. Asymmetric effects of litter removal and litter addition on the structure and function of soil microbial communities in a managed pine forest. Plant Soil 2017, 414, 81–93. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, Y.; Yang, A.; Zhang, Y.; Lu, M.; Ge, S.; Qiu, Y.; Zhang, J.; Tong, Z.-k. Understory vegetation restoration improves soil physicochemical properties, enzymatic activity, and changes diazotrophic communities in Cunninghamia lanceolata plantations but depends on site history. Plant Soil 2023, 492, 605–623. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Hu, G.; Wu, H.; Liang, Y.; Yan, J.; He, S.; Ganjurjav, H.; Gao, Q. Reclamation intensifies the positive effects of warming on N2O emission in an alpine meadow. Front. Plant Sci. 2023, 14, 1162160. [Google Scholar] [CrossRef] [PubMed]
- Truu, M.; Nõlvak, H.; Ostonen, I.; Oopkaup, K.; Maddison, M.; Ligi, T.; Espenberg, M.; Uri, V.; Mander, Ü.; Truu, J. Soil Bacterial and Archaeal Communities and Their Potential to Perform N-Cycling Processes in Soils of Boreal Forests Growing on Well-Drained Peat. Front. Microbiol. 2020, 11, 591358. [Google Scholar] [CrossRef]
- Zhang, J.; Peng, C.; Zhu, Q.; Xue, W.; Shen, Y.; Yang, Y.; Shi, G.; Shi, S.; Wang, M. Temperature sensitivity of soil carbon dioxide and nitrous oxide emissions in mountain forest and meadow ecosystems in China. Atmos. Environ. 2016, 142, 340–350. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Hai, X.; Wu, J.; Dong, L.; Pan, Y.; Shangguan, Z.; Wang, K.; Deng, L. Carbon inputs regulate the temperature sensitivity of soil respiration in temperate forests. J. Arid. Land 2022, 14, 1055–1068. [Google Scholar] [CrossRef]
- Fang, H.J.; Yu, G.R.; Cheng, S.L.; Zhu, T.H.; Wang, Y.S.; Yan, J.H.; Wang, M.; Cao, M.; Zhou, M. Effects of multiple environmental factors on CO2 emission and CH4 uptake from old-growth forest soils. Biogeosciences 2010, 7, 395–407. [Google Scholar] [CrossRef]
- Du, M.; Yuan, J.; Zhuo, M.; Sadiq, M.; Wu, J.; Xu, G.; Liu, S.; Li, J.; Li, G.; Yan, L. Effects of different land use patterns on soil properties and N2O emissions on a semi-arid Loess Plateau of Central Gansu. Front. Ecol. Evol. 2023, 11, 1128236. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, S.; Shen, G.; Shaaban, M.; Ju, W.; Cui, Y.; Duan, C.; Fang, L. Effects of inorganic and organic fertilizers on CO2 and CH4 fluxes from tea plantation soil. Elem. Sci. Anthr. 2021, 9, 090. [Google Scholar] [CrossRef]
- Geng, J.; Cheng, S.; Fang, H.; Yu, G.; Li, X.; Si, G.; He, S.; Yu, G. Soil nitrate accumulation explains the nonlinear responses of soil CO2 and CH4 fluxes to nitrogen addition in a temperate needle-broadleaved mixed forest. Ecol. Indic. 2017, 79, 28–36. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weather Environment | 2022 | 2023 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jul. | Aug. | Spe. | Oct. | Nov. | Dec | Jan. | Feb. | Mar. | Apr. | May. | Jun. | |
| Average monthly temperature (°C) | 30.5 | 31 | 24 | 17.5 | 15 | −4.5 | −5.5 | 7 | 12.5 | 17 | 21.5 | 25.5 |
| Precipitation (mm) | 68.2 | 20 | 25.8 | 47.4 | 27.3 | 42.9 | 55.5 | 63.3 | 50.4 | 137.5 | 121.1 | 204.6 |
| GHG | Treatment | Model | df | R2 | p |
|---|---|---|---|---|---|
| CO2 | UM1 | Y = 0.745T | 48 | 0.545 | ** |
| Y = 0.826T − 0.222WSON | 48 | 0.579 | ** | ||
| Y = 1.018T − 0.278WSON − 273WSOC | 48 | 0.615 | ** | ||
| Y = 1.251T − 0.392WSON − 426WSOC + 0.282NH4+–N | 48 | 0.662 | ** | ||
| UM2 | Y = 0.857T | 48 | 0.729 | ** | |
| Y = 1.044T − 0.303MBN | 48 | 0.783 | ** | ||
| UM3 | Y = 0.878T | 48 | 0.766 | ** | |
| Y = 1.054T − 0.246MBC | 48 | 0.792 | ** | ||
| Y = 1.223T − 0.372MBC − 0.234WSON | 48 | 0.831 | ** | ||
| Y = 1.229T − 0.409MBC − 0.179WSON + 0.136pH | 48 | 0.842 | ** | ||
| UM4 | Y = 0.884T | 48 | 0.777 | ** | |
| Y = 1.019T − 0.234NO3−–N | 48 | 0.811 | ** | ||
| Y = 1.143T − 0.375NO3−–N + 0.293NH4+–N | 48 | 0.882 | ** | ||
| CK | Y = 0.881T | 48 | 0.772 | ** | |
| Y = 0.701T + 0.315WSOC | 48 | 0.837 | ** | ||
| Y = 0.723T + 0.358WSOC − 0.224WSON | 48 | 0.883 | ** |
| GHG | Treatment | Model | df | R2 | p |
|---|---|---|---|---|---|
| N2O | UM1 | Y = 0.712MBC | 48 | 0.496 | ** |
| Y = 0.807MBC − 0.229NO3−–N | 48 | 0.531 | ** | ||
| Y = 0.456MBC − 0.305NO3−–N + 0.461T | 48 | 0.587 | ** | ||
| Y = 0.370MBC − 0.409NO3−–N + 0.688T + 0.315NH4+–N | 48 | 0.661 | ** | ||
| Y = 0.598MBC − 0.515NO3−–N + 0.802T + 0.422NH4+–N − 0.345WSOC | 48 | 0.697 | ** | ||
| UM2 | Y = 0.729T | 48 | 0.521 | ** | |
| Y = 0.720T + 0.334NH4+–N | 48 | 0.627 | ** | ||
| Y = 0.824T + 0.458NH4+–N − 0.333WSON | 48 | 0.708 | ** | ||
| UM3 | Y = 0.712T | 48 | 0.496 | ** | |
| Y = 0.785T + 0.388M | 48 | 0.635 | ** | ||
| Y = 0.755T + 0.424M + 0.237NH4+–N | 48 | 0.685 | ** | ||
| Y = 0.805T + 0.298M + 0.230NH4+–N − 0.215WSON | 48 | 0.706 | ** | ||
| Y = 1.108T + 0.302M + 0.147NH4+–N − 0.313WSON − 0.358MBC | 48 | 0.750 | ** | ||
| UM4 | Y = 0.718T | 48 | 0.505 | ** | |
| Y = 0.754T + 0.397M | 48 | 0.657 | ** | ||
| Y = 0.788T + 0.441M + 0.206NH4+–N | 48 | 0.692 | ** | ||
| CK | Y = 0.685WSOC | 48 | 0.457 | ** | |
| Y = 0.441WSOC + 0.427T | 48 | 0.573 | ** | ||
| Y = 0.503WSOC + 0.458T − 0.324WSON | 48 | 0.668 | ** | ||
| Y = 0.551WSOC + 0.715T − 0.369WSON−0.377MBN | 48 | 0.730 | ** | ||
| Y = 0.436WSOC + 0.777T − 0.329WSON − 0.349MBN + 0.194M | 48 | 0.756 | ** |
| GHG | Treatment | Model | df | R2 | p |
|---|---|---|---|---|---|
| CH4 | UM1 | Y = 0.665MBC | 48 | 0.430 | ** |
| Y = 0.805MBC − 0.339NO3−–N | 48 | 0.518 | ** | ||
| Y = 0.602MBC − 0.360NO3−–N + 0.362MBN | 48 | 0.599 | ** | ||
| UM2 | Y = 0.706MBC | 48 | 0.488 | ** | |
| Y = 0.468MBC + 0.422MBN | 48 | 0.603 | ** | ||
| Y = 0.479MBC + 0.542MBN − 0.263NO3−–N | 48 | 0.651 | ** | ||
| Y = 0.472MBC + 0.577MBN − 0.254NO3−–N − 0.176M | 48 | 0.675 | ** | ||
| Y = 0.489MBC + 0.656MBN − 0.411NO3−–N − 0.237M + 0.249NH4+–N | 48 | 0.711 | ** | ||
| UM3 | Y = 0.681MBC | 48 | 0.466 | ** | |
| Y = 0.685MBC − 0.278WSON | 48 | 0.535 | ** | ||
| Y = 0.250MBC − 0.492WSON + 0.603T | 48 | 0.666 | ** | ||
| UM4 | Y = 0.730MBC | 48 | 0.523 | ** | |
| Y = 0.663MBC + 0.279M | 48 | 0.589 | ** | ||
| Y = 0.377MBC + 0.314M + 0.354MBN | 48 | 0.630 | ** | ||
| CK | Y = 0.665T | 48 | 0.431 | ** | |
| Y = 0.792T + 0.296NH4+–N | 48 | 0.493 | ** | ||
| Y= 0.520T + 0.346NH4+–N + 0.429MBC | 48 | 0.583 | ** | ||
| Y = 0.805T + 0.486NH4+–N + 0.397MBC − 0.358WSOC | 48 | 0.649 | ** | ||
| Y = 0.900T + 0.562NH4+–N + 0.255MBC − 0.454WSOC + 0.308PH | 48 | 0.719 | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Shi, H.; Chen, Y.; Huang, S.; Wang, E.; Ni, Z.; Zhou, Y.; Shi, Y. Effects of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions and Soil C and N Pools in Chinese Hickory Plantation Forests. Forests 2024, 15, 558. https://doi.org/10.3390/f15030558
Gao Y, Shi H, Chen Y, Huang S, Wang E, Ni Z, Zhou Y, Shi Y. Effects of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions and Soil C and N Pools in Chinese Hickory Plantation Forests. Forests. 2024; 15(3):558. https://doi.org/10.3390/f15030558
Chicago/Turabian StyleGao, Yanyan, Haitao Shi, Yangen Chen, Sha Huang, Enhui Wang, Zelong Ni, Yufeng Zhou, and Yongjun Shi. 2024. "Effects of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions and Soil C and N Pools in Chinese Hickory Plantation Forests" Forests 15, no. 3: 558. https://doi.org/10.3390/f15030558
APA StyleGao, Y., Shi, H., Chen, Y., Huang, S., Wang, E., Ni, Z., Zhou, Y., & Shi, Y. (2024). Effects of Understory Vegetation Conversion on Soil Greenhouse Gas Emissions and Soil C and N Pools in Chinese Hickory Plantation Forests. Forests, 15(3), 558. https://doi.org/10.3390/f15030558