
Construction of Compatible Volume Model for Populus in Beijing, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Data
2.1.1. Study Site
2.1.2. Data Collection
2.2. Statistical Analysis
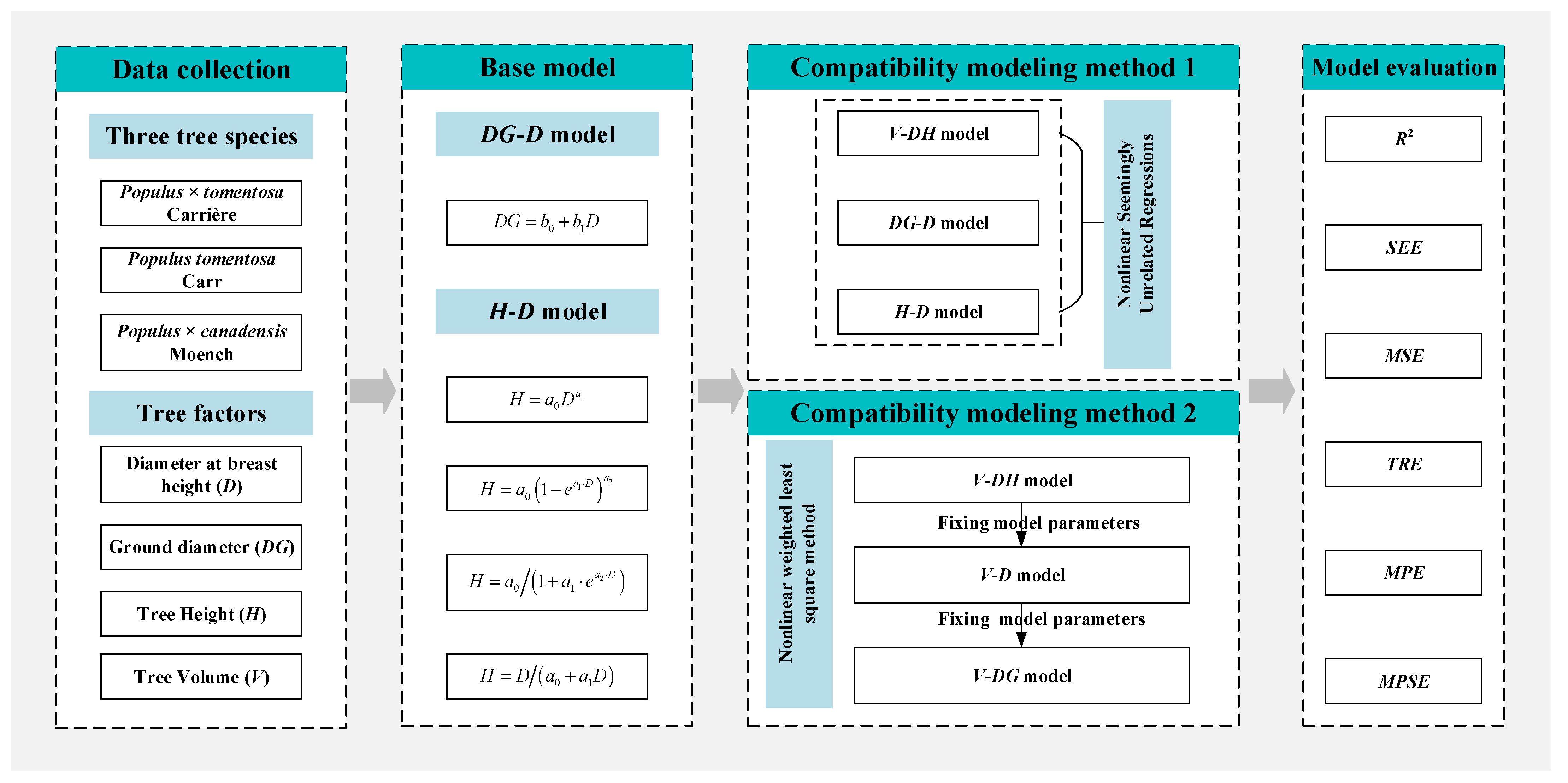
2.2.1. Model Form Selection
2.2.2. Compatible Volume Model
2.2.3. Model Evaluation and Validation
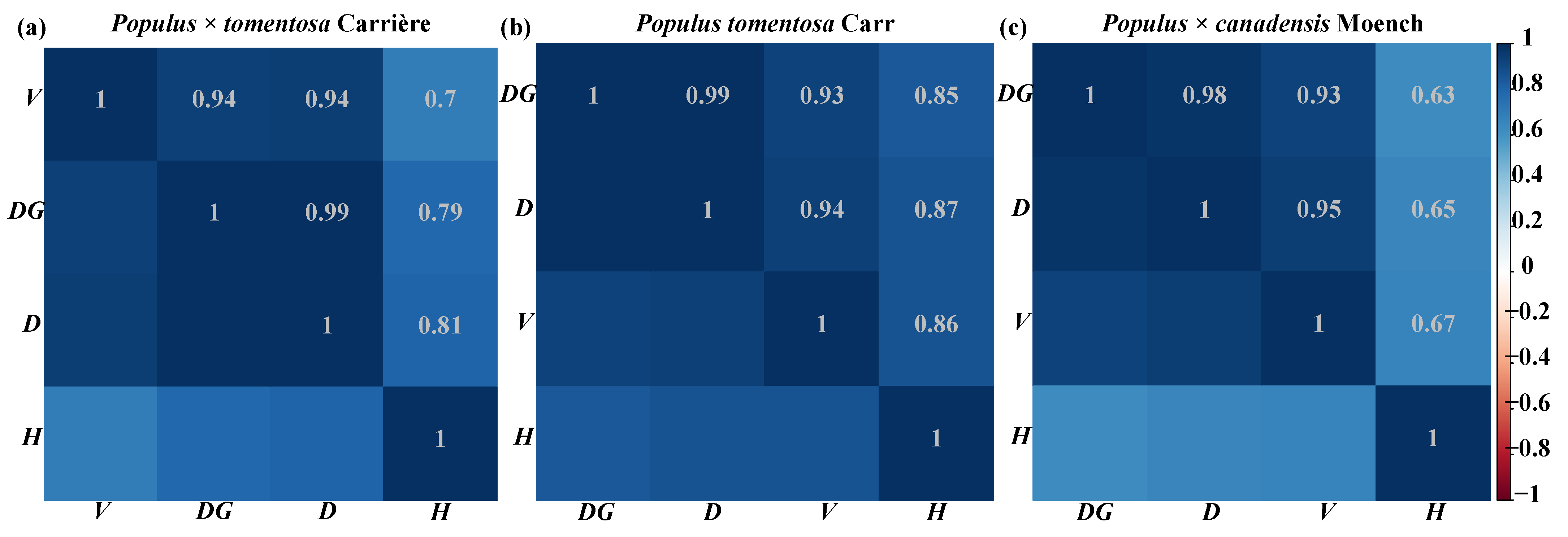
2.2.4. Correlation Analysis
3. Results
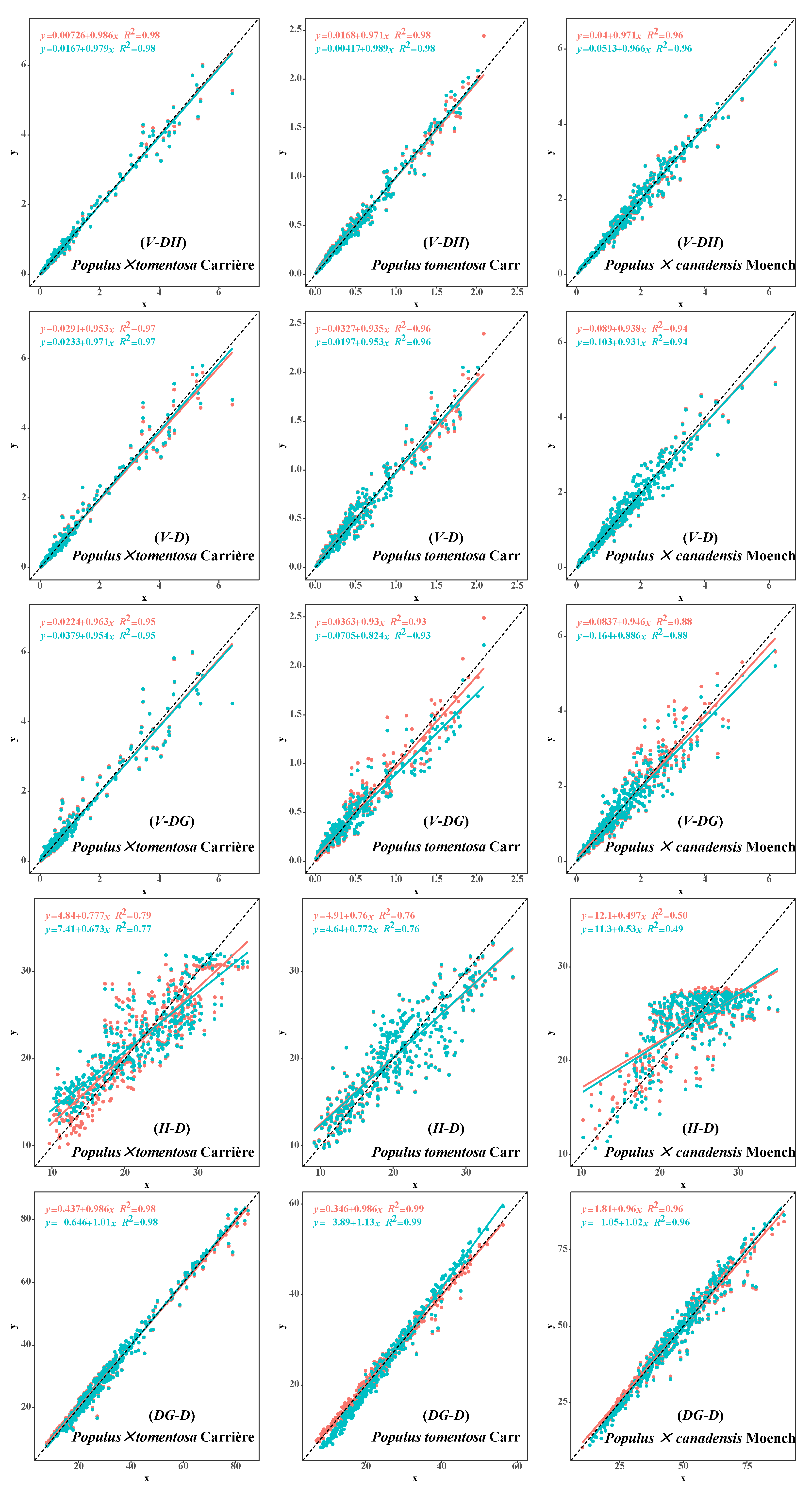
3.1. Correlation Analysis
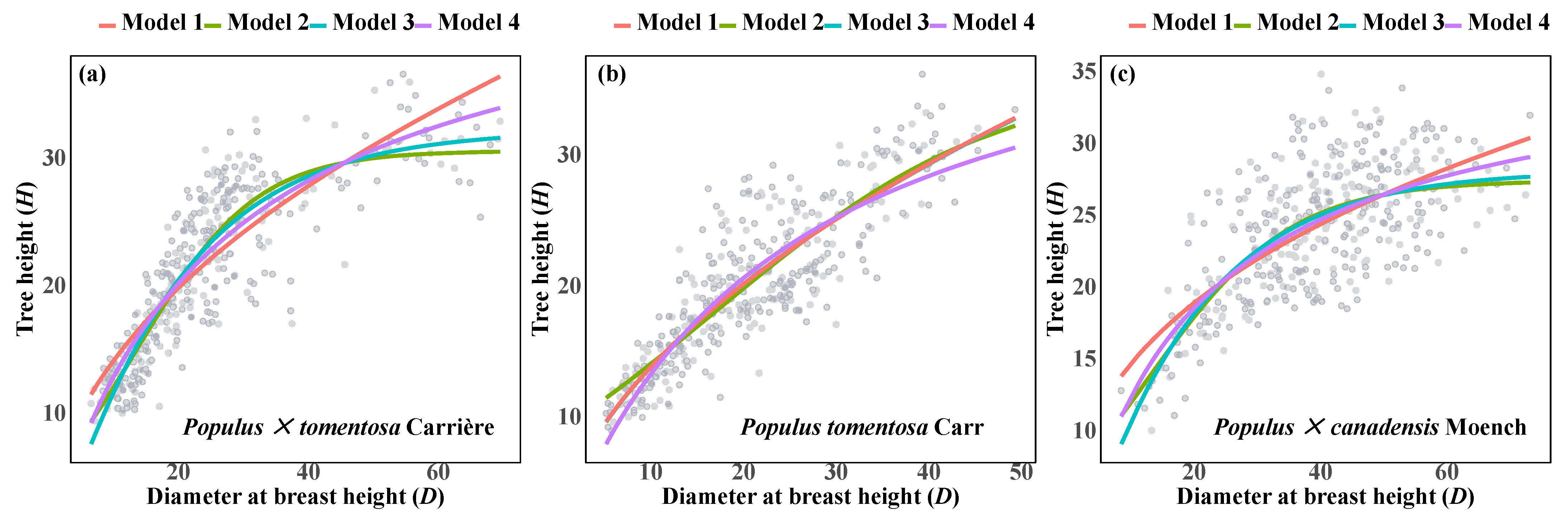
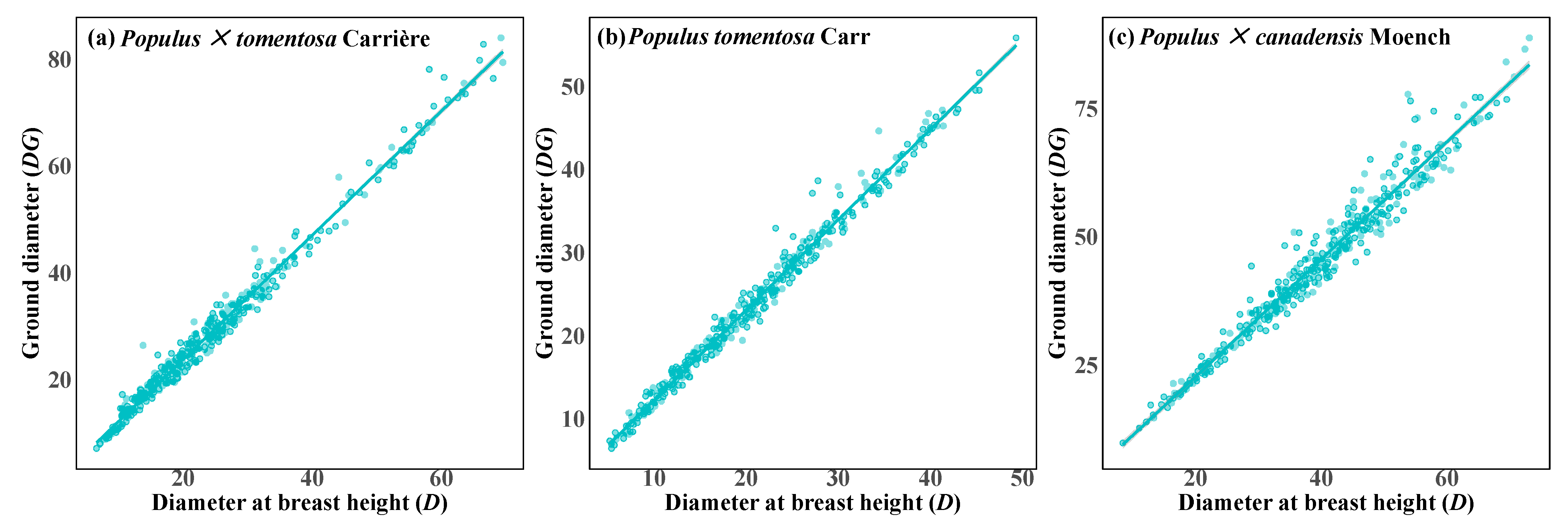
3.2. Independent Fitting Model Analysis
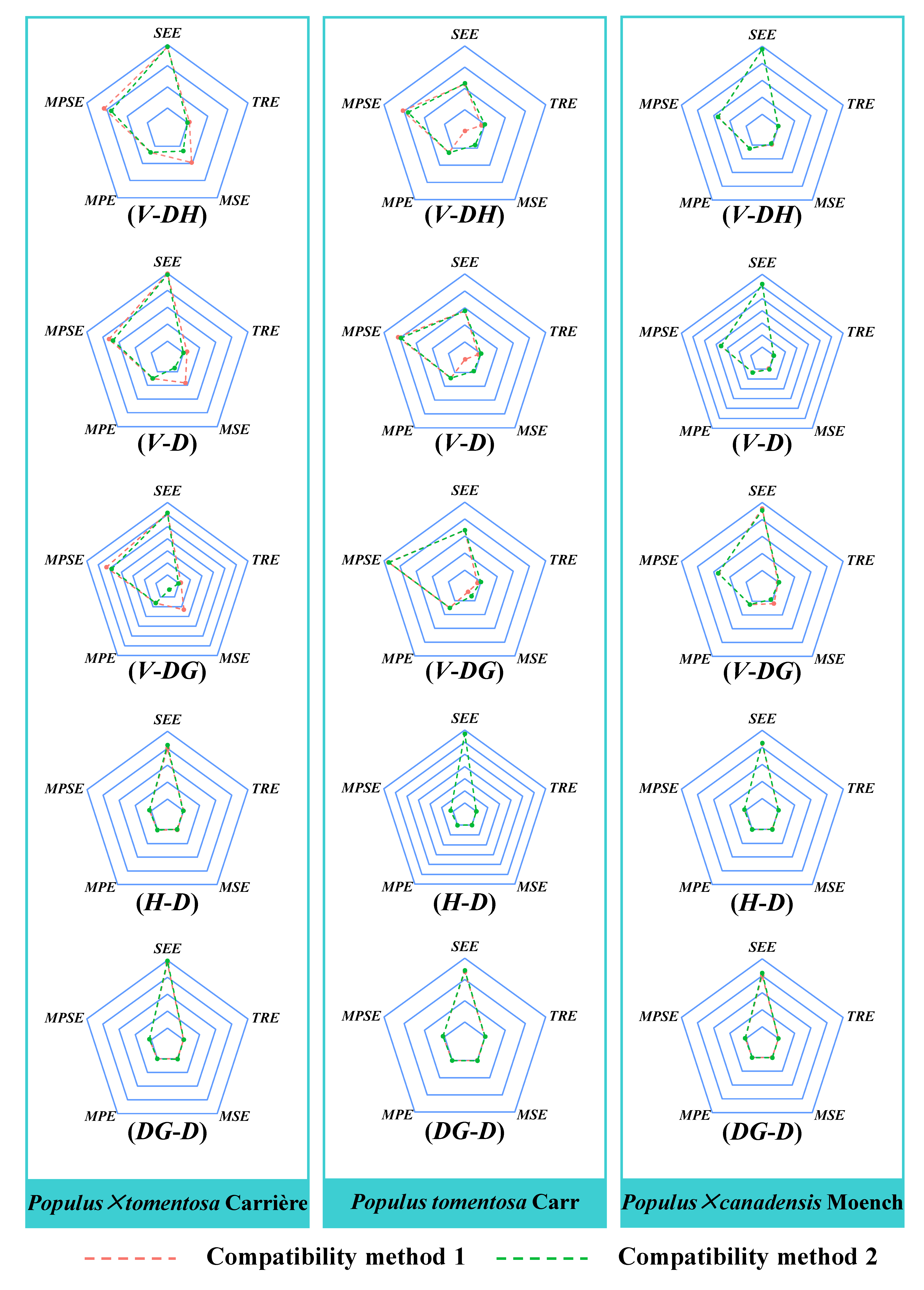
3.3. Analysis of the Two Compatibility Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cervera, M.-T.; Storme, V.; Ivens, B.; Gusmao, J.; Liu, B.H.; Hostyn, V.; Van Slycken, J.; Van Montagu, M.; Boerjan, W. Dense Genetic Linkage Maps of Three Populus Species (Populus deltoides, P. nigra and P. trichocarpa) Based on AFLP and Microsatellite Markers. Genetics 2001, 158, 787–809. [Google Scholar] [CrossRef]
- Liang, W.; Hu, H.; Liu, F.; Zhang, D. Research Advance of Biomass and Carbon Storage of Poplar in China. J. For. Res. 2006, 17, 75–79. [Google Scholar] [CrossRef]
- Subedi, T.; Bhandari, S.K.; Pandey, N.; Timilsina, Y.P.; Mahatara, D. Form Factor and Volume Equations for Individual Trees of Shorea robusta in Western Low Land of Nepal Formfaktor-Und Volumengleichungen Für Den Salbaum (Shorea robusta) Im Westlichen Tiefland von Nepal. Austrian. J. For. Sci. 2021, 138, 143–166. [Google Scholar]
- Oehmcke, S.; Li, L.; Trepekli, K.; Revenga, J.C.; Nord-Larsen, T.; Gieseke, F.; Igel, C. Deep Point Cloud Regression for Above-Ground Forest Biomass Estimation from Airborne LiDAR. Remote Sens. Environ. 2024, 302, 113968. [Google Scholar] [CrossRef]
- Yao, Q.; Wang, J.; Zhang, J.; Xiong, N. Error Analysis of Measuring the Diameter, Tree Height, and Volume of Standing Tree Using Electronic Theodolite. Sustainability 2022, 14, 6950. [Google Scholar] [CrossRef]
- Feng, Z.K.; Sui, H.D.; Deng, X.R.; Zang, S.Y. Survey and precision analysis of tree height by trigonometric leveling. J. Beijing For. Univ. 2007, 29, 31–35. (In Chinese) [Google Scholar]
- Song, S.Y.; Wang, J.; Li, X.L.; Yin, H.L.; Feng, Z.K. Improvement of pressler method for stand volume measurement using electronic theodolite. Bull. Surv. Mapp. 2015, 7, 113–116. (In Chinese) [Google Scholar]
- Basuki, T.M.; Van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric Equations for Estimating the Above-Ground Biomass in Tropical Lowland Dipterocarp Forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Liu, B.; Bu, W.; Zang, R. Improved Allometric Models to Estimate the Aboveground Biomass of Younger Secondary Tropical Forests. Glob. Ecol. Conserv. 2023, 41, e02359. [Google Scholar] [CrossRef]
- Case, B.S.; Hall, R.J. Assessing Prediction Errors of Generalized Tree Biomass and Volume Equations for the Boreal Forest Region of West-Central Canada. Can. J. For. Res. 2008, 38, 878–889. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Hattori, D.; Itioka, T.; Handa, C.; Ohkubo, T.; Kendawang, J.J.; Nakamura, M.; Sakaguchi, M.; Takahashi, N. Development of Allometric Relationships for Accurate Estimation of Above-and below-Ground Biomass in Tropical Secondary Forests in Sarawak, Malaysia. J. Trop. Ecol. 2009, 25, 371–386. [Google Scholar] [CrossRef]
- Diamantopoulou, M.J.; Özçelik, R.; Yavuz, H. Tree-Bark Volume Prediction via Machine Learning: A Case Study Based on Black Alder’s Tree-Bark Production. Comput. Electron. Agr. 2018, 151, 431–440. [Google Scholar] [CrossRef]
- Diamantopoulou, M.J.; Milios, E. Modelling Total Volume of Dominant Pine Trees in Reforestations via Multivariate Analysis and Artificial Neural Network Models. Biosyst. Eng. 2010, 105, 306–315. [Google Scholar] [CrossRef]
- Nieto, P.G.; Torres, J.M.; Fernández, M.A.; Galán, C.O. Support Vector Machines and Neural Networks Used to Evaluate Paper Manufactured Using Eucalyptus Globulus. Appl. Math. Model. 2012, 36, 6137–6145. [Google Scholar] [CrossRef]
- Ogana, F.N.; Ercanli, I. Modelling Height-Diameter Relationships in Complex Tropical Rain Forest Ecosystems Using Deep Learning Algorithm. J. Forestry. Res. 2022, 33, 883–889. [Google Scholar] [CrossRef]
- Sharma, M. Increasing Volumetric Prediction Accuracy—An Essential Prerequisite for End-Product Forecasting in Red Pine. Forests 2020, 11, 1050. [Google Scholar] [CrossRef]
- Joosten, R.; Schumacher, J.; Wirth, C.; Schulte, A. Evaluating Tree Carbon Predictions for Beech (Fagus sylvatica L.) in Western Germany. For. Ecol. Manag. 2004, 189, 87–96. [Google Scholar] [CrossRef]
- Gimenez, B.O.; Dos Santos, L.T.; Gebara, J.; Celes, C.H.; Durgante, F.M.; Lima, A.J.; Dos Santos, J.; Higuchi, N. Tree Climbing Techniques and Volume Equations for Eschweilera (Matá-Matá), a Hyperdominant Genus in the Amazon Forest. Forests 2017, 8, 154. [Google Scholar] [CrossRef]
- Sharma, M. Total and Merchantable Volume Equations for 25 Commercial Tree Species Grown in Canada and the Northeastern United States. Forests 2021, 12, 1270. [Google Scholar] [CrossRef]
- Hogg, B.W.; Lewis, T.; Huth, J.R.; Lee, D.J. Small-Tree Volume Equations for Subtropical Hardwood Plantation Species. Aust. Forestry. 2021, 84, 152–158. [Google Scholar] [CrossRef]
- Tabacchi, G.; Di Cosmo, L.; Gasparini, P. Aboveground Tree Volume and Phytomass Prediction Equations for Forest Species in Italy. Eur. J. Forest. Res. 2011, 130, 911–934. [Google Scholar] [CrossRef]
- Adame, P.; del Río, M.; Canellas, I. A Mixed Nonlinear Height–Diameter Model for Pyrenean Oak (Quercus pyrenaica Willd.). For. Ecol. Manag. 2008, 256, 88–98. [Google Scholar] [CrossRef]
- Tian, X.; Sun, S.; Mola-Yudego, B.; Cao, T. Predicting Individual Tree Growth Using Stand-Level Simulation, Diameter Distribution, and Bayesian Calibration. Ann. Forest. Sci. 2020, 77, 57. [Google Scholar] [CrossRef]
- Chukwu, O.; Osho, J.S. Basal Area-Stump Diameter Models for Tectona grandis Linn. f. Stands in Omo Forest Reserve, Nigeria. J. For. Environ. Sci. 2018, 34, 119–125. [Google Scholar]
- Spurr, S.H. Forest Inventory. In Forest Inventory; Ronald Press Co.: New York, NY, USA, 1952. [Google Scholar]
- Schumacher, F. Logarithmic Expression of Timber-Tree Volume. J. Agric. Res. 1933, 47, 719–734. [Google Scholar]
- Fonweban, J.; Gardiner, B.; Auty, D. Variable-Top Merchantable Volume Equations for Scots Pine (Pinus sylvestris) and Sitka Spruce (Picea sitchensis (Bong.) Carr.) in Northern Britain. Forestry 2012, 85, 237–253. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, D.-L.; Huang, G.-S.; Zeng, W.-S. Developing Aboveground Biomass Equations Both Compatible with Tree Volume Equations and Additive Systems for Single-Trees in Poplar Plantations in Jiangsu Province, China. Forests 2016, 7, 32. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, S. Modeling Compatible Single-Tree Aboveground Biomass Equations for Masson Pine (Pinus massoniana) in Southern China. J. For. Res. 2012, 23, 593–598. [Google Scholar] [CrossRef]
- Zianis, D.; Mencuccini, M. On Simplifying Allometric Analyses of Forest Biomass. For. Ecol. Manag. 2004, 187, 311–332. [Google Scholar] [CrossRef]
- Zeng, W.; Zhang, L.; Chen, X.; Cheng, Z.; Ma, K.; Li, Z. Construction of Compatible and Additive Individual-Tree Biomass Models for Pinus tabulaeformis in China. Can. J. For. Res. 2017, 47, 467–475. [Google Scholar] [CrossRef]
- Boczniewicz, D. Developing Fully Compatible Taper and Volume Equations for All Stem Components of Eucalyptus globoidea Blakely Trees in New Zealand. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2023. [Google Scholar]
- Li, D.; Jia, W.; Guo, H.; Sun, Y.; Wang, F. Compatible Taper and Volume Systems for Larix Olgensis and Larix Kaempferi in Northeast China. Eur. J. Forest. Res. 2023, 143, 65–79. [Google Scholar] [CrossRef]
- Behling, M.; Koehler, H.S.; Behling, A. Compatibility between the Stem Volume and Taper Equations Volume for Black Wattle Trees. Floresta 2020, 50, e1518. [Google Scholar] [CrossRef]
- Bi, H.; Turner, J.; Lambert, M.J. Additive Biomass Equations for Native Eucalypt Forest Trees of Temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Parresol, B.R. Additivity of Nonlinear Biomass Equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Tang, S.; Li, Y.; Wang, Y. Simultaneous Equations, Error-in-Variable Models, and Model Integration in Systems Ecology. Ecol. Model. 2001, 142, 285–294. [Google Scholar] [CrossRef]
- Greene, W.H. Econometric Analysis; Pearson Education India: Noida, India, 2003; ISBN 81-7758-684-X. [Google Scholar]
- Xiong, N.; Qiao, Y.; Ren, H.; Zhang, L.; Chen, R.; Wang, J. Comparison of Parameter Estimation Methods Based on Two Additive Biomass Models with Small Samples. Forests 2023, 14, 1655. [Google Scholar] [CrossRef]
- Fu, L.; Lei, Y.; Wang, G.; Bi, H.; Tang, S.; Song, X. Comparison of Seemingly Unrelated Regressions with Error-in-Variable Models for Developing a System of Nonlinear Additive Biomass Equations. Trees 2016, 30, 839–857. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, D.; Liu, G.; Yang, C.; Teskey, R.O. Additive Tree Biomass Equations for Betula platyphylla Suk. Plantations in Northeast China. Ann. Forest. Sci. 2018, 75, 60. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Henningsen, A.; Hamann, J.D. Systemfit: A Package for Estimating Systems of Simultaneous Equations in R. J. Stat. Softw. 2008, 23, 1–40. [Google Scholar] [CrossRef]
- Fang, Z.; Bailey, R.L. Height–Diameter Models for Tropical Forests on Hainan Island in Southern China. For. Ecol. Manag. 1998, 110, 315–327. [Google Scholar] [CrossRef]
- Curtis, R.O. Height-Diameter and Height-Diameter-Age Equations for Second-Growth Douglas-Fir. Forest. Sci. 1967, 13, 365–375. [Google Scholar]
- Popoola, V.D.; Uii, J.N. Nonlinear Height-Diameter Models for Tectona grandis (Linn. F.) in Mbavaa Forest Reserve, Benue State, Nigeria. J. Res. For. Wildl. Environ. 2023, 15, 20–27. [Google Scholar]
- Shamaki, S.; Akindele, S.; Isah, A.; Mohammed, I. Height-Diameter Relationship Models for Teak (Tectona grandis) Plantation in Nimbia Forest Reserve, Nigeria. Asian J. Environ. Ecol. 2016, 1, 1–7. [Google Scholar] [CrossRef]
- Zang, H.; Lei, X.; Zeng, W. Height–Diameter Equations for Larch Plantations in Northern and Northeastern China: A Comparison of the Mixed-Effects, Quantile Regression and Generalized Additive Models. For. Int. J. For. Res. 2016, 89, 434–445. [Google Scholar] [CrossRef]
- Sharma, M.; Parton, J. Height–Diameter Equations for Boreal Tree Species in Ontario Using a Mixed-Effects Modeling Approach. For. Ecol. Manag. 2007, 249, 187–198. [Google Scholar] [CrossRef]
- Bronisz, K.; Mehtätalo, L. Mixed-Effects Generalized Height–Diameter Model for Young Silver Birch Stands on Post-Agricultural Lands. For. Ecol. Manag. 2020, 460, 117901. [Google Scholar] [CrossRef]
- Mugasha, W.A.; Eid, T.; Bollandsås, O.M.; Malimbwi, R.E.; Chamshama, S.A.O.; Zahabu, E.; Katani, J.Z. Allometric Models for Prediction of Above-and Belowground Biomass of Trees in the Miombo Woodlands of Tanzania. For. Ecol. Manag. 2013, 310, 87–101. [Google Scholar] [CrossRef]
- Fradette, O.; Marty, C.; Tremblay, P.; Lord, D.; Boucher, J.-F. Allometric Equations for Estimating Biomass and Carbon Stocks in Afforested Open Woodlands with Black Spruce and Jack Pine, in the Eastern Canadian Boreal Forest. Forests 2021, 12, 59. [Google Scholar] [CrossRef]
- Shahzad, M.K.; Hussain, A.; Jiang, L. A Model Form for Stem Taper and Volume Estimates of Asian White Birch (Betula platyphylla): A Major Commercial Tree Species of Northeast China. Can. J. For. Res. 2020, 50, 274–286. [Google Scholar] [CrossRef]
- Demaerschalk, J.P. Converting Volume Equations to Compatible Taper Equations. Forest. Sci. 1972, 18, 241–245. [Google Scholar] [CrossRef]
- Zhao, D.; Lynch, T.B.; Westfall, J.; Coulston, J.; Kane, M.; Adams, D.E. Compatibility, Development, and Estimation of Taper and Volume Equation Systems. Forest. Sci. 2019, 65, 1–13. [Google Scholar] [CrossRef]
- Zeng, W.S.; Zhang, H.R.; Tang, S.Z. Using the Dummy Variable Model Approach to Construct Compatible Single-Tree Biomass Equations at Different Scales—A Case Study for Masson Pine (Pinus massoniana) in Southern China. Can. J. For. Res. 2011, 41, 1547–1554. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A Compatible System of Biomass Equations for Three Conifer Species in Northeast, China. For. Ecol. Manag. 2014, 329, 306–317. [Google Scholar] [CrossRef]
- Shao, J. Linear Model Selection by Cross-Validation. J. Am. Stat. Assoc. 1993, 88, 486–494. [Google Scholar] [CrossRef]
- Kozak, A.; Kozak, R. Does Cross Validation Provide Additional Information in the Evaluation of Regression Models? Can. J. For. Res. 2003, 33, 976–987. [Google Scholar] [CrossRef]
- Zeng, W.S.; Tang, S.Z. Goodness evaluation and precision analysis of tree biomass equations. Sci. Silvae Sin. 2011, 47, 106–113. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Characteristics | Minimum | Maximum | Mean | Standard Deviation | Coefficient of Variation |
|---|---|---|---|---|---|---|
| Populus × tomentosa Carrière | D/cm | 6.6 | 69.5 | 25.0 | 13.2 | 52.9% |
| DG/cm | 7.8 | 84.7 | 30.3 | 15.5 | 51.2% | |
| H/m | 9.6 | 36.8 | 21.6 | 6.4 | 29.8% | |
| V/m3 | 0.0156 | 6.4576 | 0.7961 | 1.1344 | 142.5% | |
| Populus tomentosa Carr | D/cm | 5.2 | 49.3 | 21.1 | 9.4 | 44.4% |
| DG/cm | 6.9 | 56.3 | 24.8 | 10.3 | 41.4% | |
| H/m | 9.2 | 36.4 | 20.5 | 5.8 | 28.3% | |
| V/m3 | 0.0114 | 2.0876 | 0.4470 | 0.4480 | 100.2% | |
| Populus × canadensis Moench | D/cm | 8.5 | 73.1 | 39.3 | 12.9 | 32.8% |
| DG/cm | 10.5 | 89.5 | 45.5 | 15.1 | 33.2% | |
| H/m | 10.3 | 35.0 | 24.1 | 4.6 | 19.1% | |
| V/m3 | 0.0385 | 6.1957 | 1.4889 | 1.0187 | 68.4% |
| Tree Species | Model | a0 | a1 | a2 | AIC | R2 |
|---|---|---|---|---|---|---|
| Populus × tomentosa Carrière | Model 1 | 4.724 | 0.4830 | - | 2109.006 | 0.7285 |
| Model 2 | 32.44 | −0.06078 | 1.281 | 2020.619 | 0.7829 | |
| Model 3 | 30.83 | 4.505 | −0.1091 | 2018.225 | 0.7842 | |
| Model 4 | 0.5527 | 0.02131 | - | 2042.76 | 0.7700 | |
| Populus tomentosa Carr | Model 1 | 4.080 | 0.5370 | - | 1976.589 | 0.7590 |
| Model 2 | 192.4 | −7.984 × 10−4 | 0.5415 | 1978.77 | 0.7583 | |
| Model 3 | 36.58 | 2.965 | −0.06409 | 1990.518 | 0.7511 | |
| Model 4 | 0.5223 | 0.02185 | - | 1998.562 | 0.7454 | |
| Populus × canadensis Moench | Model 1 | 6.470 | 0.362 | - | 2104.811 | 0.4720 |
| Model 2 | 28.32 | −0.0601 | 1.214 | 2078.769 | 0.5065 | |
| Model 3 | 27.63 | 3.067 | −0.08843 | 2079.17 | 0.5060 | |
| Model 4 | 0.5270 | 0.02696 | - | 2084.055 | 0.4987 |
| Tree Species | Model | b0 | b1 | AIC | R2 |
|---|---|---|---|---|---|
| Populus × tomentosa Carrière | Model 5 | 1.188 | 1.164 | 1659.307 | 0.9848 |
| Populus tomentosa Carr | Model 5 | 1.825 | 1.087 | 1273.561 | 0.9868 |
| Populus × canadensis Moench | Model 5 | 0.4705 | 1.146 | 2023.708 | 0.9601 |
| Tree Species | Compatible Method | H-D Model | DG-D Model | V-DH Model | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a0 | a1 | a2 | Model | b0 | b1 | Model | c0 | c1 | c2 | Model | ||
| Populus × tomentosa Carrière | (1) | 30.92 | 4.313 | −0.1063 | (3) | 1.166 | 1.165 | (5) | 0.00003218 | 2.029 | 1.009 | (6) |
| (2) | 32.50 | 2.564 | −0.07401 | (3) | 0.1035 | 1.197 | (5) | 0.00004648 | 2.051 | 0.8753 | (6) | |
| Populus tomentosa Carr | (1) | 4.096 | 0.5358 | - | (1) | 1.843 | 1.086 | (5) | 0.00008537 | 1.750 | 0.9779 | (6) |
| (2) | 3.971 | 0.5454 | - | (1) | 1.169 | 1.113 | (5) | 0.00005777 | 1.798 | 1.047 | (6) | |
| Populus × canadensis Moench | (1) | 28.37 | −0.05930 | 1.193 | (2) | 0.4694 | 1.146 | (5) | 0.00005659 | 1.996 | 0.8460 | (6) |
| (2) | 27.54 | −0.08152 | 2.047 | (2) | −2.518 | 1.216 | (5) | 0.00006000 | 1.991 | 0.8341 | (6) | |
| Tree Species | Model | Compatibility Method | R2 | SEE | TRE | MSE | MPE | MPSE |
|---|---|---|---|---|---|---|---|---|
| Populus × tomentosa Carrière | V-DH model | (1) | 0.9840 | 0.1439 | 0.49% | 4.75% | 1.77% | 10.75% |
| (2) | 0.9838 | 0.1448 | −0.03% | 1.37% | 1.78% | 8.97% | ||
| H-D model | (1) | 0.7851 | 2.9979 | −0.03% | −0.13% | 1.36% | 11.00% | |
| (2) | 0.7566 | 3.1909 | −1.52% | −2.57% | 1.45% | 12.06% | ||
| V-D model | (1) | 0.9695 | 0.1996 | 1.08% | 4.10% | 2.46% | 13.15% | |
| (2) | 0.9704 | 0.1966 | −0.02% | −1.37% | 2.42% | 11.89% | ||
| DG-D model | (1) | 0.9848 | 1.9160 | −0.04% | 0.11% | 0.62% | 5.48% | |
| (2) | 0.9837 | 1.9824 | 0.85% | 1.95% | 0.64% | 5.82% | ||
| V-DG model | (1) | 0.9501 | 0.2559 | 0.86% | 6.57% | 3.15% | 21.54% | |
| (2) | 0.9500 | 0.2563 | −0.17% | −3.65% | 3.16% | 19.27% | ||
| Populus tomentosa Carr | V-DH model | (1) | 0.9822 | 0.0599 | −0.87% | −4.98% | 1.31% | 10.32% |
| (2) | 0.9812 | 0.0617 | −0.06% | −0.89% | 1.35% | 9.04% | ||
| H-D model | (1) | 0.7596 | 2.8486 | −0.01% | 0.01% | 1.36% | 10.91% | |
| (2) | 0.7595 | 2.8495 | 0.15% | 0.29% | 1.36% | 10.94% | ||
| V-D model | (1) | 0.9590 | 0.0912 | −0.77% | −4.84% | 2.00% | 15.63% | |
| (2) | 0.9595 | 0.0908 | 0.00% | −0.53% | 1.99% | 14.76% | ||
| DG-D model | (1) | 0.9868 | 1.1830 | −0.01% | −0.06% | 0.47% | 3.71% | |
| (2) | 0.9861 | 1.2127 | 0.41% | 1.00% | 0.48% | 3.94% | ||
| V-DG model | (1) | 0.9328 | 0.1172 | −1.08% | −3.33% | 2.57% | 18.63% | |
| (2) | 0.9326 | 0.1174 | −0.06% | −1.69% | 2.57% | 18.46% | ||
| Populus × canadensis Moench | V-DH model | (1) | 0.9650 | 0.1914 | −0.01% | −0.23% | 1.26% | 8.64% |
| (2) | 0.9650 | 0.1914 | −0.07% | −0.50% | 1.26% | 8.70% | ||
| H-D model | (1) | 0.5090 | 3.2326 | −0.01% | 0.00% | 1.31% | 10.98% | |
| (2) | 0.4998 | 3.2627 | 0.45% | 0.89% | 1.33% | 11.33% | ||
| V-D model | (1) | 0.9360 | 0.2597 | −0.13% | −0.30% | 1.71% | 12.88% | |
| (2) | 0.9359 | 0.2599 | −0.01% | 0.12% | 1.71% | 12.79% | ||
| DG-D model | (1) | 0.9602 | 3.0214 | −0.01% | 0.05% | 0.65% | 4.54% | |
| (2) | 0.9563 | 3.1634 | 0.48% | 1.67% | 0.68% | 5.58% | ||
| V-DG model | (1) | 0.8746 | 0.3645 | −0.22% | 1.66% | 2.40% | 16.98% | |
| (2) | 0.8817 | 0.3539 | 0.37% | −1.31% | 2.33% | 17.24% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, Z.; Feng, Z.; Yu, Z.; Li, J. Construction of Compatible Volume Model for Populus in Beijing, China. Forests 2024, 15, 1059. https://doi.org/10.3390/f15061059
Wang S, Wang Z, Feng Z, Yu Z, Li J. Construction of Compatible Volume Model for Populus in Beijing, China. Forests. 2024; 15(6):1059. https://doi.org/10.3390/f15061059
Chicago/Turabian StyleWang, Shan, Zhichao Wang, Zhongke Feng, Zhuang Yu, and Jinshan Li. 2024. "Construction of Compatible Volume Model for Populus in Beijing, China" Forests 15, no. 6: 1059. https://doi.org/10.3390/f15061059