1. Introduction
Taxus chinensis var.
mairei is a unique Tertiary relict tree species in China that is known as the “living fossil of the plant kingdom”. It has attracted much attention because it is rare and endangered, and it is used for ornamental, timber, and medicinal purposes.
T. chinensis has a beautiful crown, hard wood, and red heartwood, which can be processed into high-quality furniture. Its leaves, branches, stem, and bark are rich in paclitaxel and docetaxel, which have therapeutic effects in the treatment of ovarian cancer, breast cancer, lung cancer, hepatocellular carcinoma, and other cancers and difficult diseases [
1]. This species has attracted much attention since it was included in the list of national Class I key wild plants for protection by China’s State Forestry Administration in 1999, and it was included in the Appendix II list by the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) in 2004. Currently,
T. chinensis is mainly found in the hilly areas of Jiangnan, south of the Yellow River [
2], such as Anhui, Zhejiang, Guangdong, Guangxi, etc. It is also found in India, Myanmar, Malaysia, Indonesia, and the Philippines [
3]. Because of its wide range of uses and high ornamental value, since the 1970s, it has frequently been subjected to large-scale logging and destruction. According to the survey, it is estimated that there were 1.35 × 10
4 plants in about 9 × 10
6 km
2 in 16 counties of five prefectures in the Hengduan Mountain area of western Yunnan, but tens of thousands of
T. chinensis have been peeled off and their leaves and branches have been chopped off in recent years [
4]. Excessive exploitation and use by humans, coupled with its low natural reproduction capacity, have led to a gradual decrease in its distribution area, and it is endangered in the wild. It is thus crucial to determine the potential area of suitable habitat
T. chinensis and its influencing factors.
Species distribution models (SDMs) are now widely used tools for understanding the potential distributions of species and the factors influencing them. They are based on the interrelationships between species distributions and environmental variables, and they are used to simulate the distribution of a species’ potentially suitable habitat and characterize that species’ preference for its environment through the probability of it existing in a given area [
5]. Commonly used SDMs include the Biological Climatic Model (BioCLIM) [
6], Maximum Entropy model (MaxEnt) [
7], Genetic Algorithm for Rule-set Production (GARP) [
8], Ecological Niche Factor Analysis (ENFA) [
9], Generalized Linear Models (GLM), and Random Forest (RF). Among these, MaxEnt is widely used. MaxEnt is a machine-learning approach that estimates a target probability distribution by calculating the probability distribution of maximum entropy. Compared to other methods, MaxEnt has the advantages of using presence-only occurrence data, the ability to use both continuous and categorical variables, effective control of model fit through certain parameter settings, and simplicity of operation and repeated runs to test model robustness [
10]. It has been shown to perform better than other SDMs due to its ability to maintain high prediction accuracy with small samples [
11], and it has thus been widely used in research considering the prediction of suitable habitats and conservation for species including the Qilian cypress [
12],
Codonopsis pilosula [
13],
Cunninghamia lanceolata [
14],
Thamnocalamus spathiflorus [
15], and
Hibiscus mutabilis [
16].
According to the IPCC’s Sixth Assessment Report on Climate Change, it is projected that global atmospheric warming will exceed 1.5 °C by the end of the 22nd century. As a core driver of vegetation change, the resulting dramatic fluctuations in climate will have profound impacts on the growth environment and physiological attributes of vegetation through temperature stress, water stress, phenological changes, and changes in sunshine duration and light intensity [
17]. To date, studies of
T. chinensis in China have mainly focused on resource distribution [
4,
18,
19,
20], breeding techniques [
21], genealogical structure [
22,
23], and chemical composition [
24,
25,
26], while few researchers have examined the changes in suitable habitat for
T. chinensis in the context of climate change; however, gaining a deeper understanding of this would help to improve the management and protection of this endangered species.
To this end, the work described in this paper used simulated future climate data corresponding to the three shared socioeconomic pathways, SSP1-2.6, SSP2-4.5, and SSP5-8.5, under the Beijing Climate Center Climate System Model-Medium Resolution of the Coupled Model Intercomparison Project Phase 6 (BCC-CSM2-MR of CMIP6). SSP1-2.6 represents a sustainable development pathway with low greenhouse gas (GHG) emissions and a temperature increase controlled below 2 °C by 2100; SSP2-4.5 represents a moderate development pathway with medium GHG emissions and a temperature increase controlled below 3 °C by 2100; and SSP5-8.5 represents the conventional development pathway with high GHG emissions and a temperature increase controlled below 5 °C by 2100 [
27].
This study explored the dominant factors affecting the geographic distribution of T. chinensis and simulated the spatial distribution of its current suitable habitat using a MaxEnt model based on data relating to climate, topography, soil, and other environmental characteristics. Additionally, using the future climate data outlined above, the evolution trends of suitable habitat for T. chinensis under the influence of climate change in the 2030s and 2050s were predicted with a view towards deepening the understanding of the theoretical mechanisms underlying potential changes in its distribution.
5. Conclusions
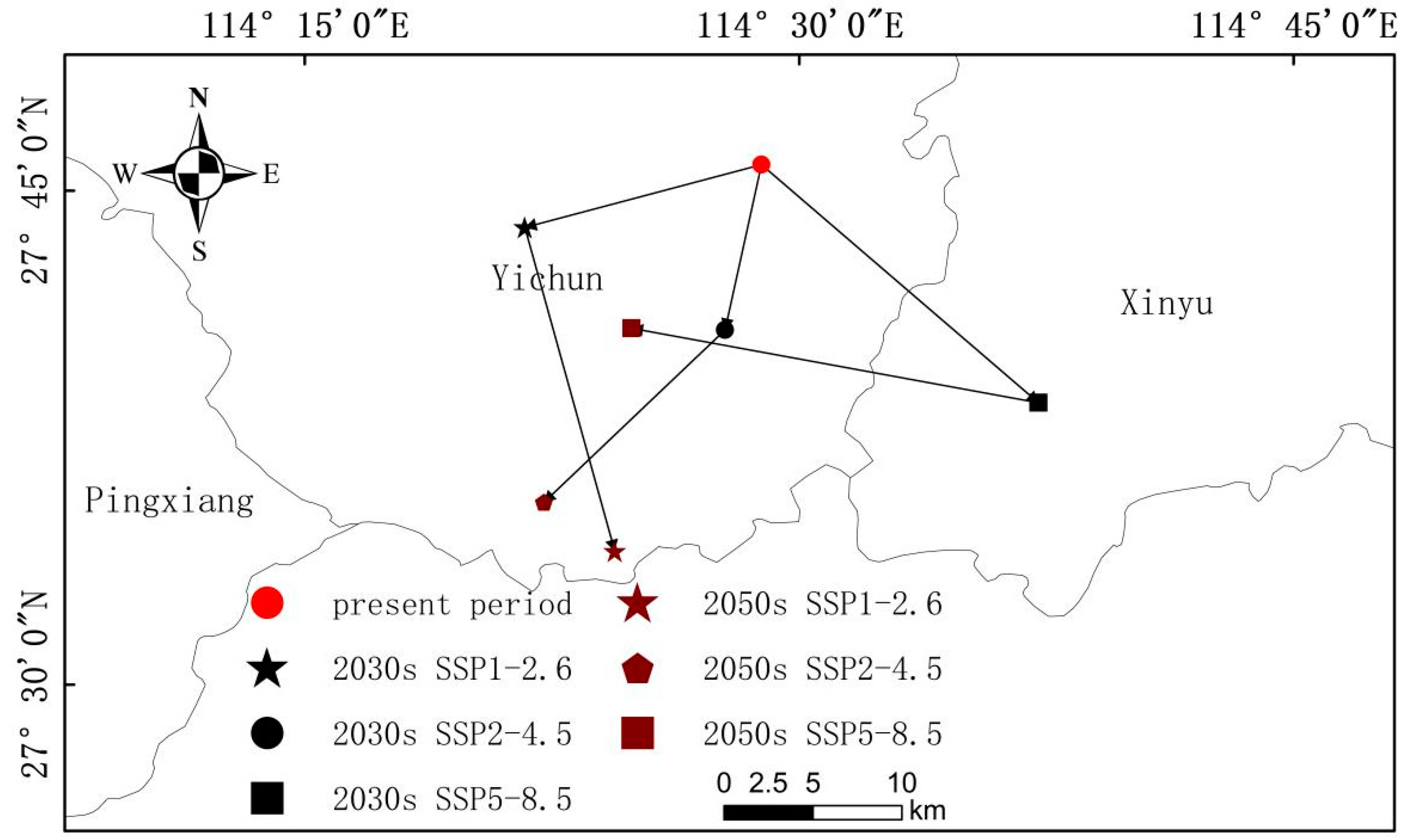
In this study, zones of potential ecological suitability for T. chinensis in the hilly area of Jiangnan and their influencing factors were modeled using meteorological data for three time periods in the present (1970–2000) and the future (2030s and 2050s) in combination with topographic and soil data. MaxEnt results show AUC = 0.848, the model prediction accuracy is high, and the prediction results are credible, from which the main conclusions can be summarized as follows.
The main environmental factors affecting the distribution of T. chinensis, in descending order, are Bio_18, Bio_15, Asp, Alt, Bio_5, and BS. According to the single-factor analysis, areas with medium altitude, a northeast or northwest slope orientation, a warmest quarterly precipitation between 508.3 and 629.2 mm, a maximum temperature in the warmest month between 34.6 and 35.9 °C, and relatively moist soil are most suitable for growing T. chinensis.
The results show that the area of suitable habitat for T. chinensis is 6.09 × 105 km2, and this is widely distributed across the whole study area. The area of high habitat suitability is 7.56 × 104 km2, and this is concentrated in northern Guangdong, northern Guangxi, southern Hubei, eastern Chongqing, southern Zhejiang, southern Jiangxi, western Fujian, and southern Hunan, and these are the key regions for in situ investigations.
With the warming of the climate, the original area of suitable habitat for T. chinensis remains unchanged, and the general trend is actually an increase.
Finally, although this study compared and screened 16 climate, topography, and soil factors for simulation, some more important environmental factors were still neglected. These include the intensity of human activities, biological invasion, and other factors. Follow-up studies can add these factors to further improve the accuracy and credibility of the predictions of the MaxEnt model.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}