

Effects of Stand Density on Tree Growth, Diversity of Understory Vegetation, and Soil Properties in a Pinus koraiensis Plantation

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Vegetation Data Collection
2.3. Soil Sample Collection
2.4. Statistical Analyses
3. Results
3.1. Effects of Stand Density on Tree Growth
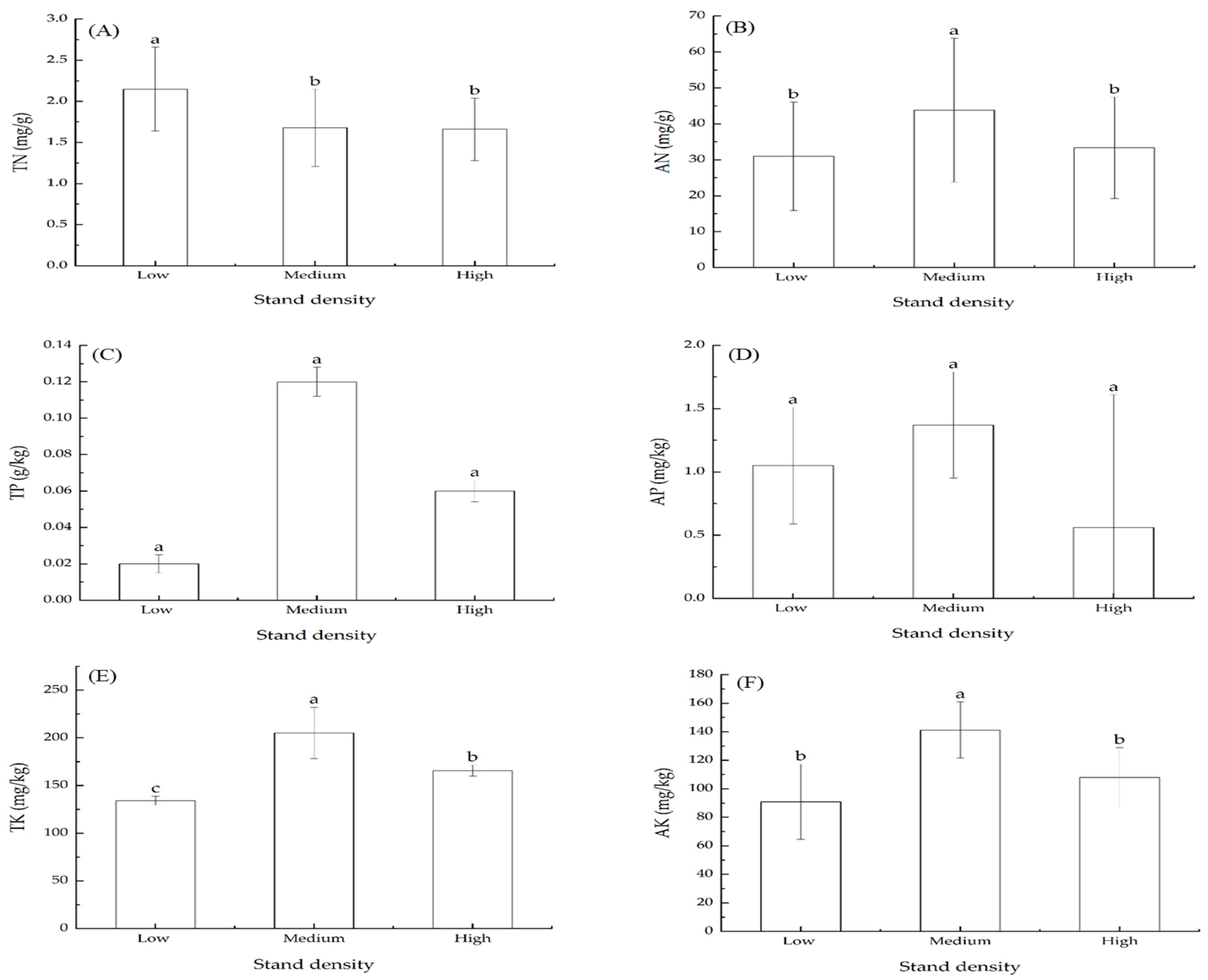
3.2. Effects of Stand Density on Total Nitrogen (TN) and Available Nitrogen (AN)
3.3. Effects of Stand Density on Total Phosphorous (TP) and Available Phosphorous (AP)
3.4. Effects of Stand Density on Total Potassium (TK) and Available Potassium (AK)
3.5. Effect of Stand Density on Physical Soil Properties (pH, SMC, BD, and SOM)
3.6. Correlation Analysis of Selected Soil Chemical and Physical Properties
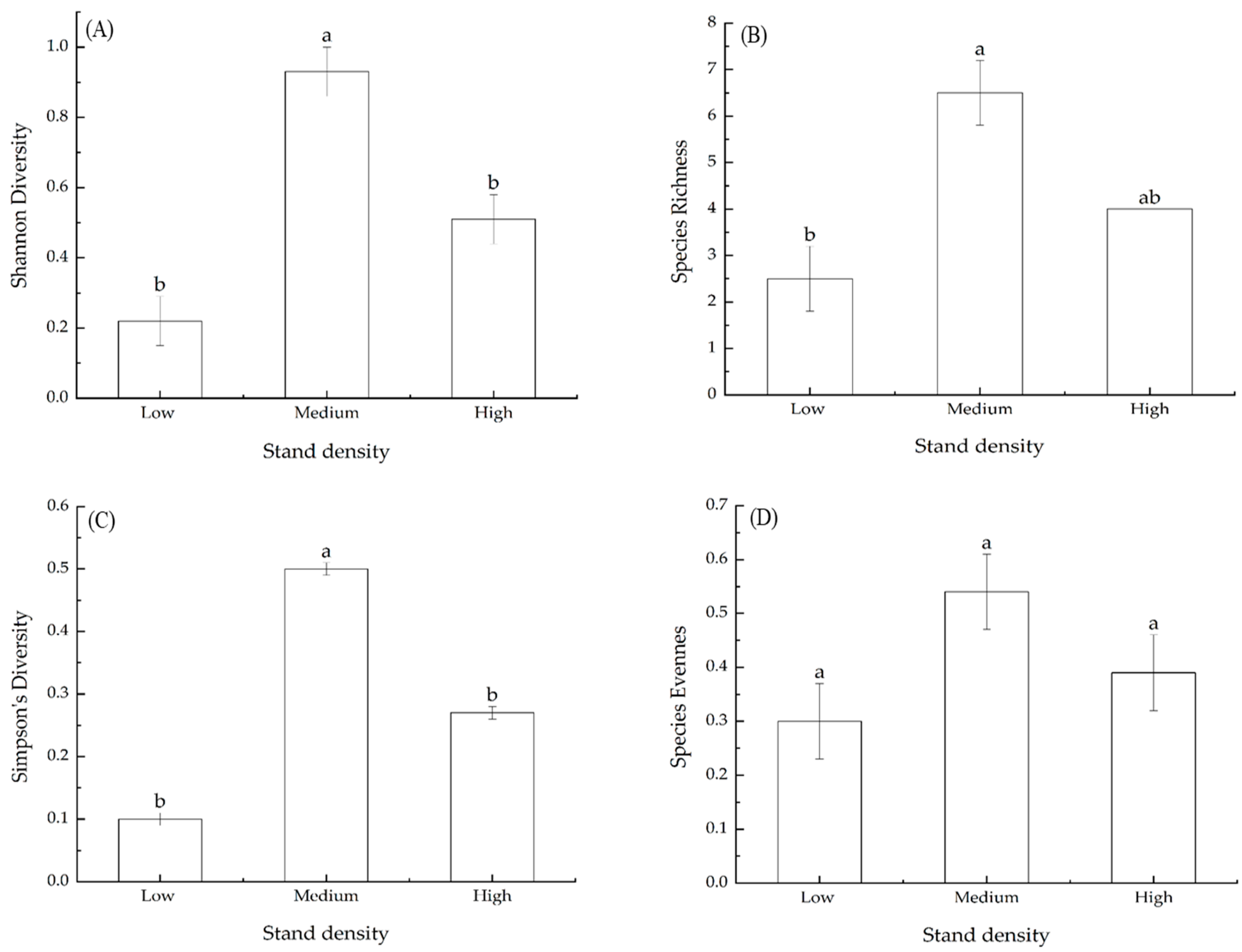
3.7. Effects of Stand Density on Species Diversity Indices of Shrubs
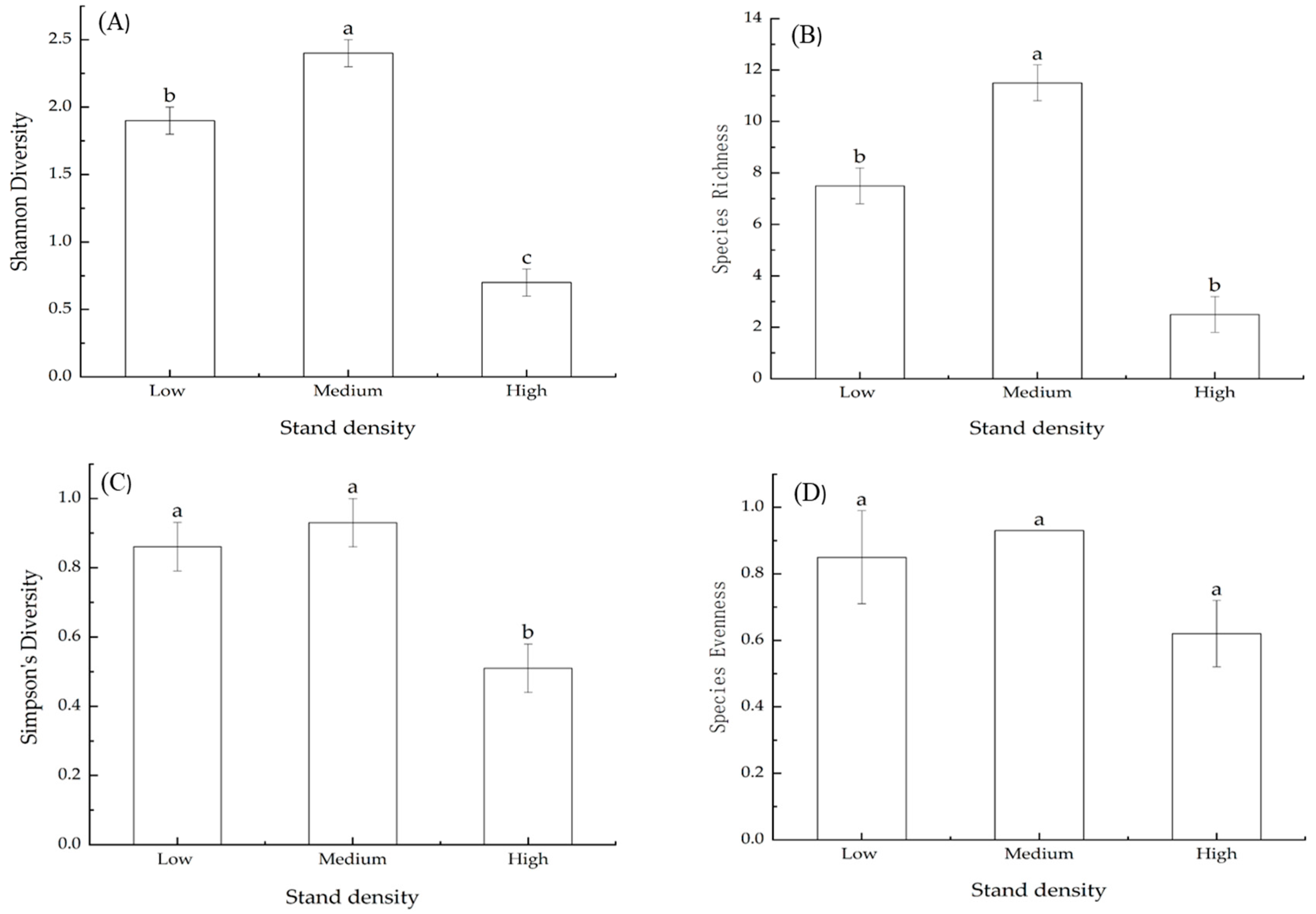
3.8. Effects of Stand Density on Species Diversity Indices of Herbs
3.9. Species Important Value across Various Stand Density Types
4. Discussion
4.1. Effect of Stand Density on Tree Growth
4.2. Effect of Stand Density on Soil Chemical Properties
4.3. Effect of Stand Density on Physical Soil Properties (pH, SMC, BD, and SOM)
4.4. Effect of Stand Density on Understory Biodiversity
4.5. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burkhart, H.E. Comparison of maximum size–density relationships based on alternate stand attributes for predicting tree numbers and stand growth. For. Ecol. Manag. 2013, 289, 404–408. [Google Scholar] [CrossRef]
- Liechty, H.O.; Mroz, G.D.; Reed, D.D. The growth and yield responses of a high site quality red pine plantation to seven thinning treatments and two thinning intervals. Can. J. For. Res. 1986, 16, 513–520. [Google Scholar] [CrossRef]
- Baker, F. Stand Density and Growth. 1952. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19536602531 (accessed on 31 May 2024).
- Qiu, X.; Peng, D.; Wang, H.; Wang, Z.; Cheng, S. Minimum data set for evaluation of stand density effects on soil quality in Larix principis-rupprechtii plantations in North China. Ecol. Indic. 2019, 103, 236–247. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 2014, 73, 238–245. [Google Scholar] [CrossRef]
- Lei, J.; Du, H.; Duan, A.; Zhang, J. Effect of stand density and soil layer on soil nutrients of a 37-year-old Cunninghamia lanceolata plantation in Naxi, Sichuan Province, China. Sustainability 2019, 11, 5410. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Cañellas, I.; Calama, R.; Gordo, J.; Sánchez-González, M. Thinning increases cone production of stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ann. For. Sci. 2013, 70, 761–768. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, D.; Li, W.; Sun, D.; Jin, C.; Yuan, F.; Wang, A.; Wu, J. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. For. Ecol. Manag. 2018, 429, 36–43. [Google Scholar] [CrossRef]
- Fu, X.; Wang, J.; Di, Y.; Wang, H. Differences in fine-root biomass of trees and understory vegetation among stand types in subtropical forests. PLoS ONE 2015, 10, e0128894. [Google Scholar] [CrossRef]
- Liu, K.; He, H.; Xu, W.; Du, H.; Zong, S.; Huang, C.; Wu, M.; Tan, X.; Cong, Y. Responses of korean pine to proactive managements under climate change. Forests 2020, 11, 263. [Google Scholar] [CrossRef]
- Choi, J.; Lee, B.; Lee, D.; Choi, I. Growth monitoring of Korean white pine (Pinus koraiensis) plantation by thinning intensity. J. Korean Soc. For. Sci. 2014, 103, 422–430. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Guo, J.; Tan, Z.; Dong, W.; Wang, H. Changes in the understory diversity of secondary Pinus tabulaeformis forests are the result of stand density and soil properties. Glob. Ecol. Conserv. 2021, 28, e01628. [Google Scholar] [CrossRef]
- Ruano, I.; Rodríguez-García, E.; Bravo, F. Effects of pre-commercial thinning on growth and reproduction in post-fire regeneration of Pinus halepensis Mill. Ann. For. Sci. 2013, 70, 357–366. [Google Scholar] [CrossRef]
- Ali, A.; Dai, D.; Akhtar, K.; Teng, M.; Yan, Z.; Urbina-Cardona, N.; Mullerova, J.; Zhou, Z. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 2019, 20, e00775. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pihlainen, S.; Niinimäki, S. On the economics of optimal timber production in boreal Scots pine stands. Can. J. For. Res. 2013, 43, 719–730. [Google Scholar] [CrossRef]
- Schoenholtz, S.H.; Van Miegroet, H.; Burger, J. A review of chemical and physical properties as indicators of forest soil quality: Challenges and opportunities. For. Ecol. Manag. 2000, 138, 335–356. [Google Scholar] [CrossRef]
- Schmitt, A.; Trouvé, R.; Seynave, I.; Lebourgeois, F. Decreasing stand density favors resistance, resilience, and recovery of Quercus petraea trees to a severe drought, particularly on dry sites. Ann. For. Sci. 2020, 77, 1–21. [Google Scholar]
- Blevins, D.P.; Prescott, C.E.; Allen, H.L.; Newsome, T.A. The effects of nutrition and density on growth, foliage biomass, and growth efficiency of high-density fire-origin lodgepole pine in central British Columbia. Can. J. For. Res. 2005, 35, 2851–2859. [Google Scholar] [CrossRef]
- Lal, R.; Shukla, M.K. Principles of Soil Physics; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Von Lützow, M.; Leifeld, J.; Kainz, M.; Kögel-Knabner, I.; Munch, J. Indications for soil organic matter quality in soils under different management. Geoderma 2002, 105, 243–258. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Ares, A.; Neill, A.R.; Puettmann, K.J. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. For. Ecol. Manag. 2010, 260, 1104–1113. [Google Scholar] [CrossRef]
- Rota, E.; Caruso, T.; Bargagli, R. Community structure, diversity and spatial organization of enchytraeids in Mediterranean urban holm oak stands. Eur. J. Soil Biol. 2014, 62, 83–91. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Bond, B.J.; Ritchie, G.A.; Scott, W. Effects of stand density on the growth of young Douglas-fir trees. Can. J. For. Res. 2002, 32, 420–427. [Google Scholar] [CrossRef]
- Noh, N.J.; Son, Y.; Lee, S.K.; Yoon, T.K.; Seo, K.W.; Kim, C.; Lee, W.-K.; Bae, S.W.; Hwang, J. Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. J. Plant Res. 2010, 123, 411–419. [Google Scholar] [CrossRef]
- Xin, S.; Wang, J.; Mahardika, S.B.; Jiang, L. Sensitivity of Stand-Level Biomass to Climate for Three Conifer Plantations in Northeast China. Forests 2022, 13, 2022. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Tai, D.T.; Zhang, P.; Razaq, M.; Shen, H.-L. Effect of thinning intensity on tree growth and temporal variation of seed and cone production in a Pinus koraiensis plantation. J. For. Res. 2019, 30, 835–845. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, G.; Yang, J.; Li, S.; Li, R.; Zou, W. Effects of planting densities on production and dynamics of litter of Mytilaria laosensis plantation. Bull. Bot. Res. 2017, 37, 768–777. [Google Scholar]
- Chen, B.; Li, Y.; Fan, S.; Peng, C.; Huang, B.; Liu, G. Soil properties and understory species diversity at different stand densities in a tropical rainforest on Hainan Island, China. Austrian J. For. Sci. 2020, 137, 225–246. [Google Scholar]
- Wang, C.; Yang, Q.; Chen, J.; Zhang, C.; Liu, K. Variations in Soil Organic Carbon Fractions and Microbial Community in Rice Fields under an Integrated Cropping System. Agronomy 2023, 14, 81. [Google Scholar] [CrossRef]
- Navarro, F.B.; Jimenez, M.N.; Canadas, E.M.; Gallego, E.; Terron, L.; Ripoll, M.A. Effects of different intensities of overstory thinning on tree growth and understory plant-species productivity in a semi-arid Pinus halepensis Mill. afforestation. For. Syst. 2010, 19, 410–417. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, H.; Wen, Y.; Goodale, U.M.; Li, X.; You, Y.; Ye, D.; Liang, H. Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. For. Ecol. Manag. 2018, 410, 164–173. [Google Scholar] [CrossRef]
- Krajnc, L.; Farrelly, N.; Harte, A.M. The influence of crown and stem characteristics on timber quality in softwoods. For. Ecol. Manag. 2019, 435, 8–17. [Google Scholar] [CrossRef]
- Farooq, T.; Ma, X.; Rashid, M.; Wu, W.; Xu, J.; Tarin, M.; He, Z.; Wu, P. Impact of stand density on soil quality in Chinese fir (Cunninghamia lanceolata) monoculture. Appl. Ecol. Environ. Res. 2019, 17, 553–3566. [Google Scholar] [CrossRef]
- Watt, M.S.; Davis, M.R.; Clinton, P.W.; Coker, G.; Ross, C.; Dando, J.; Parfitt, R.L.; Simcock, R. Identification of key soil indicators influencing plantation productivity and sustainability across a national trial series in New Zealand. For. Ecol. Manag. 2008, 256, 180–190. [Google Scholar] [CrossRef]
- Ross, C.W.; Watt, M.S.; Parfitt, R.L.; Simcock, R.; Dando, J.; Coker, G.; Clinton, P.W.; Davis, M.R. Soil quality relationships with tree growth in exotic forests in New Zealand. For. Ecol. Manag. 2009, 258, 2326–2334. [Google Scholar] [CrossRef]
- Jhariya, M.; Singh, L. Productivity and carbon accumulation in tropical dry deciduous forest ecosystem. In Tropical Dry Forests: Emerging Features and Ecological Perspectives; Chaturvedi, R.K., Singh, R., Bhadouria, R., Eds.; Nova Science: Hauppauge, NY, USA, 2021; pp. 269–287. [Google Scholar]
- Zhao, M.; Liu, S.; Sun, Y.; Chen, Y. Does stand density affect understory vegetation and soil properties of differently aged Robinia pseudoacacia plantations? For. Ecol. Manag. 2023, 548, 121444. [Google Scholar] [CrossRef]
- Vesterdal, L.; Dalsgaard, M.; Felby, C.; Raulund-Rasmussen, K.; Jørgensen, B.B. Effects of thinning and soil properties on accumulation of carbon, nitrogen and phosphorus in the forest floor of Norway spruce stands. For. Ecol. Manag. 1995, 77, 1–10. [Google Scholar] [CrossRef]
- Zarafshar, M.; Rousta, M.J.; Matinizadeh, M.; Talebi, K.S.; Bordbar, S.K.; Alizadeh, T.; Nouri, E.; Bader, M.K.-F. Scattered wild pistachio trees profoundly modify soil quality in semi-arid woodlands. Catena 2023, 224, 106983. [Google Scholar] [CrossRef]
- Powers, M.D.; Pregitzer, K.S.; Palik, B.J.; Webster, C.R. Wood δ13C, δ18O and radial growth responses of residual red pine to variable retention harvesting. Tree Physiol. 2010, 30, 326–334. [Google Scholar] [CrossRef]
- Bae, S.-W.; Hwang, J.-H.; Lee, S.-T.; Kim, H.-S.; Jeong, J.-M. Changes in soil temperature, moisture content, light availability and diameter growth after thinning in Korean pine (Pinus koraiensis) plantation. J. Korean Soc. For. Sci. 2010, 99, 397–403. [Google Scholar]
- Cheng, J.; Ding, C.; Li, X.; Zhang, T.; Wang, X. Soil quality evaluation for navel orange production systems in central subtropical China. Soil Tillage Res. 2016, 155, 225–232. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, J.; Zeng, S.; Wu, D.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J. Soils Sediments 2017, 17, 2230–2238. [Google Scholar] [CrossRef]
- Fang, X.-M.; Zhang, X.-L.; Zong, Y.-Y.; Zhang, Y.; Wan, S.-Z.; Bu, W.-S.; Chen, F.-S. Soil phosphorus functional fractions and tree tissue nutrient concentrations influenced by stand density in subtropical Chinese fir plantation forests. PLoS ONE 2017, 12, e0186905. [Google Scholar] [CrossRef]
- Chen, L.; Wang, D.; Yu, F.; Wang, Z. Effects of stand density on physical and chemical properties of soil moisture. J. Northeast. For. Univ. 2013, 41, 61–64. [Google Scholar]
- López, R.; Gondar, D.; Iglesias, A.; Fiol, S.; Antelo, J.; Arce, F. Acid properties of fulvic and humic acids isolated from two acid forest soils under different vegetation cover and soil depth. Eur. J. Soil Sci. 2008, 59, 892–899. [Google Scholar] [CrossRef]
- Matinizadeh, M.; Korori, S.; Teimouri, M.; Praznik, W. Enzyme Activities in Undisturbed and Disturbed Forest Soils under Oak (Quercus brantii var. persica) as Affected by Soil Depth and Seasonal Variation. 2008. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/20083189779 (accessed on 31 May 2024).
- Achat, D.L.; Augusto, L.; Bakker, M.R.; Gallet-Budynek, A.; Morel, C. Microbial processes controlling P availability in forest spodosols as affected by soil depth and soil properties. Soil Biol. Biochem. 2012, 44, 39–48. [Google Scholar] [CrossRef]
- Wei, L.; Archaux, F.; Hulin, F.; Bilger, I.; Gosselin, F. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests. For. Ecol. Manag. 2020, 460, 117897. [Google Scholar] [CrossRef]
- Hao, J.F.; Wang, D.Y.; Li, Y.; Zhu, Y.H.; Yao, X.L.; Qi, J.Q. Effect of altitude on structure and species diversity of Cunninghamia lanceolata plantation in Jiangyou district, Sichuan Province. Acta Bot. Boreal.-Occident. Sin. 2014, 34, 2544–2552. [Google Scholar] [CrossRef]
- Wang, S.S.; Yu, X.X.; Luo, Y.Y.; Zhao, G.L.; Yang, J.; Hai, L. Effects of reforestation on woody plant biodiversity of forest vegetation in Badaling, Beijing. J. Beijing For. Univ. 2008, 30 (Suppl. 2), 155–159. [Google Scholar]
- Thysell, D.R. Effects of Forest Management on Understory and Overstory Vegetation: A Retrospective Study; Pacific Northwest Research Station: Portland, OR, USA, 2000; Volume 488. [Google Scholar]
- Ding, J.; Zhang, Y.; Guo, Y.; Yang, S.; Ren, D.; He, Z.; Guo, J. Influences of the Density on Understory Species Composition and Diversity of Larix principis-rupprechtii Natural Forest. J. Northwest For. Univ. 2018, 4, 10–16. [Google Scholar]
- Alem, S.; Pavlis, J.; Urban, J.; Kucera, J. Pure and mixed plantations of Eucalyptus camaldulensis and Cupressus lusitanica: Their growth interactions and effect on diversity and density of undergrowth woody plants in relation to light. Open J. For. 2015, 5, 375. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Ritchie, M.W.; Neal, D.L. Overstory and understory dynamics in a ponderosa pine plantation vary with stand density in the Sierra Nevada: 40-year results. For. Sci. 2013, 59, 670–680. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecol. Monogr. 2000, 70, 171–207. [Google Scholar] [CrossRef]
- Schenk, H.J. Root competition: Beyond resource depletion. J. Ecol. 2006, 94, 725–739. [Google Scholar] [CrossRef]
- Fang, J.; Wang, X.; Shen, Z.; Tang, Z.; He, J.; Yu, D.; Jiang, Y.; Wang, Z.; Zheng, C.; Zhu, J. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Density Level | Stand Density/Plot | No. of Trees/Plot | Mean Diameter (cm)/Plot | Mean Height (m)/Plot | Mean Volume (m3)/Plot |
|---|---|---|---|---|---|
| Low | 708.33 ± 11.78 | 42.50 ± 0.71 | 22.86 ± 4.28 | 19.63 ± 2.41 | 0.45 ± 0.18 |
| Medium | 841.66 ± 11.78 | 50.50 ± 0.71 | 24.58 ± 0.09 | 20.91 ± 0.06 | 0.52 ± 0.00 |
| High | 900.00 ± 23.57 | 54.00 ± 0.71 | 23.49 ±0.83 | 21.10 ± 0.11 | 0.49 ± 0.04 |
| No. | Soil Properties | Method/Equipment |
|---|---|---|
| 1 | pH | pH meter |
| 2 | Bulk density (BD) | The core method of the Nanjing Institute of Soil Science (1978) |
| 3 | Moisture content (SMC %) | Calculated based on wet and dry weight |
| 4 | Soil organic matter (SOM) | Wet oxidation method (CN elemental analyzer) and then calculated as SOM |
| 5 | Total nitrogen | CN elemental analyzer |
| 6 | Total phosphorus | AA3 flow analyzer |
| 7 | Total potassium | Flame photometry |
| 8 | Available nitrogen | Diffusion method |
| 9 | Available phosphorus | AA3 flow analyzer |
| 10 | Available potassium | Flame photometry |
| Density Level | Soil Properties | |||||
|---|---|---|---|---|---|---|
| First Layer (0–10 cm) | TN (mg g−1) | TP (g/kg) | TK (mg/kg) | AN (mg g−1) | AP (mg/kg) | AK (mg/kg) |
| LSD | 2.78 ± 0.01 a | 0.15 ± 0.02 a | 134.23 ± 0.90 c | 46.13 ± 2.07 b | 1.01 ± 0.34 a | 117.04 ± 18.03 b |
| MSD | 2.26± 0.04 b | 0.17 ± 0.09 a | 223.48 ± 32.23 a | 61.06 ± 0.15 a | 0.87 ± 0.01 a | 159.80 ± 12.18 a |
| HSD | 2.11 ± 0.16 b | 0.10 ± 0.03 a | 166.26 ± 0.54 b | 43.28 ± 4.13 b | 0.56 ± 0.21 a | 125.69 ± 23.14 b |
| Second layer (10–20 cm) | ||||||
| LSD | 2.02 ± 0.12 a | 0.04 ± 0.12 a | 133.41 ± 9.57 a | 25.83 ± 12.12 a | 1.20 ± 0.66 a | 79.50 ± 27.39 a |
| MSD | 1.48 ± 0.11 a | 0.06 ± 0.08 a | 200.80 ± 31.14 a | 39.17 ± 16.46 a | 1.60 ± 0.21 a | 127.04 ± 22.78 a |
| HSD | 1.53 ± 0.17 a | 0.02 ± 0.06 a | 163.05 ± 3.48 a | 36.57 ± 1.13 a | 0.48 ± 1.43 a | 93.71 ± 13.10 a |
| Third layer (20–30 cm) | ||||||
| LSD | 1.66 ± 0.01 a | 0.05 ± 0.13 a | 134.86 ± 3.22 a | 20.91 ± 16.63 a | 0.93 ± 0.65 a | 76.27 ± 18.83 a |
| MSD | 1.29 ± 0.09 b | 0.12 ± 0.13 a | 191.44 ± 22.42 a | 31.17 ± 27.82 a | 1.63 ± 0.31 a | 136.85 ± 11.84 a |
| HSD | 1.33 ± 0.09 b | 0.06 ± 0.09 a | 167.77 ± 11.25 a | 20.16 ± 20.30 a | 0.62 ± 1.84 a | 104.39 ± 20.3 a |
| Density Level | Soil Properties | |||
|---|---|---|---|---|
| First Layer (0–10 cm) | pH | SMC (%) | BD (g/cm3) | OM |
| LSD | 5.85 ± 0.30 a | 0.14 ± 0.03 a | 0.37 ± 0.01 a | 52.30 ± 3.34 a |
| MSD | 6.24 ± 0.03 a | 0.09 ± 0.01 a | 0.36 ± 0.00 a | 48.00 ± 3.13 a |
| HSD | 5.94 ± 0.34 a | 0.11 ± 0.02 a | 0.37 ± 0.01 a | 48.09 ± 0.85 a |
| Second layer (10–20 cm) | ||||
| LSD | 5.57 ± 0.04 a | 0.11 ±0.01 a | 0.36 ± 0.00 a | 30.61 ± 6.36 a |
| MSD | 5.80 ± 0.03 a | 0.09 ± 0.00 ab | 0.36 ± 0.01 a | 28.54 ± 1.41 a |
| HSD | 5.51 ± 0.44 a | 0.06 ± 0.02 b | 0.37 ± 0.01 a | 29.42 ± 6.51 a |
| Third layer (20–30 cm) | ||||
| LSD | 5.42 ± 0.16 a | 0.10 ± 0.01 a | 0.36 ± 0.01 a | 21.90 ± 3.15 a |
| MSD | 5.59 ± 0.33 a | 0.08 ± 0.01 a | 0.37 ± 0.00 a | 21.00 ± 1.47 a |
| HSD | 5.50 ± 0.44 a | 0.07 ± 0.02 a | 0.36 ± 0.02 a | 19.71 ± 7.75 a |
| Index | TN | TP | TK | AN | AP | AK | ADBH | AH | TV |
|---|---|---|---|---|---|---|---|---|---|
| TN | 1 | 0.25 | −0.23 | 0.44 | −0.15 | 0.15 | −0.46 | −0.62 | −0.33 |
| TP | 1 | 0.46 | 0.42 | 0.28 | 0.73 ** | −0.25 | −0.59 | −0.52 | |
| TK | 1 | 0.43 | 0.08 | 0.80 ** | 0.36 | 0.07 | −0.44 | ||
| AN | 1 | −0.22 | 0.67 ** | −0.11 | −0.47 | −0.43 | |||
| AP | 1 | 0.1 | −0.15 | −0.21 | 0.17 | ||||
| AK | 1 | −0.41 | −0.73 | −0.72 | |||||
| ADBH | 1 | 0.91 * | 0.43 | ||||||
| AH | 1 | 0.65 | |||||||
| TV | 1 |
| Density Level | Species | RD (%) | RF (%) | RC (%) | IV |
|---|---|---|---|---|---|
| (a) | |||||
| Low | Lespedeza bicolor Turcz. | 95.83 | 63.64 | 81.13 | 80.20 |
| Aralia chinensis L. | 4.17 | 36.36 | 18.87 | 19.80 | |
| Medium | Lespedeza bicolor Turcz. | 71.6 | 30.77 | 55.04 | 52.50 |
| Prunus padus L. | 1.23 | 7.69 | 0.44 | 3.10 | |
| Corylus heterophylla Fisch. | 2.47 | 15.38 | 4.38 | 7.40 | |
| Sambucus williamsii Hance | 1.23 | 15.38 | 3.65 | 6.80 | |
| Vitis amurensis Rupr. | 3.7 | 15.38 | 7.3 | 8.80 | |
| Rubus crataegifolius Bge. | 19.75 | 15.38 | 29.2 | 21.40 | |
| High | Lespedeza bicolor Turcz. | 87.93 | 63.64 | 70.30 | 74.00 |
| Sorbaria sorbifolia (L.) A. Braun | 1.72 | 9.09 | 3.96 | 4.90 | |
| Sambucus williamsii Hance | 8.62 | 18.18 | 21.78 | 16.20 | |
| Corylus heterophylla Fisch. | 1.72 | 9.09 | 3.96 | 4.90 | |
| (b) | |||||
| Low | Festuca rubra L. | 18.2 | 17.6 | 1.15 | 12.3 |
| Potentilla cryptotaeniae Maxim. | 9.1 | 8.6 | 0.03 | 5.9 | |
| Athyrium brevifrons Nakai ex Kitagawa | 27.3 | 26.1 | 93.31 | 48.9 | |
| Sedum aizoon L. | 18.2 | 19.2 | 1.69 | 13.0 | |
| Lamium barbatum Sieb. et Zucc. | 9.1 | 9.0 | 1.12 | 6.4 | |
| Clematis florida Thunb. | 9.1 | 9.4 | 0.45 | 6.3 | |
| Geum aleppicum Jacq. | 9.1 | 10.2 | 2.25 | 7.2 | |
| Medium | Clematis terniflora DC. | 7.14 | 8.61 | 1.67 | 5.81 |
| Linnaea borealis L. | 7.14 | 7.50 | 0.03 | 4.89 | |
| Thalictrum aquilegiifolium var. sibiricum Linnaeus | 14.29 | 14.44 | 0.74 | 9.82 | |
| Deyeuxia pyramidalis | 7.14 | 8.61 | 1 | 5.58 | |
| Agrimonia pilosa Ldb. | 14.29 | 14.17 | 2.51 | 10.32 | |
| Vicia unijuga A. Br. | 7.14 | 8.89 | 0.2 | 5.41 | |
| Lunaria annua L. | 7.14 | 5.83 | 1.67 | 4.88 | |
| Impatiens noli-tangere L. | 7.14 | 6.39 | 0.13 | 4.55 | |
| Crepidiastrum denticulatum (Houtt.) Pak & Kawano | 7.14 | 6.67 | 0.13 | 4.65 | |
| Lamium barbatum Sieb. et Zucc. | 14.29 | 12.78 | 8.36 | 11.81 | |
| Athyrium brevifrons Nakai ex Kitagawa | 7.14 | 6.11 | 83.56 | 32.27 | |
| High | Festuca rubra L. | 75 | 55.47 | 56.25 | 62.2 |
| Thalictrum aquilegiifolium var. sibiricum Linnaeus | 12.5 | 41.20 | 12.5 | 22.1 | |
| Vicia unijuga A. Br. | 12.5 | 3.33 | 31.25 | 15.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iddrisu, A.-Q.; Hao, Y.; Issifu, H.; Getnet, A.; Sakib, N.; Yang, X.; Abdallah, M.M.; Zhang, P. Effects of Stand Density on Tree Growth, Diversity of Understory Vegetation, and Soil Properties in a Pinus koraiensis Plantation. Forests 2024, 15, 1149. https://doi.org/10.3390/f15071149
Iddrisu A-Q, Hao Y, Issifu H, Getnet A, Sakib N, Yang X, Abdallah MM, Zhang P. Effects of Stand Density on Tree Growth, Diversity of Understory Vegetation, and Soil Properties in a Pinus koraiensis Plantation. Forests. 2024; 15(7):1149. https://doi.org/10.3390/f15071149
Chicago/Turabian StyleIddrisu, Abdul-Qadir, Yuanqin Hao, Hamza Issifu, Ambachew Getnet, Nazmus Sakib, Xiubo Yang, Mutaz Mohammed Abdallah, and Peng Zhang. 2024. "Effects of Stand Density on Tree Growth, Diversity of Understory Vegetation, and Soil Properties in a Pinus koraiensis Plantation" Forests 15, no. 7: 1149. https://doi.org/10.3390/f15071149
APA StyleIddrisu, A.-Q., Hao, Y., Issifu, H., Getnet, A., Sakib, N., Yang, X., Abdallah, M. M., & Zhang, P. (2024). Effects of Stand Density on Tree Growth, Diversity of Understory Vegetation, and Soil Properties in a Pinus koraiensis Plantation. Forests, 15(7), 1149. https://doi.org/10.3390/f15071149