Abstract
Nitrogen (N) is an essential mineral element for plants and is the main component of protein, nucleic acid, phospholipid, chlorophyll, hormones, vitamins and alkaloids. It is involved in all stages of plant growth and development. Low-N stress seriously hinders plant growth and reduces yield and quality. Plants have evolved a series of elaborate regulatory mechanisms for N uptake and assimilation to cope with different soil N states. After N is absorbed and utilized by plants, it plays important regulatory roles in phytohormones, microRNA (miRNA), root development and mycorrhizal symbiosis to cope with environmental stress. Here, we highlight the research progress on the regulation mechanisms for N absorption and assimilation. Then, we emphasize the regulation roles of N regarding hormone signals, miRNA, lateral root growth, drought resistance, anthocyanin synthesis and mycorrhizal symbiosis. A thorough understanding of N uptake, utilization and interaction with other biological processes is helpful to improve N use efficiency and to breed plants with “less-input-more-output”.
1. Introduction
As an essential macronutrient, the content of nitrogen (N) is second only to carbon and oxygen in the dry weight of plants. N is the primary constituent of proteins, nucleic acids, phospholipids, chlorophyll, hormones, vitamins and alkaloids, and participates in all stages of plant growth and development [1,2,3]. In natural ecosystems and agricultural production systems, N deficiency is often one of the important limiting factors, because soil erosion seriously aggravates soil poverty, resulting in a reduction in soil N. Forest ecosystems may contain some fast-growing tree species, which require large amounts of N. When N absorbed by plants cannot meet their normal growth and development, low-N stress occurs. In response to low-N stress, plants will degrade their own proteins to achieve N reuse. However, this often results in decreased photosynthesis, accelerated leaf senescence and a slowing down of biomass accumulation, which then causes irreversible damage to the plants [4]. A large amount of N fertilizer is used in order to increase yield and only 30%–50% of the applied fertilizer N is actually captured by plants [5], resulting in low N use efficiency (NUE). The extensive application of N fertilizer not only increases the cost of crop production but also causes serious environmental problems such as the eutrophication of surface water sources, nitrate pollution of drinking water and soil acidification. Therefore, determining how to improve plant NUE has become a critical point of focus that must be solved for the sustainable development of current agriculture practices.
Nitrogen has an important effect on the uptake of water from soil by plants. A plant’s acquisition of N sources (mainly nitrate and ammonium N) is generally a function of root uptake. The effect of inorganic N sources (nitrate and ammonium N) on water uptake varies with plant type. N and water exert both synergistic and antagonistic effects on plant growth [6,7]. The effect of N fertilizer application on plant growth under drought stress is influenced by the plant species, N fertilizer absorption preferences, drought stress severity, N fertilizer application levels and the treatment time. The N content in plants is also associated with the ability of the antioxidant system to remove reactive oxygen species (ROS). Shi et al. [8] reported an enhancement in antioxidant enzyme activities, including superoxide dismutase (SOD) activity and glutathione (GSH) content, under normal N supply conditions compared with a low-N treatment, effectively inhibiting ROS accumulation and consequently reducing drought stress damage to plants. N can also induce the expression of hormone metabolism genes in root systems, suggesting its ability to act as a signaling molecule via hormone metabolism [9].
The role of N in plants is twofold; namely, it provides a large amount of essential N element for plants and serves as a signal substance that participates in a wide range of plant developmental processes. This review focuses on the absorption, assimilation and regulation of two inorganic N sources in plants, as well as the interaction between N and drought resistance, lateral root growth, hormone signals, anthocyanin synthesis, mycorrhizal symbiosis and microRNA (miRNA) regulation. The information presented here on the regulation of the N metabolism can be usefully adopted in research on the regulation of N uptake and utilization in plants, and it provides theoretical knowledge for the efficient application of N fertilizers to crops to cope with biotic and abiotic stresses caused by low-N stress.
2. Study of the Mechanism of N Uptake and Assimilation in Plants
Plants have evolved different absorption systems for inorganic N, including nitrate transporters and ammonium transporters, with extremely complex regulatory mechanisms in order to adapt to the varying types and concentrations of N in the external environment.
2.1. Research Progress on NO3− Uptake in Plants
Nitrate transporters in plants can be classified into the nitrate transporter 1/peptide transporter family (NRT1/PTR), nitrate transporter 2 family (NRT2), chloride channel (CLC) and the slow anion channel-associated 1/slow anion channel homologues (SLAC1/SLAH) based on the structure, location and function of the transporter [10,11].
Plant nitrate transporters (NRTs) play key roles in NO3−–N uptake by roots, as well as long-distance transport and redistribution in roots and stems [7,12]. NO3−–N transport systems in plants can be divided into two types, including low-affinity transport systems (LATS, >1 mM) and high-affinity transport systems (HATS, 1 μm–1 mM). In addition, each transport system can be categorized as cLATS/cHATS and iLATS/iHATS, depending on whether gene expression is induced by the NO3−–N concentration in the soil [1,13]. Plant NRT genes are mainly divided into three families, namely, NRT1/PTR, NRT2 and NRT3. Since NRT1 and PTR are closely related in terms of the gene structure and function, the nitrate transporter 1/peptide transporter family (NRT1/PTR Family) is denoted as NPF [14]. A total of 53 NRT1 and seven NRT2 members in Arabidopsis thaliana (hereafter, Arabidopsis) have been identified. NAR2 is the accessory/partner protein of NRT2, which forms a complex with NRT2 then maintains NRT2 protein stability [11]. NRT1 not only transports nitrate N but also transports a variety of substrate molecules, indicating that the NRT1 protein plays an important role in all aspects of plant growth and development [11]. Among them, some NRT1 family transporters and all NRT2 family transporters exhibit the capacity to transport NO3−–N (Figure 1). The first identified NRT member in Arabidopsis was AtNRT1.1 or CHL1 [15], which is a double-affinity transporter and possesses transport capacity for both high and low NO3−–N concentrations [16].
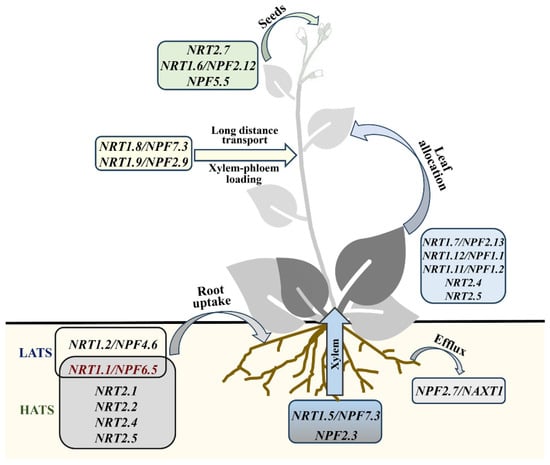
Figure 1.
Role of NPF and NRT2 families in nitrogen uptake and utilization of Arabidopsis. HATS, high-affinity transport system; LATS, low-affinity transport system.
CLC transporters are a class of chloride and other anion channel proteins localized in organelles or cytoplasmic membranes. Such proteins generally include 10–12 transmembrane domains and C-terminal and N-terminal regions located in the cytoplasm. There are generally three highly conserved amino acid sequences related to anion-specific recognition among different transmembrane domains [17]. In Arabidopsis, the CLC transporter family includes seven members, denoted as a–g [18]. CLCa is located on the vacuolar membrane and can store nitrate in the vacuole of plants while transporting chloride ions in the reverse direction [19,20]. Recent studies showed that nucleotides and phospholipids can regulate the CLCa activity of Arabidopsis in different physiological scenarios [18]. CLCb is also localized in the vacuolar membrane. Research on Xenopus oocytes has demonstrated that CLCb also has the function of Cl−/NO3− reverse cotransport [21]. Shi et al. [22] showed that under low-N conditions, HRS1 HOMOLOG2 (HHO2) expression was down-regulated and negatively regulated CLCb, resulting in the up-regulated expression of CLCb, which promoted vacuolar NO3− efflux, improved NUE and thus improved plant adaptability to low N.
SLAC1/SLAH slow anion channel transporters are localized in the plasma membrane of plant guard cells and are closely related to ABA-regulated stomatal closure. Arabidopsis has five SLAC1/SLAH family members, namely SLAC1 and SLAH1–SLAH4, all of which contain 10 helical domains [23]. SLAH2 is expressed in root column cells and may mediate nitrate transport from the root to the shoot [24]. Following changes in the endogenous ABA signal induced by environmental changes, SLAC1 and SLAH3 have been reported to exhibit transport activity that regulates the efflux process of chloride ions and nitrate, which subsequently leads to stomatal closure [25]. SLAC1 shows a similar transport capacity for chloride and nitrate, while a stronger transport capacity has been observed for SLAH3 [26]. Recent studies have revealed that NRT1.1 and SLAH3 can form a functional unit to regulate the nitrate-dependent alleviation of ammonium toxicity in Arabidopsis [27].
2.2. Research Progress on NH4+ Uptake in Plants
Ammonium transporters (AMTs) are responsible for the uptake of NH4+-N from soil [28]. The first ammonia transporter, denoted as AtAMT1.1, was isolated by Ninnemann in 1994 [29]. The AMT gene family in plants consists of two subfamilies, AMT1 and AMT2. The number of members in each subfamily varies with the plant species. For example, in Arabidopsis, the AMT1 and AMT2 families include five and two members, respectively [30]. The AMT1 family of poplar includes six members, while the AMT2 family includes eight members. The AMT1 and AMT2 families of rice include three and seven members, respectively [31]. The protein sequences of the AMT1 and AMT2 members are distinct, yet the high-level structures of the proteins are similar and contain 9–11 transmembrane structural regions, while the tertiary structures are also very similar [32]. The expression of AMT family members varies depending on the plant tissues and organs. This may be a result of their different functions [33]. In Arabidopsis, AMT1.1 and AMT1.3 are localized on the cell membrane of the root epidermis and cortex and occupy approximately 30% of the high-affinity absorption system, respectively [34]. Overexpression of OsAMT1.1 can significantly improve crop NH4+-N uptake and yield under low NH4+-N conditions. Moreover, the NO3−-inducible promoter region on 5′UTR of OsNAR2.1 can promote the expression of OsAMT1.1 and increase the grain yield of pOsNAR2.1:OsAMT1.1 transgenic plants compared with the wild type and pUbi:OsAMT1.1 transgenic plants [35]. AMT1.2 is localized in the root cortex and is mainly responsible for the transportation of ammonium ions to the vascular system or the absorption of ammonium ions from non-protoplasts of roots. AMT1.4 is specifically expressed in plant flowers, and its primary function is transporting ammonium ions to pollen to maintain pollen activity [36]. AMT2.1 is expressed in numerous plant tissues, including the root tips, cortex and vascular bundles, yet its absorption capacity for ammonium ions in the soil is weak [37].
2.3. Nitrogen Assimilation in Plants
The assimilation process of NO3− typically includes the following steps: (i) the absorption of NO3− and reduction to nitrite; (ii) reduction to NH4+; (iii) NH4+ synthesis to glutamine (Gln) under the action of glutamine synthetase (GS); (iv) the glutamate synthetase (GOGAT) catalytic formation of glutamate (Glu); and (v) conversion to the synthesis of other amino acids used by plants (Figure 2). Nitrate reductase (NR), which catalyzes the reduction of NO3−–N to NO2−–N in plants, is a rate-limiting enzyme induced by NO3−–N concentrations [38,39]. NR also has a sensitive response to abiotic stress [40]. Nitrite reductase (NiR) is mainly found in plant chloroplasts and catalyzes the reduction of NO2− to NH4+. The activity of NiR is affected by the supply of NO2−. Reduced NH4+ is catalyzed by GS and subsequently combines with Glu to form Gln by consuming ATP. The NH4+ utilized in the above process mainly comes from two sources: direct root absorption and NO3− reduction. GS is generally observed in plants in two forms, namely GS1 in the cytoplasm and GS2 in the plastid [41,42]. In plants, glutamate dehydrogenase (GDH) regulates the synthesis and decomposition of Glu by catalyzing the reversible reactions of amination/deamination [10]. Masclaux-Daubresse et al. [43] revealed that GDH is mainly responsible for the deamination of Glu, while the GS/GOGAT pathway acts as the dominant synthesis pathway of Glu. Many genes are involved in the regulation of enzyme activity related to N metabolism, such as NR, NiR and GS [44].
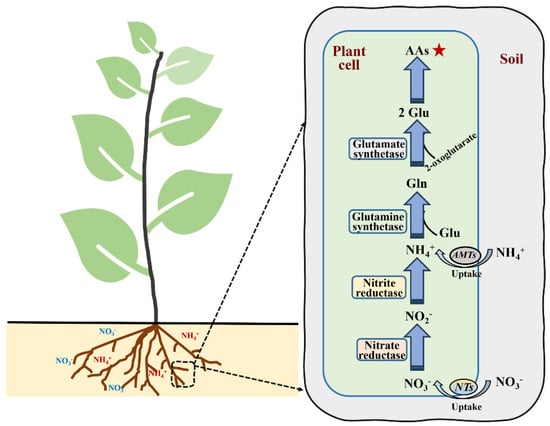
Figure 2.
The process of nitrogen assimilation in plants. Gln: glutamine; Glu: glutamate; AAs: amino acids; NTs: nitrate transporters; AMTs: Ammonium transporters. The star that the products of nitrogen assimilation are amino acids (AAs).
3. Research Progress on Plants’ N Utilization and Regulation
The N transporters, assimilation enzymes and signal transduction in plants are regulated by transcriptional regulation in response to changes in available N concentrations. Posttranscriptional modification and calcium- and phosphorylation-related signaling cascades are all critical posttranscriptional regulatory pathways [45]. The metabolic pathways of carbon metabolism and hormones also undergo corresponding changes to adapt to the regulation of N metabolism and the plant growth process [46]. The double-affinity nitrate transporter NRT1.1 in Arabidopsis is a signal transduction protein with a nitrate sensing function. An NRT1.1-induced nitrate response in plants is required for the normal expression of nitrate-sensitive genes in plants [47]. NRT1.1 shows dual-affinity nitrate transport ability, which is due to the phosphorylation and dephosphorylation of Thr101, and nitrate signals promote this change. When Thr101 phosphorylates, NRT1.1 shows high-affinity nitrate transport ability; when Thr101 dephosphorylates, it shows low-affinity nitrate transport ability [48]. Previous studies have demonstrated that NRT1.1 may not be the only nitrate receptor, and NRT2.1 could potentially be an additional nitrate sensor mediating root growth, yet its role in nitrate signaling remains unclear [49].
Numerous transcription factors (TFs) related to N metabolism have been identified in plants [50,51,52,53]. In particular, the nitrate signaling pathway and the role of calcium ions as downstream second messengers of nitrate signaling in plant roots and stems are well understood. Recent research has identified NIN-like protein 7 (NLP7) as a transcription factor of the N signaling pathway and also as a nitrate receptor [12]. In addition to NLP7, other TFs in plants that regulate the expression of genes related to N metabolism include Arabidopsis Regulated 1 (ANR1), LOB Domain-Containing proteins (LBD37/38/39), Squamosa Promoter Binding Protein-Like 9 (SPL9), Basic Leucine-Zipper 1 (bZIP1), NAC Domain-Containing Protein 4 (NAC4), TGA1/TGA4, NLP2, Teosinte Branched1/Cycloidea/Proliferating Cell Factor 20 (TCP20), CIPK23 and Antares Regulatory Gene 2 (NRG2) [46,54,55]. BES1, a brassinosteroid TF, is strongly induced and bound to the promoters of NRT2.1 and NRT2.2 directly, which then promote their expression under nitrate-deficient conditions [56]. NRT1.1B-NLP3-mediated NO3−–N signal transduction can regulate the utilization of ammonium, thus endowing rice with the ability to reduce ammonium toxicity and adapt to high-ammonium conditions [57]. In addition, ABI2, a member of the protein phosphatase 2C family, and CBL1, a calcium sensor, also exert regulatory effects on NRT1.1 nitrate transport [58]. CIPK8 is specifically involved in nitrate response under high-N conditions and acts as a positive regulator, while CIPK23 is specifically involved in nitrate response under low-N conditions through the direct phosphorylation of NRT1.1Thr101 [59]. eIF4E1 is upstream of NRT1.1 in the nitrate signal transduction pathway and can regulate nitrate signal transduction by regulating NRT1.1 at the translation and transcription levels. The overexpression of eIF4E1 can significantly promote plant growth and increase yield and N use efficiency [60].
4. Regulation Roles of Nitrogen in Plants
NUE is an important agronomic indicator. Generally, NUE can be defined as the crop yield per unit of N uptake from soil. Another evaluation method of NUE is the usage index, which is expressed as the ratio of the aboveground biomass to the aboveground N concentration and usually used to indicate the NUE at the vegetative growth stage [61]. Plant N status can affect many biological processes that then change the NUE of plants.
4.1. Nitrogen and Hormone Signals
Ethylene acts as an important cellular signal to mediate a range of adaptive responses to external N status, and the biosynthesis and signaling of ethylene are regulated by N availability and N forms [62]. The N-triggered ethylene signal can regulate a number of morphological and physiological responses in plants, including the root system architecture (RSA), N uptake and translocation, ammonium toxicity, anthocyanin accumulation and premature senescence [63]. Moreover, ethylene also plays an important role in nodulation formation and N fixation in legumes [64]. Chen et al. [2] showed that the expression of 1-aminocylopropane-1-carboxylic acid (ACC) synthase gene ACS2/6/7/11 and ACC oxidase gene ACO2 in the ethylene biosynthetic pathway were up-regulated by ammonium as the sole N source (5 mM NH4+) and down-regulated by nitrate as the sole N source (5 mM NO3−). Plants can typically produce ethylene under both low- and high-N conditions. When plants are transferred from high-N to low-N conditions (10 mM to 0.2 mM NO3− media), the expression of EIN3 is induced for the rapid production of ethylene [65]. When plants are transferred from low- to high-N conditions (0.1 mM to 10 mM NO3− media), the expression of ACS2/4/5/6/7/8/11 and ACO1/2 is up-regulated to produce a surge in ethylene emission [66]. Previous work has revealed that ethylene production induced by excessive N is often accompanied by negative effects such as cell death, leaf necrosis and growth inhibition, while ethylene production induced by N deficiency generally plays a positive role in plant adaptive response [67]. Comparing the natural variation in ethylene production in Arabidopsis under nitrate-deficient conditions indicates that regulating the ethylene pathway can improve NUE [68].
Auxin (IAA) is an important plant endogenous hormone and plays a key role in the growth and development of plants [69]. Ma et al. [70] found that environmental N availability directly regulates the expression of TAA1 and its homologs TAR1 and TAR2, which are fundamental in the first step of IAA synthesis. The authors also determined that high concentrations of auxin can inhibit the growth of primary roots and promote the formation of lateral roots. This is of great significance for the response of plants to low-N stress. This positive regulatory mechanism plays an important role in cucumber resistance to low-N environments. Krouk et al. [71] proved that nitrate can regulate auxin transport by means of NRT1.1, as the NRT1.1 and AUX/LAX proteins from different transporter families (PTR and ATF1, respectively) share common amino acid transporters. MdATG5a can positively regulate N uptake under low-N conditions by increasing the accumulation of IAA in apple roots, suggesting an important link between N uptake and IAA [4]. In addition, Chen et al. [72] proved the close relationship between the auxin receptor gene OsTIR1 and the uptake and utilization of N. This indicates that auxin plays an important role in N uptake and utilization. The F-box protein 3 (AFB3), which belongs to a group of F-box receptors for IAA, was found to be the only IAA receptor gene rapidly induced by NO3−, and specifically in the main root tip and pericycle. This suggests that NO3− not only modulates IAA gradients in roots, through NRT1.1 dual NO3−/IAA transport activity, but also increases IAA sensitivity of the roots through stimulation of AFB3 expression [1]. The aforementioned findings from previous studies demonstrate the interaction between N and auxin, yet the specific regulatory mechanism requires further research.
Abscisic acid (ABA), also known as a plant stress hormone, plays an important role in plant growth and can inhibit lateral root development [73]. High nitrate concentrations can promote an increase in available ABA content in Arabidopsis by stimulating the conversion of abiotically active ABA–glucose ester (ABA-GE) to biologically active ABA [74]. The low affinity nitrate transporter NRT1.2/NPF4.6/AIT1 mediates ABA uptake in Arabidopsis seeds during germination and post-germination growth. Thus, the N signal can relieve the inhibitory effect of ABA on seed germination by reducing ABA levels [75]. NLP8 in nitrate signaling has also been observed to participate in the regulation of the ABA metabolism by directly binding to the promoter of CYP707A2, a key enzyme gene for ABA decomposition [76]. Su et al. [77] found that SnRK2s, key protein kinases in ABA signaling, play an important role in root growth and nitrate uptake in Arabidopsis under low-N stress. Under N starvation stress, SnRK2s in plants phosphorylate NRT1.1 to regulate nitrate transport activity and mediate plant tolerance to low-N stress. This reveals a new mechanism of ABA signaling for the balance of stress adaptation and growth in plants under low-N stress, and it expands the regulatory network of plants responding to low-N stress. Furthermore, the resupply of nitrate after N starvation in wheat can induce ABA accumulation in roots through activating the ANR1/BG1 pathway, and as a consequence, ABA can up-regulate NRT2/NAR2 gene expression through Wabi5 [78]. These results suggest that ABA can also regulate nitrate uptake and signal transduction through transcriptional regulation and post-translational modification under low-N stress.
Cytokinins (CTKs) act as important signaling molecules in long-distance communication between organs in systemic growth regulation in response to N conditions. N-dependent isopentenyl transferase (IPT) expression accumulates N6-(Δ2-isopentenyl) adenine (iP)- and trans-zeatin (tZ)-type CTK species in the roots and shoots, then activates CTK signaling [79]. Previous research on Arabidopsis has shown that the expression of IPT3, a key enzyme in the first step of CTK synthesis, is induced by N. An NRT1.1-mediated primary N signal induces a strong expression of IPT3 in roots and a weak expression in stems. This indicates the close relationship between nitrate and CTK synthesis in plants, particularly in roots, through the activation of IPT3 [80]. Interestingly, the transcription factor TCP20 has been reported to participate in nitrate–cytokinin–auxin signaling. Guan et al. [81] revealed that lateral root growth was inhibited under a heterogeneous nitrate concentration environment and promoted under low-N conditions in a root split experiment, highlighting the root foraging deficiency of this phenotype. However, TCP20 inhibits the CTK metabolism-related gene ARR4/6/7 and auxin AUX/IAA13, 16 and 27, which consequently reduces auxin signaling [82]. These results suggest that TCP20 may act as an intermediate node in the nitrate–cytokinin–auxin signaling network.
4.2. Nitrogen Metabolism and miRNA Regulation
miRNA plays an important regulatory role in numerous plant growth stages and can regulate plant response to low-N stress [83,84]. The sequencing of two wheat small RNA libraries proved that many target genes were predicted to be involved in the process of N metabolism [85]. Studies have shown that N deficiency has an impact on the elongation of the primary root and the growth of the lateral root of miR393-overexpressing plants. Furthermore, miR393 can be induced by nitrate to regulate the auxin receptor gene in Arabidopsis roots [86]. Yu et al. [87] reported that Osa-MIR169o overexpression will contribute to increased NUE. ARF6 and ARF8, the target genes of miR167, regulate the lateral root development of plants. Thus, the down-regulation of miR167 and the up-regulation of ARF8 will induce lateral root development [88]. However, IAR3, another target gene of miR167, can hydrolyze indole-3-acetic acid-alanine to auxin, and miR167 can affect plant growth by regulating IAR3 [89]. The up-regulation of miR167 expression has been reported under low-N conditions, and changes in miR167 precursor genes vary with tissue type [90]. The expression levels of miR169 and miR398 can increase under low-N conditions, which may regulate the expression of the target gene ARF8 and control the lateral root structure, while miR169 can also shear HAP2-1, a gene that regulates nodule primordium differentiation. This shows that miR169 is significantly down-regulated in Arabidopsis under low-nitrate stress, while its target gene NFYA family is significantly up-regulated [91]. Cheng et al. [92] also concluded that the miR169c–NFYA10 module confers tolerance to low-N stress in Betula luminifera. The overexpression of miR444 can increase NO3− accumulation and increase NRTs gene expression under high-N conditions, but it reduces the ability of plants to transport NO3− from old leaves to new leaves and ultimately affects the plant’s ability to adapt to N deficiency [93]. Other research demonstrated the applicability of miRNA171a as a simple sequence repeat (SSR) candidate marker to discriminate contrasting genotypes for NUE in wheat [94].
4.3. Nitrogen Regulates Lateral Root Growth
The imbalanced distribution of nutrient elements in the soil, particularly essential elements such as N and phosphorus, affects the morphogenesis of plant roots [95,96,97]. Research on Arabidopsis has shown that plants can sense and respond to different concentrations of nitrate and subsequently regulate the density and length of their lateral roots through their interaction with various hormones, thus enabling more efficient usage of nitrate N [98,99]. The Arabidopsis ABF2 and ABF3 TFs can directly regulate most of the nitrate response genes in the root endodermis cells. At the same time, the growth and development of lateral roots in response to nitrate could be regulated by the ABA signal, which establishes a relationship between the nitrate signal, ABA pathway and lateral root development [100]. In addition, ammonium transporters play an important role in the synergistic regulation of rice crown root formation as well as the response to ammonium N [101]. Under low N conditions, NRT1.1 promotes coreceptor QSK1 phosphorylation, transmits signals to PM ATPase AHA2, controls the activation of its phosphorylation site and inhibits the phosphorylation rate. This subsequently leads to changes in proton pump activity and the regulation of lateral root growth [102].
Nitrate can affect root development by regulating the polar transport and signaling of auxin [103]. Krouk et al. [71] revealed that NRT1.1 not only transports nitrate but is also involved in the uptake of auxin. In particular, NRT1.1 represses lateral root growth during times of low nitrate availability by promoting the transport of basipetal auxin out of these roots. The phosphorylation of the AtNRT1.1 protein is involved in the regulation of lateral root development and pattern-triggered immunity (PTI) response in Arabidopsis under low nitrate concentrations. At low nitrate concentrations, the AtNRT1.1 protein promotes the basal transport of auxin and reduces the accumulation of auxin in lateral roots, thus inhibiting the growth of lateral roots. Under high nitrate concentrations, auxin transport mediated by the AtNRT1.1 protein is inhibited, and the accumulation of auxin in the lateral roots is enhanced, which promotes the generation of lateral root primordia and the growth of new lateral roots [104]. Bouguyon et al. [105] demonstrated that high concentrations of nitrate down-regulated the expression level of AtNRT1.1 proteins in lateral roots through post-transcriptional levels. Therefore, the AtNRT1.1 protein has a low content in the lateral roots and exhibits a poor basipetal auxin transport ability under high nitrate concentrations. Together, these two effects promote the germination and growth of the lateral roots. In addition to auxin, cytokinin, ethylene, abscisic acid and other phytohormones are also involved in the nitrate-regulated lateral root growth process [106].
In addition to its role in co-regulating lateral root development with auxin, the AtNRT1.1 protein also regulates lateral root growth through its downstream CpKS–NLP7–ANR1 (Arabidopsis nitrate regulated 1) signal pathway to adapt to different nitrate supply conditions in the outside world. The MADS-box transcription factor ANR1 is the first identified key factor in the nitrate regulation of lateral root development. The overexpression of ANR1 can up-regulate the number and length of lateral roots of Arabidopsis across different nitrate concentrations. Under high nitrate concentrations, the down-regulation of ANR1 expression results in a phenotype with arrested lateral root development [107], while under sufficient N, nitrate stimulates the expression of ANR1 in the lateral root primordial and other sites in an AtNRT1.1-dependent manner, which consequently induces the occurrence of lateral roots [108]. Analysis has revealed the ability of AtNLP7 to bind to the promoter of the ANR1 gene and regulate its expression [109]. When nitrate is deficient, the SENSITIVE-TO-PROTON-RHIZOTOXICITY1 (STOP1) transcription factor accumulates in the nucleus of lateral root primordia cells and directly regulates the expression of NRT1.1 in Arabidopsis (Figure 3). Zhang et al. [106] also found that the phosphorylation status of the AtNRT1.1 protein T101 can control lateral root development by regulating its endocytosis and signal transduction capacity, thereby affecting auxin transport and ANR1 gene expression.
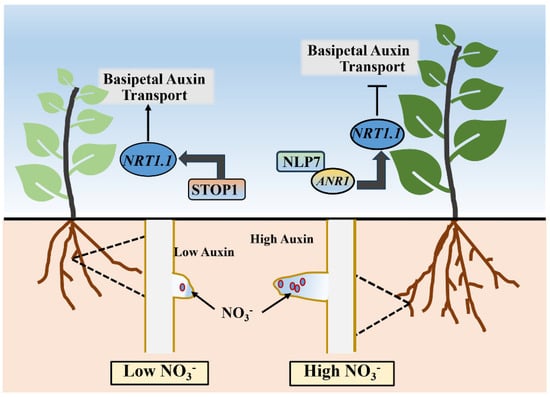
Figure 3.
The NRT1.1-mediated nitrate-dependent lateral root development pathway in plants.
4.4. Nitrogen and Drought Resistance
The effect of the N source on water uptake by plant roots varies with the type of source [110]. Guo et al. [111] reported that, when subjected to drought stress, the photosynthetic rate of rice with NH4+-N as the only N source decreases, and the root growth is less inhibited. Previous research has revealed that plants’ water use efficiency showed an increase with the application of N fertilizer, which contributes to ROS scavenging and cell membrane maintenance, consequently increasing water absorption [112]. Furthermore, the application of N fertilizer can alleviate nutrient stress, maintain plant physiological activities, promote biomass accumulation and avoid damage caused by both water and nutrient stress [113]. The sufficient application of N fertilizer is able to alleviate the inhibitory effect of drought on root growth, particularly for diameters < 0.2 mm, thus promoting plant resistance to drought stress [8]. Drought stress enhances plants’ N allocation process in leaves, while N application reduces this trend [6]. Moderate nitrate supply can alleviate apple drought stress by maintaining the photosynthetic capacity of seedlings [114]. Recent studies have shown that calcium-dependent protein kinase 6 (CPK6) regulates Arabidopsis drought stress resistance by altering the phosphorylation of nitrate transporter AtNRT1.1 under high N conditions [7]. The overexpression of a DREB-like transcription factor, GmTDN1, from soybean (Glycine max L. Merr.) can enhance the resistant ability of two winter wheat (Triticum aestivum L.) varieties, Shi4185 and Jimai22, to drought and low-N stress by improving photosynthesis, osmotic regulation, ROS scavenging, drought stress-related genes and nitrate transporter NRT2.5 expression [115].
Drought stress significantly inhibits plant growth and development mainly because it affects plant N metabolism and the NO3−–N signaling [110,116]. A recent study proved that TFs involved in ABA signaling could affect N metabolism and NO3−–N responses, and, meanwhile, NO3−–N signaling might affect the expression of ABA and drought resistance genes [117]. The opening process of plant stomata also requires the participation and regulation of N, and the content of ABA in leaves will decrease under high-N conditions, which is conducive to the opening of the plant stomata. Shi et al. [8] demonstrated that under drought stress, an appropriate N element supply could increase the ABA content in the leaves of Catalpa bungei and improve drought tolerance. In plants, the target-of-rapamycin (TOR) kinase plays an important role in N signals and drought stress, which can phosphorylate the ABA receptors PYL. When drought stress occurs, TOR was inhibited by ABA-activated SnRK2s phosphorylate to deal with adversity. Meanwhile, N signals can trigger the small GTPase Rho-related protein 2 (ROP2) in plants, which will activate TOR. Thus, the ROP2–TOR module may perceive and conduct N signals to promote plant growth [118,119]. ABA directly suppresses NO3−–N signal perception and transport [117]. NO3−–N is sensed and transported by NRT1.1 (a transporter and sensor, also known as CHL1 or NPF6.3) to plant root cells [59], while ABA signaling can oversee the activity of NRT1.1 through the phosphatase ABI2. During this regulatory process, ABI2 dephosphorylates the CBL1–CIPK23 complex, which regulates phosphorylation and subsequently inhibits NRT1.1 [58] (Figure 4). ABF2 and ABF3 play an important role in N response by means of regulating a series of genes related to NO3−–N response. ABF2 and ABF3 also have important roles in drought resistance; therefore, those two TFs may act as the main contact channel integrating NO3−–N, ABA and drought response signals [100]. OsABF1 also can regulate the expression of genes responding to both drought stress and N status, which links gene expression to phenotypic responses under low-N conditions and drought stress [120].
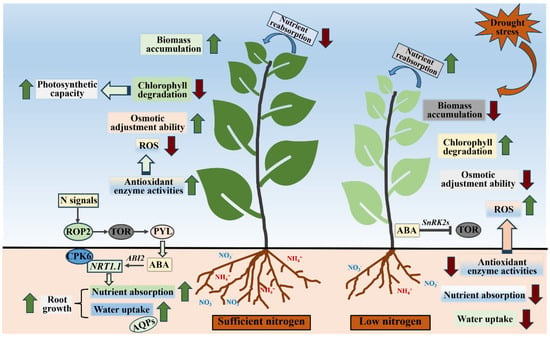
Figure 4.
Mitigation effect of sufficient nitrogen supply on drought stress in plants.
In contrast, Friedrich et al. [121] showed that increasing N application under drought conditions can promote the growth of above-ground plant components, enhance leaf transpiration and accelerate water loss, which are not conducive to plant resistance to drought stress. In addition, enhanced N application in beech (Fagus sylvatica) has been observed not to be conducive to root growth, preventing the plant from effectively absorbing water from the soil and consequently promoting drought stress damage to the plant. In this case, reducing the application rate of N fertilizer can be beneficial to plant resistance under drought stress [122]. Therefore, the effects of N application on plant response to drought stress need to be further studied in different species.
4.5. Nitrogen Regulates Anthocyanin Synthesis
Anthocyanins belong to a class of flavonoids that play an important role in both biotic and abiotic stresses in plants [123]. These substances typically accumulate in the vacuoles of plants and give plant organs a pink to purple color, which is useful for pollination and seed dispersal [124]. In addition, anthocyanins are important ROS scavengers, and thus anthocyanin-rich fruits are favored by consumers [125,126]. N is also an important factor regulating anthocyanin synthesis in plants [127,128]. Low-N conditions generally promote anthocyanin accumulation, while high N inhibits anthocyanin synthesis and accumulation in plants [129,130,131]. RNA interference of proline-rich protein 6 (MdPRP6) proves that plant MdPRP6 negatively regulates low-N tolerance by affecting anthocyanin synthesis and IAA concentration in apple (Malus domestica) roots [96]. In Arabidopsis, nitrate reduces anthocyanin accumulation mainly by inhibiting PRODUCTION OF ANTHOCYANIN PIGMENT (PAP) gene expression. Members of the LATERAL ORGAN BOUNDARIES DOMAIN (LBD) family, which are induced by nitrate, act as negative regulators of AtPAPs to regulate the accumulation of anthocyanin [132]. DELLAs can interact with PAP1 and enhance the transcriptional activity of PAP1, which plays an important role in the accumulation of anthocyanins induced by N deficiency and the regulation of plant adaptation to low-N stress [126]. MYB genes are involved in the anthocyanin accumulation process induced by nitrate at the transcriptional level. Research has revealed that the MYB10 transcription factor plays an important role in the color formation of red-fleshed apple and can improve nitrate uptake efficiency by transcriptionally activating the expression of MdNRT2.4-1 [133]. Wang et al. [134] found that the transcriptional and posttranslational modulation of MdMYB1 (a master regulator of anthocyanin biosynthesis and fruit coloration) occurred in response to nitrate treatment, which consequently degraded via a ubiquitin-dependent pathway in response to nitrate, ultimately inhibiting anthocyanin synthesis in apple.
4.6. Nitrogen and Mycorrhizal Symbiosis
Mycorrhizas are mutualistic symbionts formed by mycorrhizal fungi in soil and plant roots and are universally observed in nature [135]. Ectomycorrhizal (ECM) fungi can produce exocellular hydrolytic enzymes then obtain small organic N-bearing molecules from soil organic matter [136]. Arbuscular mycorrhizas (AMs) are the most widely distributed endophytic mycorrhizas in nature and play an important role in the rhizosphere of plants [137,138]. AM symbiosis can effectively improve the N response of N-source absorption and utilization by host plants, transferring N between fungi and host plants, as well as between different plants via hyphal bridges [135]. ECM fungi, to some extent, will help plants access organic N from soil organic matter and transfer it to host plants [139]. Interestingly, soil N availability in AM forests is higher than that in ECM forests. This may be attributed to higher N decomposition due to the greater nitrification rates facilitated by lower acidity and higher ammonium pools in AM forests compared to ECM forests [140]. The root mycorrhizal community will also change with the mycorrhizal inoculum species and the soil’s available N content [141,142].
Arbuscular mycorrhizal fungi (AMFs) have a system for absorbing N from the soil, and three AMT genes (GintAMT1, GintAMT2 and GintAMT3) have been identified from Rhizophagus irregularis [143]. GintAMT1 and GintAMT2 are expressed in both extraradical and intraradical mycelium, and they are involved in the NH4+ absorption of AMFs and recovery of NH4+ loss at the symbiotic interface due to fungal metabolism [144]. GintAMT3 is highly expressed only in intraradical mycelium, and its expression is regulated by matrix concentration and carbon source [145]. The process of N absorption from peri-arbuscular space to symbiotic plants mainly occurs through N transporters, which are located in plants’ peri-arbuscular membranes, such as the AMF-induced NO3− transporter OsNPF4.5, the amino acid transporter LjLHT1.2 and the NH4+ transporter LjAMT2;2 [146,147,148,149]. The expression level of NPF4.5 homologs in maize (Zea mays) and sorghum (Sorghum bicolor) is also induced after AMF treatment, demonstrating that the NO3− absorption process is highly activated by AM symbiosis [146]. Saia et al. [150] found that AMFs can significantly increase the expression of NRT1.1, AMT1;2 and AMT2;1 in durum wheat (Triticum durum Desf.). In addition, ZmAMT3;1, which is induced by AMFs and specifically expressed in cortical cells, can absorb 68%–74% of the N transported from AMFs to maize plants [151]. The extracorporeal hyphae of AMFs can expand the absorption of nutrients and develop the root system of mycorrhizal plants. This also promotes the absorption and utilization of N (Figure 5). Moreover, leguminous plants inoculated with mycorrhizal and N-fixing microorganisms can form root nodules while developing mycorrhizal symbionts. This trinity symbiosis system provides sufficient N nutrition for host plants.
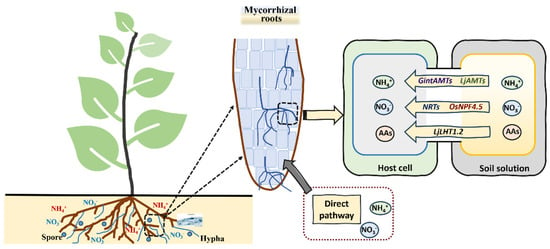
Figure 5.
The nitrogen uptake by plants in mycorrhizal roots. Mycorrhizal plants can absorb both NO3− and NH4+, as well as organic N forms, such as amino acids (AAs), via either the direct pathway or the mycorrhizal pathway.
4.7. Nitrogen and Rhizobium–Legume Symbiosis
Legume roots can form symbiotic nodules with rhizobia. Then, bacteria fix and reduce N2 to NH4+, which will be assimilated into amino acids by plants. Some abiotic stresses may reduce symbiotic activity and lead to plant N deficiency. Then, systemic signaling may compensate for the N deficit by stimulating symbiotic root N foraging. Furthermore, N also plays an important role in rhizobium–legume symbiosis. The autoregulation of nodule number (AON) is also affected by plant N demand. Under N deficiency, the symbiotic plant will release the AON repression signal to increase the nodule number [152]. NO3−–N application can reduce symbiotic N fixation via changing nodule gene expression and activating local and long-distance (systemic) responses [153]. Some TFs, such as NLPs, regulate the NO3−–N inhibition nodulation formation process [154]. Some members of the N uptake and transport gene family have been reported to be associated with nodule induction in legumes, such as Medicago truncatula NPF7.6, Lotus japonicus NRT2.4 and AMT1;1 [155,156,157].
5. Conclusions and Future Perspectives
Nitrogen (N) fertilizer application increases forests’ productivity and enhances the carbon sequestration of forest stands. However, overuse of N fertilizers increases the release of nitrous oxide (N2O), a greenhouse gas more harmful to global warming than CO2 [158]. Therefore, it is urgent to improve the NUE of forest ecosystem to reduce the application of N fertilizer. With global environmental changes, plants in forest ecosystems will not only suffer from low-N stress in the growth process, but this will also be accompanied by other stresses. It is important to gain a comprehensive understanding of plant N uptake and its regulation of other physiological processes. This review focused on the absorption, assimilation and regulation of N in plants, and it subsequently explained the interaction between N and hormone signals, miRNA regulation, lateral root growth, drought resistance, anthocyanin synthesis and mycorrhizal symbiosis (Figure 6). The in-depth understanding of N’s uptake and regulation of other physiological processes and stresses can improve forest ecosystem NUE and reduce the environmental pollution caused by excessive N application.
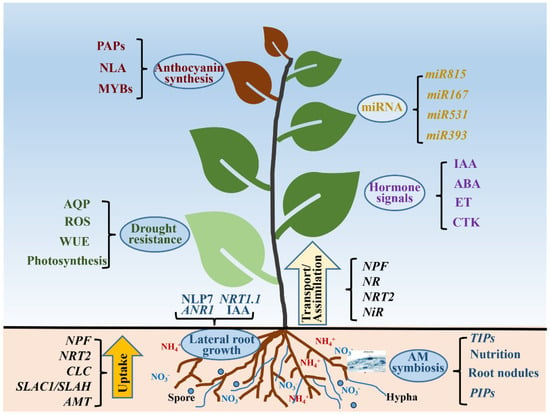
Figure 6.
Uptake and some functions of nitrogen in plants. AQP: aquaporin; ROS: reactive oxygen species; WUE: water use efficiency; AM: arbuscular mycorrhiza; IAA: auxin; ABA: abscisic acid; ET: ethylene; CTK: cytokinins; TIPs: plasma membrane intrinsic proteins; PIPs: plasma membrane intrinsic proteins; NPF: nitrate transporter 1/peptide transporter family; NRT2: nitrate transporter 2 family; CLC: chloride channel; SLAC1/SLAH: slow anion channel associated 1/slow anion channel homologues; AMT: ammonium transporter.
The improvement of plant NUE can be considered from the following perspectives: (1) Use reverse genetic strategies to identify new regulators for plant responses to N starvation stress based on the phenotypes of primary root elongation and lateral root genesis. (2) Depend on increasingly sophisticated whole-genome sequencing technologies to expand the scope of research on plant responses to N signals and provide optimal gene resources for breeding. (3) Through further research on the relationship between N signaling and a variety of abiotic stresses, improve plant NUE by allowing plants to adapt to increasingly severe environmental challenges.
Author Contributions
Acquired funding and wrote the manuscript, Q.W.; contributed to improving the presentation of the manuscript, S.L. and J.L.; acquired funding and supervision, D.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Science and Technology Project of Guizhou Province (Grant number: Qiankehe-ZK [2023]103 and 101); the Scientific Research Project of General Undergraduate Universities (Youth Project) of Guizhou (Grant number: Qianjiaoji [2022]117); the Open Project Fund of the Key Laboratory of Plant Resources Conservation and Germplasm Innovation in Mountainous Region (Ministry of Education) (Grant number: Qianjiaoji [2022]431); and the Natural Science Foundation of Guizhou University (Grant number: 600106).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- O′Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, Q.; Wang, X.; Zhang, J.; Ismail, A.M.; Zhang, Z. Nitrogen form-mediated ethylene signal regulates root-to-shoot K+ translocation via NRT1.5. Plant Cell Environ. 2021, 44, 3806–3818. [Google Scholar] [CrossRef]
- Jiang, H.; Qi, C.H.; Gao, H.N.; Feng, Z.Q.; Wu, Y.T.; Xu, X.X.; Cui, J.Y.; Wang, X.F.; Lv, Y.H.; Gao, W.S.; et al. MdBT2 regulates nitrogen-mediated cuticular wax biosynthesis via a MdMYB106-MdCER2L1 signalling pathway in apple. Nat. Plants 2024, 10, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, Z.; Li, J.; Bian, N.; Guo, Z.; Guo, J.; Zhao, S.; Chu, B.; Niu, C.; Ma, F.; et al. Interfering small ubiquitin modifiers (SUMO) exhibits apple’s enhanced tolerance to nitrogen deficiency. Fruit Res. 2023, 3, 24. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Luo, L.; Zhao, C.; Zheng, D.; Wang, E.; Liang, J.; Yin, C. Nitrogen uptake preference and allocation in Populus cathayana in response to drought stress. Environ. Exp. Bot. 2023, 213, 105415. [Google Scholar] [CrossRef]
- Ma, Q.; Zhao, C.; Hu, S.; Zuo, K. Arabidopsis calcium-dependent protein kinase CPK6 regulates drought tolerance under high nitrogen by the phosphorylation of NRT1.1. J. Exp. Bot. 2023, 74, 5682–5693. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ma, W.; Song, J.; Lu, M.; Rahman, S.; Bui, T.T.X.; Vu, D.D.; Zheng, H.; Wang, J.; Zhang, Y. Physiological and transcriptional responses of Catalpa bungei to drought stress under sufficient- and deficient-nitrogen conditions. Tree Physiol. 2017, 247, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Cao, X.; Zhang, M. Plant nitrogen availability and crosstalk with phytohormones signallings and their biotechnology breeding application in crops. Plant Biotechnol. J. 2023, 21, 1320–1342. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate transport, signaling, and use efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Liu, M.; Lin, Z.; Wang, Z.; Chen, B.; Liu, C.; Guo, A.; Konishi, M.; Yanagisawa, S.; Wagner, G.; et al. NIN-like protein 7 transcription factor is a plant nitrate sensor. Science 2022, 377, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, S.; Chaput, V.; Przybyla-Toscano, J.; Fayos, I.; Ibarra, C.; Moyano, T.; Fizames, C.; Tillard, P.; O’Brien, J.A.; Gutiérrez, R.A.; et al. Genome-wide analysis in response to nitrogen and carbon identifies regulators for root AtNRT2 transporters. Plant Physiol. 2021, 186, 696–714. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of Nitrate Transporter 1/Peptide Transporter family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Tsay, Y.F.; Schroeder, J.I.; Feldmann, K.A.; Crawford, N.M. The herbicide sensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitrate transporter. Cell 1993, 72, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, B.; Li, A.; Chu, C. NRT1.1s in plants: Functions beyond nitrate transport. J. Exp. Bot. 2020, 71, 4373–4379. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, T.J. CLC chloride channels and transporters, from genes to protein structure, pathology and physiology. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 3–36. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, X.; Ye, S.; Zheng, J.; Huang, X.; Yu, F.; Chen, Z.; Cai, S.; Zhang, P. Molecular mechanism underlying regulation of Arabidopsis CLCa transporter by nucleotides and phospholipids. Nat. Commun. 2023, 14, 4879. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Song, H.; Liao, Q.; Yu, Y.; Jian, S.; Lepo, J.E.; Liu, Q.; Rong, X.; Tian, C.; Zeng, J.; et al. Nitrogen use efficiency is mediated by vacuolar nitrate sequestration capacity in roots of Brassica napus. Plant Physiol. 2016, 170, 1684–1698. [Google Scholar] [CrossRef] [PubMed]
- Hodin, J.; Lind, C.; Marmagne, A.; Espagne, C.; Bianchi, M.W.; Angeli, A.D.; Abou-Choucha, F.; Bourge, M.; Chardon, F.; Thomine, S.; et al. Proton exchange by the vacuolar nitrate transporter CLCa is required for plant growth and nitrogen use efficiency. Plant Cell 2022, 35, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Fecht-Bartenbach, J.; Bogner, M.; Dynowski, M.; Ludewig, U. CLCb-mediated NO3−/H+ exchange across the tonoplast of Arabidopsis vacuoles. Plant Cell Physiol. 2010, 51, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, D.; He, Y.; Tang, J.; Chen, H.; Gong, P.; Luo, J.; Zhang, Z. CHLORIDE CHANNEL-b mediates vacuolar nitrate efflux to improve low nitrogen adaptation in Arabidopsis. Plant Physiol. 2023, 193, 1987–2002. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Fang, X.; Xiao, C.; Ma, Z.; Huang, X.; Su, J.; Li, J.; Wang, J.; Wang, S.; Luan, S.; et al. Kinase SnRK1.1 regulates nitrate channel SLAH3 engaged in nitrate-dependent alleviation of ammonium toxicity. Plant Physiol. 2021, 186, 731–749. [Google Scholar] [CrossRef] [PubMed]
- Maierhofer, T.; Lind, C.; Huttl, S.; Scherzer, S.; Papenfuss, M.; Simon, J.; Al-Rasheid, K.A.; Ache, P.; Rennenberg, H.; Hedrich, R.; et al. A single-pore residue renders the Arabidopsis root anion channel SLAH2 highly nitrate selective. Plant Cell 2014, 26, 2554–2567. [Google Scholar] [CrossRef] [PubMed]
- Geiger, D.; Maierhofer, T.; Al-Rasheid, K.A.; Scherzer, S.; Mumm, P.; Liese, A.; Ache, P.; Wellmann, C.; Marten, I.; Grill, E. Stomatal closure by fast abscisic acid signaling is mediated by the guard cell anion channel SLAH3 and the receptor RCAR1. Sci. Signal. 2011, 4, ra32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Sun, D.; Liu, B.; Fang, X.; Li, P.; Jiang, Y.; He, M.; Li, J.; Luan, S.; He, K. Nitrate transporter NRT1.1 and anion channel SLAH3 form a functional unit to regulate nitrate-dependent alleviation of ammonium toxicity. J. Integr. Plant Biol. 2022, 64, 942–957. [Google Scholar] [CrossRef]
- Jung, J.H.; Li, Z.; Chen, H.; Yang, S.; Li, D.; Priatama, R.A.; Kumar, V.; Xuan, Y.H. Mutation of phytochrome B promotes resistance to sheath blight and saline-alkaline stress via increasing ammonium uptake in rice. Plant J. 2023, 113, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef]
- Giehl, R.; Laginha, A.; Duan, F.; Rentsch, D.; Yuan, L.; Wirén, N. A critical role of AMT2;1 in root-to-shoot translocation of ammonium in Arabidopsis. Mol. Plant 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Suenaga, A.; Moriya, K.; Sonoda, Y.; Ikeda, A.; Wiren, N.; Hayakawa, T.; Yamaguchi, J.; Yamaya, T. Constitutive expression of a novel-type ammonium transporter OsAMT2 in rice plants. Plant Cell Physiol. 2003, 44, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Tami, M.; Fred, D.; Franois, L. Multiple horizontal gene transfers of ammonium transporters/ammonia permeases from prokaryotes to eukaryotes: Toward a new functional and evolutionary classification. Mol. Biol. Evol. 2011, 29, 51–60. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, W.; Wu, J.; Xie, K.; Li, X. LjAMT2;2 promotes ammonium nitrogen transport during arbuscular mycorrhizal fungi symbiosis in Lotus japonicus. Int. J. Mol. Sci. 2022, 23, 9522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhao, Y.; Luo, W.; Li, R.; He, Q.; Fang, X.; Michele, R.D.; Ast, C.; Wirén, N.V.; Lin, J. Single-particle analysis reveals shutoff control of the Arabidopsis ammonium transporter AMT1;3 by clustering and internalization. Proc. Natl. Acad. Sci. USA 2013, 110, 13204–13209. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, Y.; Lai, L.; Liu, X.; Miao, C.; Liu, R.; Li, X.; Tan, J.; Gao, Z.; Chen, J. OsAMT1.1 expression by nitrate-inducible promoter of OsNAR2.1 increases nitrogen use efficiency and rice yield. Rice Sci. 2023, 30, 222–234. [Google Scholar] [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; Wirén, N. AtAMT1;4, a pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Transporters involved in mineral nutrient uptake in rice. J. Exp. Bot. 2016, 67, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Yue, X.; Fang, S.; Qian, M.; Zhou, S.; Shang, X.; Yang, W. Responses of nitrogen metabolism, photosynthetic parameter and growth to nitrogen fertilization in Cyclocarya paliurus. For. Ecol. Manag. 2021, 502, 119715. [Google Scholar] [CrossRef]
- Huang, W.T.; Zheng, Z.C.; Hua, D.; Chen, X.F.; Zhang, J.; Chen, H.H.; Ye, X.; Guo, J.X.; Yang, L.T.; Chen, L.S. Adaptive responses of carbon and nitrogen metabolisms to nitrogen-deficiency in Citrus sinensis seedlings. BMC Plant Biol. 2022, 22, 370. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, K.; Yamaya, T.; Hayakawa, T.; Mae, T.; Ojima, K. Vascular bundle-specific localization of cytosolic glutamine synthetase in rice leaves. Plant Physiol. 1992, 99, 1481–1486. [Google Scholar] [CrossRef]
- Avila-Ospina, L.; Marmagne, A.; Talbotec, J.; Krupinska, K.; Masclaux-Daubresse, C. The identification of new cytosolic glutamine synthetase and asparagine synthetase genes in barley (Hordeum vulgare L.), and their expression during leaf senescence. J. Exp. Bot. 2015, 66, 2013–2026. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.H.; Feraud, M.; Jouglet, T.; Suzuki, A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, C.; Huang, D.; Dong, A.; Li, P.; Ma, F. High-efficient utilization and uptake of N contribute to higher NUE of ‘Qinguan’ apple under drought and N-deficient conditions compared with ‘Honeycrisp’. Tree Physiol. 2019, 39, 1880–1895. [Google Scholar] [CrossRef]
- Liu, K.H.; Niu, Y.; Konishi, M.; Wu, Y.; Du, H.; Chung, H.S.; Li, L.; Boudsocq, M.; McCormack, M.; Maekawa, S.; et al. Discovery of nitrate-CPK-NLP signalling in central nutrient growth networks. Nature 2017, 545, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Brehaut, V.; Clement, G.; Kelemen, Z.; Macé, J.; Feil, R.; Duville, G.; Launay-Avon, A.; Roux, C.P.L.; Lunn, J.E.; et al. The Arabidopsis transcription factor NLP2 regulates early nitrate responses and integrates nitrate assimilation with energy and carbon skeleton supply. Plant Cell 2023, 35, 1429–1454. [Google Scholar] [CrossRef] [PubMed]
- Bouguyon, E.; Perrine-Walker, F.; Pervent, M.; Bach, L.; Krouk, G.; Gojon, A.; Nacry, P. Nitrate controls root development through posttranscriptional regulation of the NRT1.1/NPF6.3 transporter/sensor. Plant Physiol. 2016, 172, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Tsay, Y.F. Switching between the two action modes of the dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J. 2003, 22, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Bellegarde, F.; Gojon, A.; Martin, A. Signals and players in the transcriptional regulation of root responses by local and systemic N signaling in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 2553–2565. [Google Scholar] [CrossRef]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef] [PubMed]
- Medici, A.; Marshall-Colon, A.; Ronzier, E.; Szponarski, W.; Wang, R.C.; Gojon, A.; Crawford, N.M.; Ruffe, S.; Coruzzi, G.M.; Krouk, G. AtNIGT1/HRS1 integrates nitrate and phosphate signals at the Arabidopsis root tip. Nat. Commun. 2015, 6, 6274. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Li, T.; Wang, X.; Sun, W.; Zhang, T.; You, C.; Wang, X. Identification and characterization of apple MdNLP7 transcription factor in the nitrate response. Plant Sci. 2022, 316, 111158. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Liu, Z.; Guo, J.; Jia, Z.; Shi, Y.; Kang, K.; Peng, W.; Wang, Z.; Chen, L.; Neuhaeuser, B.; et al. ZmNRT1.1B (ZmNPF6.6) determines nitrogen use efficiency via regulation of nitrate transport and signalling in maize. Plant Biotechnol. J. 2024, 22, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Armijo, G.; Gutiérrez, R.A. Emerging players in the nitrate signaling pathway. Mol. Plant 2017, 10, 1019–1022. [Google Scholar] [CrossRef] [PubMed]
- Ródenas, R.; Vert, G. Regulation of root nutrient transporters by CIPK23: ‘One kinase to rule them all’. Plant Cell Physiol. 2021, 62, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, M.; Yang, J.; Li, M.; Zhang, Z.; Gao, H.; Wang, C.; Tian, H. Brassinosteroid transcription factor BES1 modulates nitrate deficiency by promoting NRT2.1 and NRT2.2 transcription in Arabidopsis. Plant J. 2023, 114, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, Z.; Sun, H.; Liu, X.; Xie, J.; Qiu, Y.; Chai, T.; Chu, C.; Hu, B. Nitrate confers rice adaptation to high ammonium by suppressing its uptake but promoting its assimilation. Mol. Plant 2023, 16, 1871–1874. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Edel, K.H.; Pervent, M.; Hashimoto, K.; Corratge-Faillie, C.; Offenborn, J.N.; Tillard, P.; Gojon, A.; Kudla, J.; Lacombe, B. Nitrate sensing and uptake in Arabidopsis are enhanced by ABI2, a phosphatase inactivated by the stress hormone abscisic acid. Sci. Signal. 2015, 8, ra43. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Lin, S.H.; Hu, H.C.; Tsay, Y.F. CHL1 functions as a nitrate sensor in plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Duan, Y.; Ye, Q.; Ma, Y.; Ma, R.; Zhao, L.; Zhu, S.; Yu, F.; Qi, S.; Wang, Y. The Arabidopsis eIF4E1 regulates NRT1.1-mediated nitrate signaling at both translational and transcriptional levels. New Phytol. 2023, 240, 338–353. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Wang, Q.; Ye, Y.; Li, T.; Sun, X.; Huo, L.; Wang, P.; Gong, X.; Ma, F. MdATG5a positively regulates nitrogen uptake under low nitrogen conditions by enhancing the accumulation of flavonoids and auxin in apple roots. Environ. Exp. Bot. 2022, 197, 104840. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Trivellini, A.; Fatma, M.; Masood, A.; Francini, A.; Iqbal, N.; Ferrante, A.; Khan, N.A. Role of ethylene in responses of plants to nitrogen availability. Front. Plant Sci. 2015, 6, 927. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Ma, T.; Xian, W.; Hu, B.; Chu, C. Interplay between ethylene and nitrogen nutrition: How ethylene orchestrates nitrogen responses in plants. J. Integr. Plant Biol. 2023, 65, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Guinel, F.C. Ethylene, a hormone at the center-stage of nodulation. Front. Plant Sci. 2015, 6, 1121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yi, H.; Gong, J. The Arabidopsis ethylene/jasmonic acid-NRT signaling module coordinates nitrate reallocation and the trade-off between growth and environmental adaptation. Plant Cell 2014, 26, 3984–3998. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Sun, P.; Zhang, W. Ethylene is involved in nitrate-depend root growth and branching in Arabidopsis thaliana. New Phytol. 2009, 184, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Gernier, H.D.; Pessemier, J.D.; Xu, J.; Cristescu, S.M.; Straeten, D.V.D.; Verbruggen, N.; Hermans, C. A comparative study of ethylene emanation upon nitrogen deficiency in natural accessions of Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, Q.; Jing, G.; Ma, M.; Li, C.; Ma, F. Overexpression of MdIAA24 improves apple drought resistance by positively regulating strigolactone biosynthesis and mycorrhization. Tree Physiol. 2021, 41, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Li, J.; Qu, B.; He, X.; Zhao, X.; Li, B.; Fu, X.; Tong, Y. Auxin biosynthetic gene TAR2 is involved in low nitrogen mediated reprogramming of root architecture in Arabidopsis. Plant J. 2014, 78, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qi, T.; Hu, Z.; Fan, X.; Zhu, L.; Iqbal, M.; Yin, X.; Xu, G.; Fan, X. OsNAR2.1 positively regulates drought tolerance and grain yield under drought stress conditions in rice. Front. Plant Sci. 2019, 10, 197. [Google Scholar] [CrossRef] [PubMed]
- Hauser, F.; Li, Z.; Waadt, R.; Schroeder, J.I. SnapShot: Abscisic acid signaling. Cell 2017, 171, 1708–1708.e1. [Google Scholar] [CrossRef]
- Ondzighi-Assoume, C.A.; Chakraborty, S.; Harris, J.M. Environmental nitrate stimulates abscisic acid accumulation in Arabidopsis root tips by releasing it from inactive stores. Plant Cell 2016, 28, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Easwaran, V.; Chau, V.; Okamoto, M.; Ierullo, M.; Kimura, M.; Endo, A.; Yano, R.; Pasha, A.; Gong, Y.; et al. NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis. Nat. Commun. 2016, 7, 13179. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Wang, T.; Ju, C.; Deng, J.; Zhang, T.; Li, M.; Tian, H.; Wang, C. Abscisic acid signaling negatively regulates nitrate uptake via phosphorylation of NRT1.1 by SnRK2s in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, P.; Liu, Q.; Li, G.; Di, D.; Xia, G.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W. TaANR1-TaBG1 and TaWabi5-TaNRT2s/NARs link ABA metabolism and nitrate acquisition in wheat roots. Plant Physiol. 2020, 182, 1440–1453. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H. Cytokinin biosynthesis and transport for systemic nitrogen signaling. Plant J. 2021, 105, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xing, X.; Wang, Y.; Tran, A.; Crawford, N.M. A genetic screen for nitrate regulatory mutants captures the nitrate transporter gene NRT1.1. Plant Physiol. 2009, 151, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Wang, R.; Nacry, P.; Breton, G.; Kay, S.A.; Pruneda-Paz, J.L.; Davani, A.; Crawford, N.M. Nitrate foraging by Arabidopsis roots is mediated by the transcription factor TCP20 through the systemic signaling pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 15267–15272. [Google Scholar] [CrossRef] [PubMed]
- Hervé, C.; Dabos, P.; Bardet, C.; Jauneau, A.; Auriac, M.C.; Ramboer, A.; Lacout, F.; Tremousaygue, D. In vivo interference with AtTCP20 function induces severe plant growth alterations and deregulates the expression of many genes important for development. Plant Physiol. 2009, 149, 1462–1477. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Du, C.; Gao, H.; Liu, S.; Sun, W.; Lu, H.; Kang, J.; Xie, Y.; Ma, D.; Wang, C. Identification of microRNAs in developing wheat grain that are potentially involved in regulating grain characteristics and the response to nitrogen levels. BMC Plant Biol. 2020, 22, 87. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Chai, X.; Foster, T.M.; Deng, C.H.; Wu, T.; Zhang, X.; Han, Z.; Wang, Y. miR164-MhNAC1 regulates apple root nitrogen uptake under low nitrogen stress. New Phytol. 2024, 242, 1218–1237. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, D.L.; Paola, D.D.; Janni, M.; Curci, P.L.; Sonnante, G. Durum wheat miRNAs in response to nitrogen starvation at the grain filling stage. PLoS ONE 2017, 12, e0183253. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Estelle, M. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Chen, Y.; Cao, Y.; Chen, H.; Wang, J.; Bi, Y.; Tian, F.; Yang, F.; Rothstein, S.J.; Zhou, X.; et al. Overexpression of miR169o, an overlapping MicroRNA in response to both nitrogen limitation and bacterial infection, promotes nitrogen use efficiency and susceptibility to bacterial blight in rice. Plant Cell Physiol. 2018, 59, 1234–1247. [Google Scholar] [CrossRef] [PubMed]
- Gifford, M.L.; Dean, A.; Gutierrez, R.A.; Coruzzi, G.M.; Birnbaum, K.D. Cell-specific nitrogen responses mediate developmental plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.J.; Beatty, P.H.; Good, A.G.; Muench, D.G. Manipulation of microRNA expression to improve nitrogen use efficiency. Plant Sci. 2013, 210, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; Macpherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.A.; Chua, H.N. IAA-Ala resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.D.; Musialaklange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.R. Identification of nutrient-responsive Arabidopsis and rapeseed micro RNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wu, F.; Lin, Y.; Han, X.; Xu, X.; Zhang, Y.; Yang, Q.; Huang, H.; Tong, Z.; Zhang, J. A miR169c-NFYA10 module confers tolerance to low-nitrogen stress to Betula luminifera. Ind. Crops Prod. 2021, 172, 13988. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, H.; Hamera, S.; Chen, X.; Fang, R. miR444a has multiple functions in the rice nitrate-signaling pathway. Plant J. 2014, 78, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Sagwal, V.; Sihag, P.; Singh, Y.; Mehla, S.; Kapoor, P.; Balyan, P.; Kumar, A.; Mir, R.R.; Dhankher, O.P.; Kumar, U. Development and characterization of nitrogen and phosphorus use efficiency responsive genic and miRNA derived SSR markers in wheat. Heredity 2022, 128, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Bellegarde, F.; Sakakibara, H. Nitrate-dependent modulation of root system architecture in maize: A balance between strigolactone and auxin pathways. Plant Cell Physiol. 2021, 62, 541–542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gong, X.; Cheng, S.; Yu, H.; Li, D.; Su, X.; Lei, Z.; Li, M.; Ma, F. Proline-rich protein MdPRP6 alters low nitrogen stress tolerance by regulating lateral root formation and anthocyanin accumulation in transgenic apple (Malus domestica). Environ. Exp. Bot. 2022, 197, 104841. [Google Scholar] [CrossRef]
- Tahir, M.M.; He, X.; Liu, Y.; Raza, H.; Aziz, U.; Fan, L.; Asghar, Z.; Li, S.; Sun, S.; Zhang, D.; et al. Nitrate stimulates adventitious rooting by increasing auxin content and regulating auxin- and root development-related genes expression in apple. Hortic. Adv. 2023, 1, 18. [Google Scholar] [CrossRef]
- Rellan-Alvarez, R.; Lobet, G.; Dinneny, J.R. Environmental control of root system biology. Annu. Rev. Plant Biol. 2016, 67, 619–642. [Google Scholar] [CrossRef]
- Tokizawa, M.; Enomoto, T.; Chandnani, R.; Mora-Macías, J.; Burbridge, C.; Armenta-Medina, A.; Kobayashi, Y.; Yamamoto, Y.Y.; Koyama, H.; Kochian, L.V. The transcription factors, STOP1 and TCP20, are required for root system architecture alterations in response to nitrate deficiency. Proc. Natl. Acad. Sci. USA 2023, 120, e2300446120. [Google Scholar] [CrossRef] [PubMed]
- Contreras-López, O.; Vidal, E.A.; Riveras, E.; Alvarez, J.M.; Moyano, T.C.; Sparks, E.E.; Medina, J.; Pasquino, A.; Benfey, P.N.; Coruzzi, G.M.; et al. Spatiotemporal analysis identifies ABF2 and ABF3 as key hubs of endodermal response to nitrate. Proc. Natl. Acad. Sci. USA 2022, 119, e2107879119. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhu, M.; Jia, L.; Xie, Y.; Wang, Z.; Xuan, W. Ammonium transporters cooperatively regulate rice crown root formation responding to ammonium nitrogen. J. Exp. Bot. 2022, 73, 3671–3685. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Krall, L.; Li, Z.; Xi, L.; Luo, H.; Li, S.; He, M.; Yang, X.; Zan, H.; Gilbert, M.; et al. Transceptor NRT1.1 and receptor-kinase QSK1 complex controls PM H+-ATPase activity under low nitrate. Curr. Biol. 2024, 34, 1479–1491.e6. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ji, Z.; Tao, Y.; Wei, S.; Jiao, W.; Fang, Y.; Jian, P.; Shen, C.; Qin, Y.; Zhang, S.; et al. Improving rice nitrogen-use efficiency by modulating a novel monouniquitination machinery for optimal root plasticity response to nitrogen. Nat. Plants 2023, 9, 1902–1914. [Google Scholar] [CrossRef] [PubMed]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Auxin-mediated nitrate signalling by NRT1.1 participates in the adaptive response of Arabidopsis root architecture to the spatial heterogeneity of nitrate availability. Plant Cell Environ. 2014, 37, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Bouguyon, E.; Brun, F.; Meynard, D.; Kubeš, M.; Pervent, M.; Léran, S.; Lacombe, B.; Krouk, G.; Guiderdoni, E.; Zažímalová, E.; et al. Multiple mechanisms of nitrate sensing by Arabidopsis nitrate transceptor NRT1.1. Nat. Plants 2015, 1, 15015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cui, Y.; Yu, M.; Su, B.; Gong, W.; Baluska, F.; Komis, G.; Samaj, J.; Shan, X.; Lin, J. Phosphorylation-mediated dynamics of nitrate transceptor NRT1.1 regulate auxin flux and nitrate signaling in lateral root growth. Plant Physiol. 2019, 181, 480–498. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Bernreiter, A.; Filleur, S.; Abram, B.; Forde, B.G. Overexpressing the ANR1 MADS-box gene in transgenic plants provides new insights into its role in the nitrate regulation of root development. Plant Cell Physiol. 2012, 53, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Remans, T.; Nacry, P.; Pervent, M.; Filleur, S.; Diatloff, E.; Mounier, E.; Tillard, P.; Forde, B.G.; Gojon, A. The Arabidopsis NRT1.1 transporter participates in the signaling pathway triggering root colonization of nitrate-rich patches. Proc. Natl. Acad. Sci. USA 2006, 103, 19206–19211. [Google Scholar] [CrossRef] [PubMed]
- Marchive, C.; Roudier, F.; Castaings, L.; Brehaut, V.; Blondet, E.; Colot, V.; Meyer, C.; Krapp, A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 2013, 4, 1713. [Google Scholar] [CrossRef] [PubMed]
- Plett, D.C.; Ranathunge, K.; Melino, V.J.; Kuya, N.; Uga, Y.; Kronzucker, H.J. The intersection of nitrogen nutrition and water use in plants: New paths toward improved crop productivity. J. Exp. Bot. 2020, 71, 4452–4468. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Li, Y.; Gao, Y.; Shen, Q. Effects of different nitrogen forms and osmotic stress on water use efficiency of rice (Oryza sativa). Ann. Appl. Biol. 2008, 153, 127–134. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Mao, Q.Z.; Watanabe, M.; Imori, M.; Kim, Y.S.; Kita, K.; Koike, T. Photosynthesis and nitrogen allocation in needles in the sun and shade crowns of hybrid larch saplings: Effect of nitrogen application. Photosynthetica 2012, 50, 422–428. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, C.; Huang, D.; Dong, Q.; Li, P.; Ma, F. Physiological evaluation of nitrogen use efficiency of different apple cultivars under various nitrogen and water supply conditions. J. Integr. Agric. 2020, 19, 709–720. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, J.; Guo, J.; Wang, Y.; Ji, H.; Chu, X.; Xiao, K.; Qi, X.; Hu, L.; Li, H.; et al. GmTDN1 improves wheat yields by inducing dual tolerance to both drought and low-N stress. Plant Biotechnol. J. 2022, 20, 1606–1621. [Google Scholar] [CrossRef] [PubMed]
- Araus, V.; Swift, J.; Alvarez, J.M.; Henry, A.; Coruzzi, G.M. A balancing act: How plants integrate nitrogen and water signals. J. Exp. Bot. 2020, 71, 4442–4451. [Google Scholar] [CrossRef] [PubMed]
- Cerda, A.; Alvarez, J.M. Insights into molecular links and transcription networks integrating drought stress and nitrogen signaling. New Phytol. 2024, 241, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Duan, X.; Zhao, X.; Ding, W.; Wang, Y.; Xiong, Y. Diverse nitrogen signals activate convergent ROP2-TOR signaling in Arabidopsis. Dev. Cell 2021, 56, 1283–1295.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, Y.; Li, Z.; Hsu, C.C.; Liu, X.; Fu, L.; Hou, Y.J.; Du, Y.; Xie, S.; Zhang, C.; et al. Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response. Mol. Cell 2018, 69, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Shanks, C.M.; Huang, J.; Cheng, C.Y.; Shih, H.J.S.; Brooks, M.D.; Alvarez, J.M.; Araus, V.; Swift, J.; Henry, A.; Coruzzi, G.M. Validation of a high-confidence regulatory network for gene-to-NUE phenotype infield-grown rice. Front. Plant Sci. 2022, 13, 1006044. [Google Scholar] [CrossRef]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.U.; Schlebelmann, K.; Weber, M.S.; Härdtle, W. Nitrogen deposition increases susceptibility to drought-experimental evidence with the perennial grass Moliniacaerulea (L.). Moench. Plant Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Dziedek, C.; Härdtle, W.; Oheimb, G.; Fichtner, A. Nitrogen addition enhances drought sensitivity of young deciduous tree species. Front. Plant Sci. 2016, 7, 1100. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.R.; Zhao, Q.; Yang, Y.Y.; Zhang, T.E.; Wang, X.F.; You, C.X.; Hao, Y.J. The apple 14-3-3 protein MdGRF11 interacts with the BTB protein MdBT2 to regulate nitrate deficiency-induced anthocyanin accumulation. Hort. Res. 2021, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Venail, J.; Mackay, S.; Bailey, P.C.; Schwinn, K.E.; Jameson, P.E.; Martin, C.R.; Davies, K.M. The molecular basis for venation patterning of pigmentation and its effect on pollinator attraction in flowers of Antirrhinum. New Phytol. 2011, 189, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Liu, J.; Lin, S.; Wang, J.; Lin, W.; Xu, W. GA-DELLA pathway is involved in regulation of nitrogen deficiency-induced anthocyanin accumulation. Plant Cell Rep. 2017, 36, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Xiao, W.; Mu, Q.; Li, D.; Chen, X.; Wu, H.; Li, L.; Peng, F. How does nitrate regulate plant senescence? Plant Physiol. Biochem. 2020, 157, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Utasee, S.; Jamjod, S.; Lordkaew, S.; Prom-U-Thai, C. Improve anthocyanin and zinc concentration in purple rice by nitrogen and zinc fertilizer application. Rice Sci. 2022, 29, 435–450. [Google Scholar] [CrossRef]
- Fredes, I.; Moreno, S.; Diaz, F.P.; Gutierrez, R.A. Nitrate signaling and the control of Arabidopsis growth and development. Curr. Opin. Plant Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Gao, Y.; Han, M.; Liu, P.; Yang, C.; Shen, T.; Li, H. In vitro anthocyanin induction and metabolite analysis in Malus spectabilis leaves under low nitrogen conditions. Hortic. Plant J. 2020, 6, 284–292. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Yang, J.; Ma, C.; Zhang, Y.; Ge, T.; Qi, Z.; Kang, Y. Arabidopsis ROOT HAIR DEFECTIVE3 is involved in nitrogen starvation-induced anthocyanin accumulation. J. Integr. Plant Biol. 2015, 57, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Li, H.; An, X.; Song, L.; You, C.; Zhao, L.; Tian, Y.; Wang, X. MdMYB10 affects nitrogen uptake and reallocation by regulating the nitrate transporter MdNRT2.4-1 in red-fleshed apple. Hortic. Res. 2022, 9, uhac016. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; An, J.; Liu, X.; Su, L.; You, C.; Hao, Y. The nitrate-responsive protein MdBT2 regulates anthocyanin biosynthesis by interacting with the MdMYB1 transcription factor. Plant Physiol. 2018, 178, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Ren, Y.; Chen, A.; Yang, C.; Zheng, Q.; Chen, J.; Wang, D.; Li, Y.; Hu, S.; Xu, G. Plant nitrogen nutrition: The roles of arbuscular mycorrhizal fungi. J. Plant Physiol. 2022, 269, 153591. [Google Scholar] [CrossRef] [PubMed]
- Chalot, M.; Brun, A. Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas. FEMS Microbiol. Rev. 1998, 22, 21–44. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lozano, J.M.; Aroca, R.; Zamarreno, A.M.; Molina, S.; Andreo-Jimenez, B.; Porcel, R.; Garcia-Mina, J.M.; Ruyter-Spira, C.; Lopez-Raez, J.A. Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. Plant Cell Environ. 2016, 39, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, M.; Wang, Z.; Li, J.; Liu, K.; Huang, D. The role of arbuscular mycorrhizal symbiosis in plant abiotic stress. Front. Microbiol. 2024, 14, 1323881. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Pellitier, P.T.; Argiroff, W.A.; Castillo, B.; James, T.Y.; Nave, L.E.; Averill, C.; Beidler, K.V.; Bhatnagar, J.; Blesh, J.; et al. Exploring the role of ectomycorrhizal fungi in soil carbon dynamics. New Phytol. 2019, 223, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, C.; Taniguchi, T.; Du, S.; Yamanaka, N.; Tateno, R. Soil nitrogen cycling is determined by the competition between mycorrhiza and ammonia-oxidizing prokaryotes. Ecology 2020, 101, e02963. [Google Scholar] [CrossRef] [PubMed]
- Bittebiere, A.K.; Vandenkoornhuyse, P.; Maluenda, E.; Gareil, A.; Dheilly, A.; Coudouel, S.; Bahin, M.; Mony, C. Past spatial structure of plant communities determines arbuscular mycorrhizal fungal community assembly. J. Ecol. 2019, 108, 546–560. [Google Scholar] [CrossRef]
- Tatsumi, C.; Hyodo, F.; Taniguchi, T.; Shi, W.Y.; Koba, K.; Fukushima, K.; Du, S.; Yamanaka, N.; Templer, P.; Tateno, R. Arbuscular mycorrhizal community in roots and nitrogen uptake patterns of understory trees beneath ectomycorrhizal and non-ectomycorrhizal overstory trees. Front. Plant Sci. 2021, 11, 583585. [Google Scholar] [CrossRef] [PubMed]
- Kobae, Y.; Tamura, Y.; Takai, S.; Banba, M.; Hata, S. Localized expression of arbuscular mycorrhiza-inducible ammonium transporters in soybean. Plant Cell Physiol. 2010, 51, 1411–1415. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tienda, J.; Valderas, A.; Camañes, G.; García-Agustín, P.; Ferrol, N. Kinetics of NH4+ uptake by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2012, 22, 485–491. [Google Scholar] [CrossRef]
- Calabrese, S.; Pérez-Tienda, J.; Ellerbeck, M.; Arnould, C.; Chatagnier, O.; Boller, T.; Schüßler, A.; Brachmann, A.; Wipf, D.; Ferrol, N.; et al. GintAMT3-a low-affinity ammonium transporter of the arbuscular mycorrhizal Rhizophagus irregularis. Front. Plant Sci. 2016, 7, 679. [Google Scholar] [CrossRef]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef] [PubMed]
- Guether, M.; Volpe, V.; Balestrini, R.; Requena, N.; Wipf, D.; Bonfante, P. LjLHT1.2-a mycorrhiza-inducible plant amino acid transporter from Lotus japonicus. Biol. Fertil. Soils 2022, 47, 925–936. [Google Scholar] [CrossRef]
- Wang, Y.; Xuan, Y.; Wang, S.; Fan, D.; Wang, X.; Zheng, X. Genome-wide identification, characterization, and expression analysis of the ammonium transporter gene family in tea plants (Camellia sinensis L.). Physiol. Plant. 2022, 1, 174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, H.; Druzhinina, I.S.; Xie, X.; Wang, E.; Martin, F.; Yuan, Z. Phosphorus/nitrogen sensing and signaling in diverse root-fungus symbioses. Trends Microbiol. 2024, 32, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.; An, X.; Li, Z.; Neuhäuser, B.; Ludewig, U.; Wu, X.; Schulze, W.X.; Chen, F.; Feng, G.; Lambers, H.; et al. The mycorrhiza-specific ammonium transporter ZmAMT3;1 mediates mycorrhiza-dependent nitrogen uptake in maize roots. Plant Cell 2022, 34, 4066–4087. [Google Scholar] [CrossRef] [PubMed]
- Lepetit, M.; Brouquisse, R. Control of the rhizobium–legume symbiosis by the plant nitrogen demand is tightly integrated at the whole plant level and requires inter-organ systemic signaling. Front. Plant Sci. 2023, 14, 1114840. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, R. Nitrogen and phosphorus signaling and transport during Legume–rhizobium symbiosis. Front. Plant Sci. 2021, 12, 683601. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Li, X.; Luo, Z.; Mysore, K.S.; Wen, J.; Xie, F. NIN interacts with NLPs to mediate NO3− inhibition of nodulation in Medicago truncatula. Nat. Plants 2018, 4, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, Y.; Ren, Z.; Zhang, X.; Ren, J.; Su, J.; Zhang, C.; Tian, J.; Yu, Y.; Gao, G.F.; et al. Transfer cells mediate nitrate uptake to control root nodule symbiosis. Nat. Plants 2020, 6, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Valkov, V.T.; Sol, S.; Rogato, A.; Chiurazzi, M. The functional characterization of LjNRT2.4 indicates a novel, positive role of nitrate for an efficient nodule N2-ffxation activity. New Phytol. 2020, 228, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Rogato, A.; D’Apuzzo, E.; Barbulova, A.; Omrane, S.; Stedel, C.; Simon-Rosin, U.; Katinakis, P.; Flemetakis, M.; Udvardi, M.; Chiurazzi, M. Tissue-speciffc down-regulation of LjAMT1;1 compromises nodule function and enhances nodulation in Lotus japonicus. Plant Mol. Biol. 2008, 68, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Hou, Y.; Xu, X. Optimized strategies for nitrogen fertilizer application in populus plantations in the context of climate change mitigation. For. Policy Econ. 2024, 159, 103139. [Google Scholar] [CrossRef]

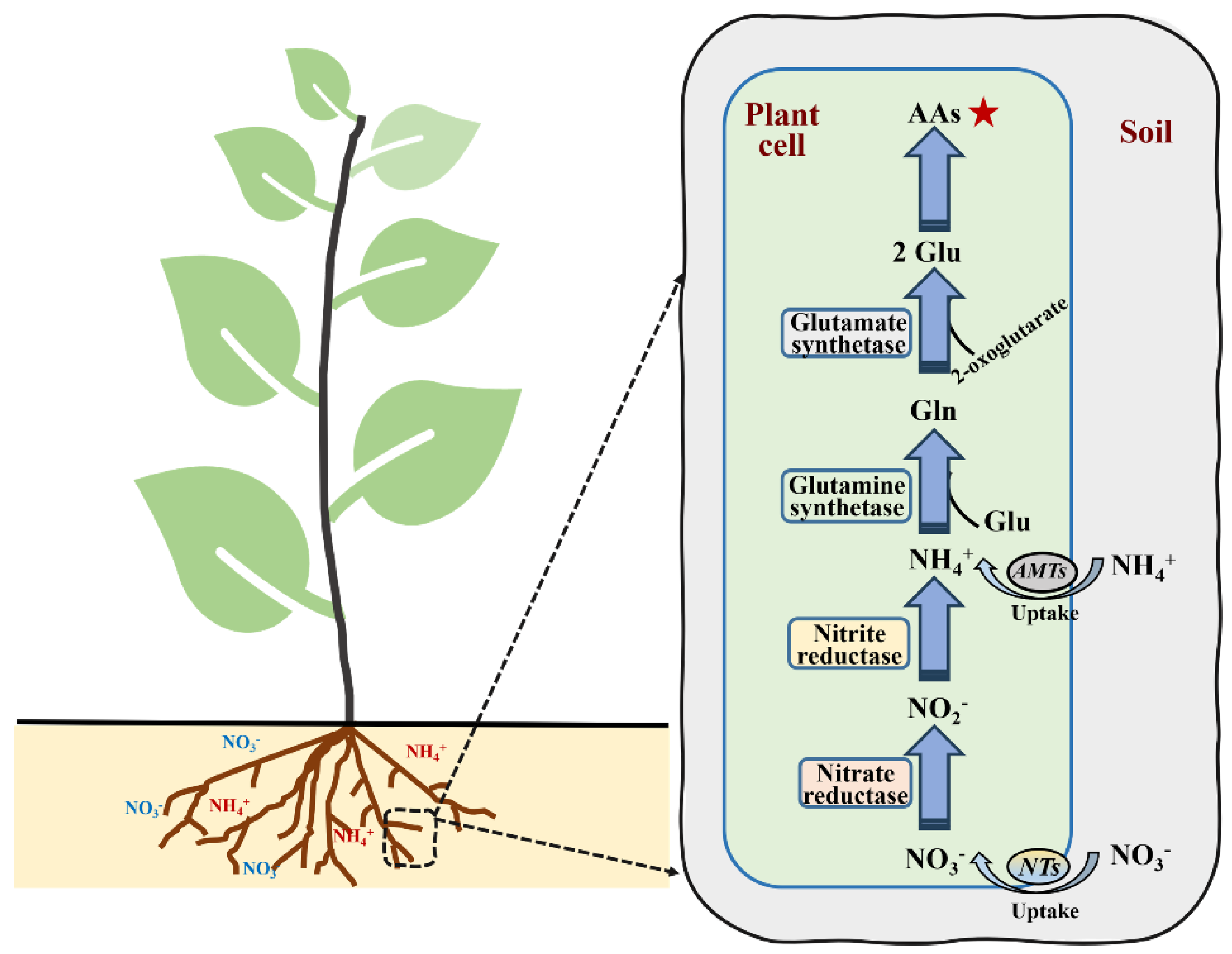


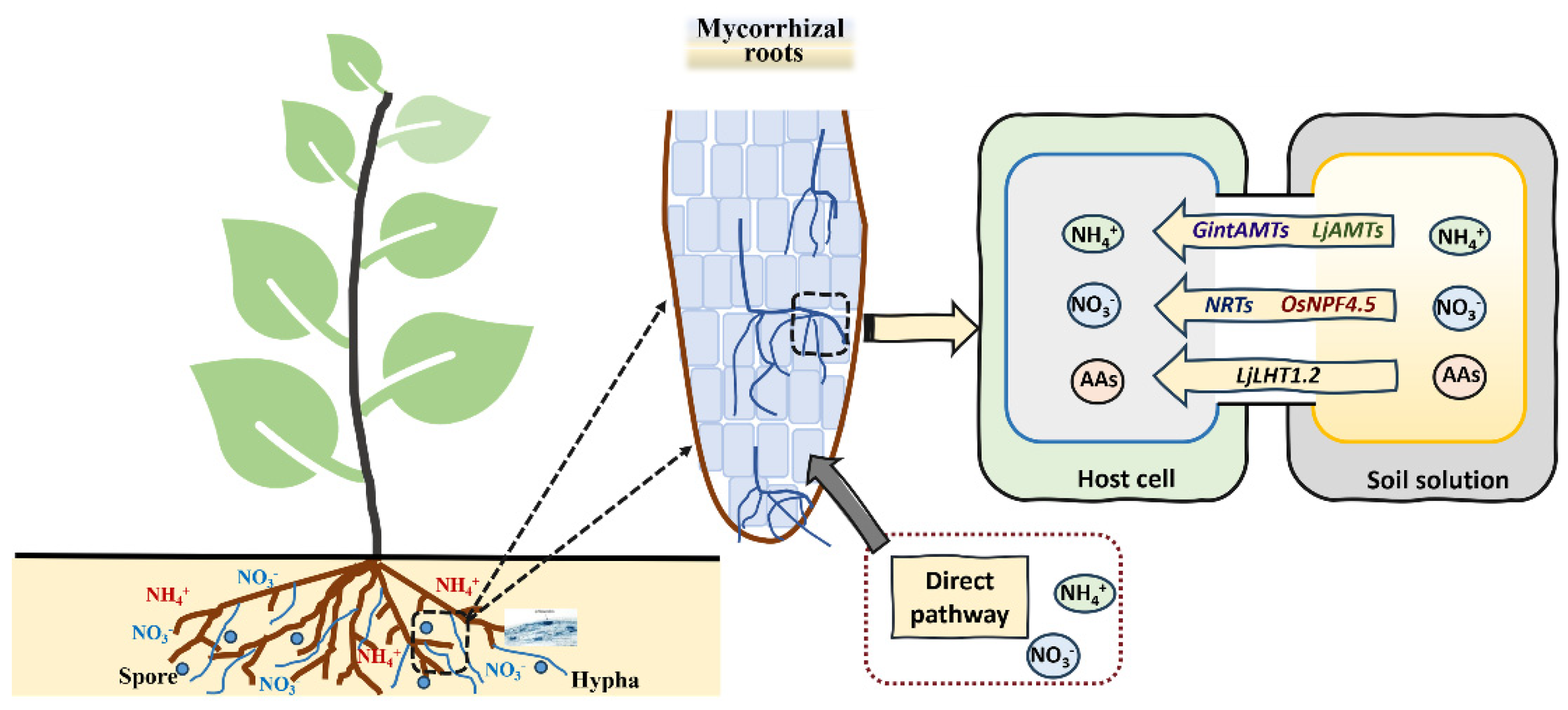
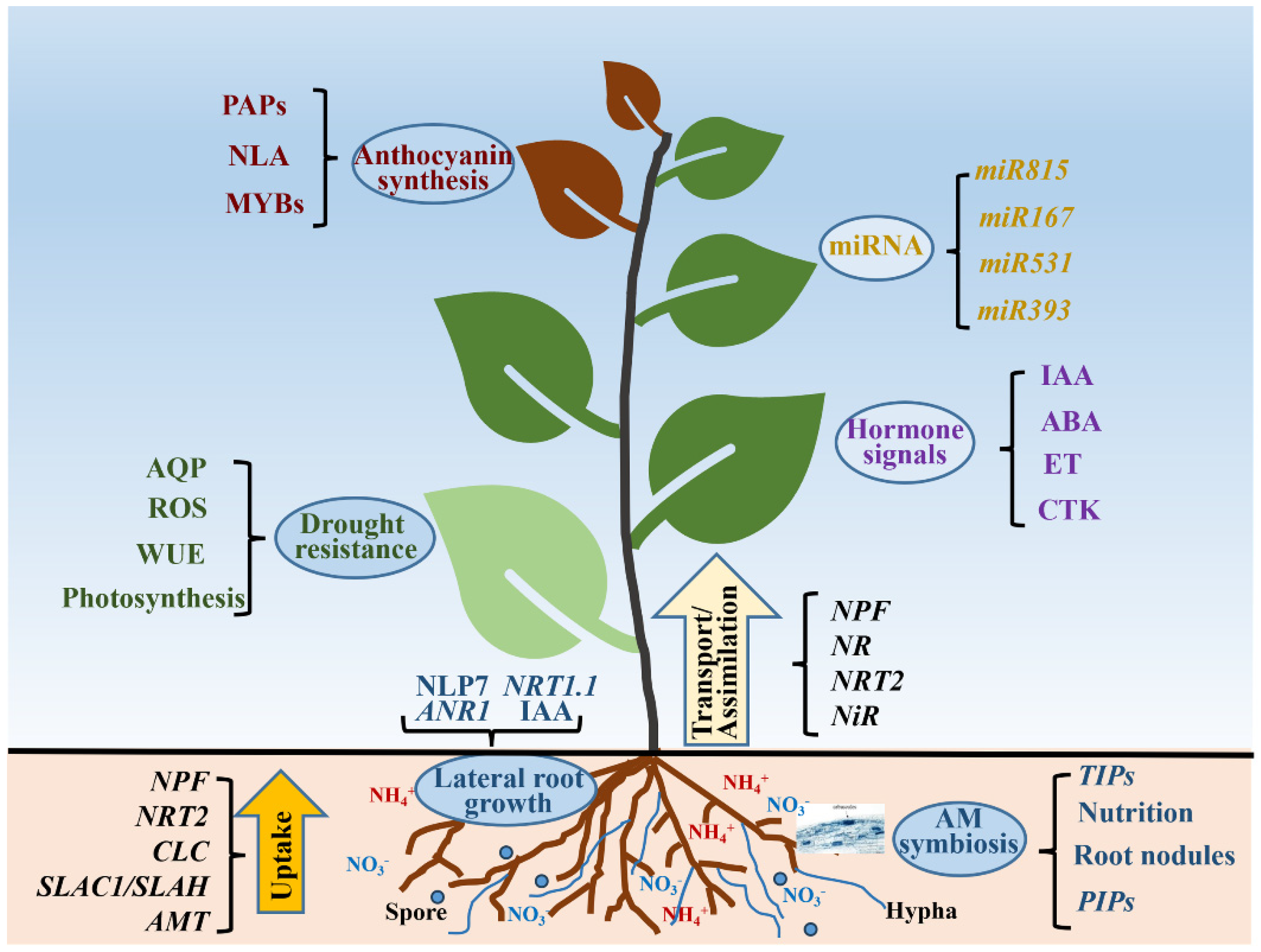
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).