
Effects of Anthropogenic Disturbance on the Structure, Competition, and Succession of Abies ziyuanensis Communities
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Research Methods
2.2.1. Site Investigation and Data Processing
- (1)
- The canopy layer. We surveyed all trees and large shrubs with a diameter at breast height (DBH) of 4 cm or greater. We recorded factors including tree height, DBH, location, count, canopy width, and species attributes.
- (2)
- The understory layer. We established four 5 m × 5 m subplots at each plot corner. This layer included saplings taller than 10 cm and with DBH < 4 cm, as well as all shrubs not investigated in the canopy layer. The recorded factors were plant height, cover, count, and species attributes.
- (3)
- The ground layer. We placed five 1 m × 1 m subplots evenly at each plot’s corners and center. This layer targeted all herbaceous plants and tree seedlings less than 10 cm in height. The recorded factors included plant height, cover, count, and species attributes.
2.2.2. Analysis of Structural Characteristics of the Abies ziyuanensis Communities
- (1)
- Diameter Class Structure Characteristics of Abies ziyuanensis Species
- (2)
- Species Importance Value Index
- (3)
- Species Diversity Characteristic Indices
2.2.3. Analysis of Species Composition Differences in Communities
- (1)
- Extreme difference: 0.50 R 1.00 and p-value ≤ 0.05, indicating a vast difference in species composition between the two forest conditions;
- (2)
- Significant difference: 0.50 and p-value ≤ 0.05, showing a clear distinction between the two forest conditions;
- (3)
- Moderate difference: 0.50 and p-value ≤ 0.075, where the two forest conditions partially overlap but are distinct to a certain degree;
- (4)
- Slight difference: p-value ≤ 0.10, where, although the forest conditions are similar, some differences still exist;
- (5)
- Almost no difference: p-value where the two forest conditions can be considered extremely similar.
2.2.4. Analysis of Individual Competition Differences in the Abies ziyuanensis Communities
3. Results
3.1. Description of Plant Genus Information
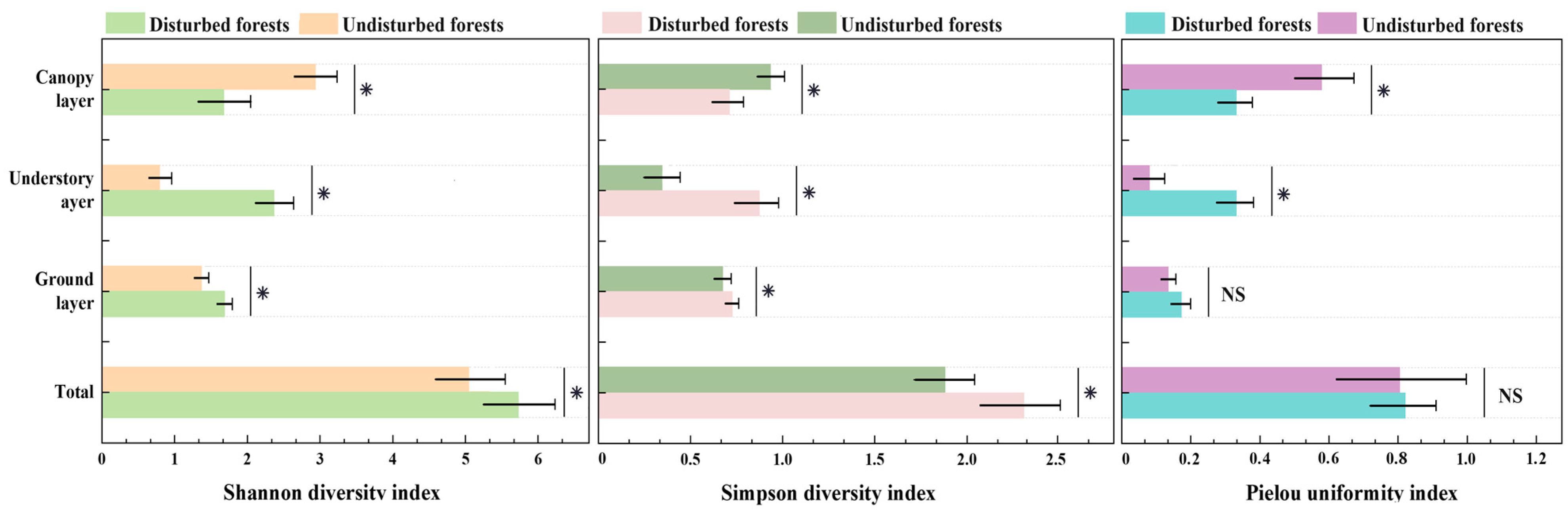
3.2. Structural Characteristics and Species Diversity
3.3. Species Composition Differences in Abies ziyuanensis Communities
3.4. Single-Tree Competition in Abies ziyuanensis Communities
4. Discussion
4.1. Species Diversity of Abies ziyuanensis Communities
4.2. Spatial Competition among Abies ziyuanensis Trees
4.3. Growth Environment of Abies ziyuanensis Communities
4.4. Disturbance Effects on Succession of Abies ziyuanensis Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tang, C.Q.; Yang, Y.; Momohara, A.; Wang, H.C.; Luu, H.T.; Li, S.; Song, X.; LePage, B.; Dong, Y.F.; Han, P.B.; et al. Forest Characteristics and Population Structure of Glyptostrobus pensilis, a Globally Endangered Relict Species of Southeastern China. Plant Divers. 2019, 41, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Manchester, S.R.; Chen, Z.D.; Lu, A.M.; Uemura, K. Eastern Asian Endemic Seed Plant Genera and Their Paleogeographic History throughout the Northern Hemisphere. J. Syst. Evol. 2009, 47, 1–42. [Google Scholar] [CrossRef]
- Qian, S.; Yang, Y.; Tang, C.Q.; Momohara, A.; Yi, S.; Ohsawa, M. Effective Conservation Measures Are Needed for Wild Cathaya argyrophylla Populations in China: Insights from the Population Structure and Regeneration Characteristics. For. Ecol. Manag. 2016, 361, 358–367. [Google Scholar] [CrossRef]
- Boscutti, F.; Casolo, V.; Beraldo, P.; Braidot, E.; Zancani, M. Shrub Growth and Plant Diversity along an Elevation Gradient: Evidence of Indirect Effects of Climate on Alpine Ecosystems. PLoS ONE 2018, 13, e0196653. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Zhang, X.; Xu, X.; Yi, H.; He, J.; He, L. Beta Diversity of Plant Communities in Relation to Soil C:N:P Stoichiometry across 150 Years of Vegetation Restoration in a Temperate Zone. Forests 2023, 14, 553. [Google Scholar] [CrossRef]
- Yousefzadeh, H.; Amirchakhmaghi, N.; Naseri, B.; Shafizadeh, F.; Kozlowski, G.; Walas, Ł. The impact of climate change on the future geographical distribution range of the endemic relict tree Gleditsia caspica (Fabaceae) in Hyrcanian forests. Ecol. Inform. 2022, 71, 101773. [Google Scholar] [CrossRef]
- Stevens, G.C. The Elevational Gradient in Altitudinal Range: An Extension of Rapoport’s Latitudinal Rule to Altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. The Use of ‘Altitude’ in Ecological Research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.F.; Chen, H.Y.H. Is Understory Plant Species Diversity Driven by Resource Quantity or Resource Heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef]
- Kim, Y.J.; Song, Y.K.; Lee, D.K. Understanding the Critical Impact Path on Vegetation Growth under Climate Extremes and Human Influence. Forests 2019, 10, 947. [Google Scholar] [CrossRef]
- McGinley, K.A.; Robertson, G.C.; Friday, K.S. Examining the Sustainability of Tropical Island Forests: Advances and Challenges in Measurement, Monitoring, and Reporting in the U.S. Caribbean and Pacific. Forests 2019, 10, 946. [Google Scholar] [CrossRef]
- Su, X.; Wang, M.; Huang, Z.; Fu, S.; Chen, H.Y.H. Forest Understorey Vegetation: Colonization and the Availability and Heterogeneity of Resources. Forests 2019, 10, 944. [Google Scholar] [CrossRef]
- Finegan, B. Forest succession. Nature 1984, 312, 109–114. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2018-1. Available online: https://www.iucnredlist.org (accessed on 20 August 2018).
- Li, X.X.; Tao, C.; Wang, Q.C.; Cui, G.F. Characteristics of Geographic Distribution of Four Critically Endangered Species of Abies in Subtropical China and Its Relationship with Climate. Chin. J. Plant Ecol. 2012, 36, 1154. [Google Scholar] [CrossRef]
- Zhang, Y.R. Studies on Endangered Mechanism and Population Conservation of Abies ziyuanensis. PhD Dissertation, Beijing Forestry University, Beijing, China, 2009. [Google Scholar]
- Fu, Z.X.; Wang, X.Y.; Fan, P.Z.; Tian, X.Y.; Shao, Y.Z. The Complete Chloroplast Genome of the Endangered Pinaceae Species Abies Ziyuanensis and Its Phylogenetic Implications. Mitochondrial DNA Part B 2019, 4, 137–138. [Google Scholar] [CrossRef]
- Davies, G.M.; Gray, A. Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecol. Evol. 2015, 5, 5295–5304. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Song, Y.; Pang, Y.; Kang, H.; Xue, Y.; Wang, D. Driving Factors of Chinese Pine Population Distribution in the Ridge Habitats of the Southern Slope of the Mid-Qinling Mountains, China. Forests 2023, 14, 2252. [Google Scholar] [CrossRef]
- Li, W.; Zhang, G.F. Population Structure and Spatial Pattern of the Endemic and Endangered Subtropical Tree Parrotia subaequalis (Hamamelidaceae). Flora-Morphol. Distrib. Funct. Ecol. Plants 2015, 212, 10–18. [Google Scholar] [CrossRef]
- He, C.; Jia, S.; Luo, Y.; Hao, Z.; Yin, Q. Spatial Distribution and Species Association of Dominant Tree Species in Huangguan Plot of Qinling Mountains, China. Forests 2022, 13, 866. [Google Scholar] [CrossRef]
- Obiang, N.L.E.; Kenfack, D.; Picard, N.; Lutz, J.A.; Bissiengou, P.; Memiaghe, H.R.; Alonso, A. Determinants of Spatial Patterns of Canopy Tree Species in a Tropical Evergreen Forest in Gabon. J. Veg. Sci. 2019, 30, 929–939. [Google Scholar] [CrossRef]
- Ahmed, M.; Shaukat, S.S. A Textbook of Vegetation Ecology; Abrar Sons: Karachi, Pakistan, 2012; pp. 1–396. [Google Scholar]
- Alam, N.M.; Shaheen, H.; Manzoor, M.; Tinghong, T.; Arfan, M.; Idrees, M. Spatial Distribution and Population Structure of Himalayan Fir (Abies pindrow (Royle ex D. Don) Royle) in Moist Temperate Forests of the Kashmir Region. Forests 2023, 14, 482. [Google Scholar] [CrossRef]
- Pereki, H.; Wala, K.; Thiel-Clemen, T.; Bessike, M.P.B.; Zida, M.; Dourma, M. Woody Species Diversity and Important Value Indices in Dense Dry Forests in Abdoulaye Wildlife Reserve (Togo, West Africa). Int. J. Biodivers. Conserv. 2013, 5, 358–366. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; John and Wiley and Sons: Hoboken, NJ, USA, 2017; pp. 1–15. ISBN 9781118445112. [Google Scholar] [CrossRef]
- Oksanen, J. Vegan: Community Ecology Package. 2010. Available online: https://r-forge.r-project.org/projects/vegan (accessed on 18 November 2022).
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Hui, G.; Wang, Y.; Zhang, G.; Zhao, Z.; Bai, C.; Liu, W. A Novel Approach for Assessing the Neighborhood Competition in Two Different Aged Forests. For. Ecol. Manag. 2018, 422, 49–58. [Google Scholar] [CrossRef]
- Hegyi, F. A Simulation Model for Managing Jack-Pine Standssimulation. R. Coll. For. Res. Notes 1974, 30, 74–90. [Google Scholar]
- Pang, L.; Wang, G.; Sharma, R.P.; Lu, J.; Tang, X.; Fu, L. Simulation of Thinning by Integrating Tree Competition and Species Biodiversity for Target Tree-Based Management of Secondary Forests. Forests 2023, 14, 1896. [Google Scholar] [CrossRef]
- Sandoval, S.; Cancino, J. Modeling the Edge Effect in Even-Aged Monterrey Pine (Pinus radiata D. Don) Stands Incorporating a Competition Index. For. Ecol. Manag. 2008, 256, 78–87. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, Z.; Li, Q.; He, L.; Gao, Y.; Wang, L.; Han, D. A Structural Equation Model Suggests That Soil Physical Properties Had the Greatest Negative Influence on the Competition Index of Dominant Trees of Pinus sylvestris var. Mongolia. Forests 2023, 14, 1191. [Google Scholar] [CrossRef]
- Bhandari, S.K.; Veneklaas, E.J.; McCaw, L.; Mazanec, R.; Whitford, K.; Renton, M. Individual Tree Growth in Jarrah (Eucalyptus marginata) Forest is Explained by Size and Distance of Neighbouring Trees in Thinned and Non-Thinned Plots. For. Ecol. Manag. 2021, 494, 119364. [Google Scholar] [CrossRef]
- Amat, M.E.; Vargas, P.; Gomez, J.M. Effects of Human Activity on the Distribution and Abundance of an Endangered Mediterranean High-Mountain Plant (Erysimum penyalarense). J. Nat. Conserv. 2013, 21, 262–271. [Google Scholar] [CrossRef]
- Lombardi, F.; Lella, S.D.; Altieri, V.; Di Benedetto, S.; Giancola, C.; Lasserre, B.; Kutnar, L.; Tognetti, R.; Marchetti, M. Early Responses of Biodiversity Indicators to Various Thinning Treatments in Mountain Beech Forests. iForest-Biogeosci. For. 2018, 11, 609. [Google Scholar] [CrossRef]
- Von-Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsch, I.; Yu, M.; Härdtle, W. Individual-Tree Radial Growth in a Subtropical Broad-Leaved Forest: The Role of Local Neighbourhood Competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Qiu, H.; Zhang, H.; Lei, K.; Hu, X.; Yang, T.; Jiang, X. A New Tree-Level Multi-Objective Forest Harvest Model (MO-PSO): Integrating Neighborhood Indices and PSO Algorithm to Improve the Optimization Effect of Spatial Structure. Forests 2023, 14, 441. [Google Scholar] [CrossRef]
- Song, M.; Hu, Q.; Tian, Y.; Ouyang, H. Seasonal Patterns of Root and Shoot Interactions in an Alpine Meadow on the Tibetan Plateau. J. Plant Ecol. 2012, 5, 182–190. [Google Scholar] [CrossRef]
- Kullman, L. Rapid Recent Range-Margin Rise of Tree and Shrub Species in the Swedish Scandes. J. Ecol. 2002, 90, 68–77. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub Expansion in Tundra Ecosystems: Dynamics, Impacts and Research Priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Sturm, M.; Racine, C.; Tape, K. Climate Change: Increasing Shrub Abundance in the Arctic. Nature 2001, 411, 546–547. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Popadiouk, R.V. Dynamics of North American boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Taylor, A.R.; Gao, B.; Chen, H.Y. The effect of species diversity on tree growth varies during forest succession in the boreal forest of central Canada. For. Ecol. Manag. 2020, 455, 117641. [Google Scholar] [CrossRef]
- Wagner, S.; Fischer, H.; Huth, F. Canopy effects on vegetation caused by harvesting and regeneration treatments. Eur. J. For. Res. 2011, 130, 17–40. [Google Scholar] [CrossRef]
- O. Knapp, B.; G. Pallardy, S. Forty-Eight Years of Forest Succession: Tree Species Change across Four Forest Types in Mid-Missouri. Forests 2018, 9, 633. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Plot Number | Site | Longitude | Latitude | Elevation (m) | Slope (°) | Canopy Cover | Conditions of Communities |
|---|---|---|---|---|---|---|---|
| SHM1 | Shunhuang Mountains | 111°00.695′ | 26°22.266′ | 1713.03 | 38 | 0.70 | Undisturbed |
| EDM2 | Erbaoding Mountains | 110°32.410′ | 26°16.602′ | 1783.12 | 24 | 0.80 | Undisturbed |
| JGS3 | Jinggang Mountains | 114°06.689′ | 26°30.136′ | 1657.71 | 20 | 0.90 | Undisturbed |
| DYF4 | Dayuan Farm | 114°01.793′ | 26°24.470′ | 1501.10 | 18 | 0.95 | Undisturbed |
| DYF5 | Dayuan Farm | 114°01.727′ | 26°24.476′ | 1483.27 | 9 | 0.70 | Disturbed |
| DYF6 | Dayuan Farm | 114°03.170′ | 26°26.240′ | 1467.00 | 18 | 0.75 | Disturbed |
| DYF7 | Dayuan Farm | 114°02.824′ | 26°25.910′ | 1407.85 | 2 | 0.85 | Disturbed |
| EDM8 | Erbaoding Mountains | 110°30.466′ | 26°18.233′ | 1490.34 | 35 | 0.65 | Disturbed |
| Conditions of Communities | Vegetation Layer | Number of Family | Number of Genus | Number of Species |
|---|---|---|---|---|
| Undisturbed forests | Canopy | 19 | 25 | 31 |
| Understory | 14 | 16 | 16 | |
| Ground | 9 | 9 | 9 | |
| Total | 32 | 41 | 48 | |
| Disturbed forests | Canopy | 14 | 17 | 17 |
| Understory | 15 | 16 | 16 | |
| Ground | 11 | 14 | 14 | |
| Total | 29 | 39 | 40 | |
| All total | 39 | 60 | 71 | |
| Conditions of Communities | Level Ⅰ | Level Ⅱ | Level Ⅲ | Level Ⅳ | Level V | Level Ⅵ | Level Ⅶ |
|---|---|---|---|---|---|---|---|
| Undisturbed forests | 0 | 0 | 0 | 5 | 9 | 1 | 4 |
| Disturbed forests | 603 | 1 | 2 | 19 | 23 | 12 | 1 |
| Total | 603 | 1 | 2 | 24 | 32 | 13 | 5 |
| Community Conditions | Sample Plot Number | DBH of Abies ziyuanensis (cm) | DBH of all Tree (cm) | Number of Trees | Number of Abies ziyuanensis | of Abies ziyuanensis | of all Tree |
|---|---|---|---|---|---|---|---|
| Undisturbed forests | SHM1 | 44.8 | 18.3 | 53 | 4 | 5.33 | 16.12 |
| EDM2 | 8.2 | 14.5 | 32 | 2 | 9.40 | 8.87 | |
| JGS3 | 20.0 | 9.1 | 69 | 1 | 4.05 | 14.77 | |
| DYF4 | 19.0 | 12.1 | 38 | 12 | 5.38 | 9.21 | |
| Mean | 23.00 | 13.5 | 48 | 5 | 6.04 | 12.24 | |
| Disturbed forests | DYF5 | 33.5 | 23.1 | 17 | 7 | 2.76 | 5.26 |
| DYF6 | 11.2 | 12.5 | 58 | 30 | 21.10 | 17.68 | |
| DYF7 | 10.9 | 10.9 | 15 | 14 | 3.57 | 3.42 | |
| EDM8 | 27.3 | 22.5 | 27 | 7 | 3.61 | 3.35 | |
| Mean | 20.7 | 17.2 | 29 | 15 | 7.76 | 7.43 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Li, Y.; Xu, K.; Yu, L.; He, P.; Zeng, S.; Song, Y.; Liu, R.; Sun, Y. Effects of Anthropogenic Disturbance on the Structure, Competition, and Succession of Abies ziyuanensis Communities. Forests 2024, 15, 1001. https://doi.org/10.3390/f15061001
Zhang H, Li Y, Xu K, Yu L, He P, Zeng S, Song Y, Liu R, Sun Y. Effects of Anthropogenic Disturbance on the Structure, Competition, and Succession of Abies ziyuanensis Communities. Forests. 2024; 15(6):1001. https://doi.org/10.3390/f15061001
Chicago/Turabian StyleZhang, Huacong, Yueqiao Li, Keqin Xu, Longhua Yu, Ping He, Suping Zeng, Yunxia Song, Ren Liu, and Yun Sun. 2024. "Effects of Anthropogenic Disturbance on the Structure, Competition, and Succession of Abies ziyuanensis Communities" Forests 15, no. 6: 1001. https://doi.org/10.3390/f15061001