
Range-Wide Assessment of Recent Longleaf Pine (Pinus palustris Mill.) Area and Regeneration Trends


Abstract
1. Introduction
2. Materials and Methods
2.1. Data
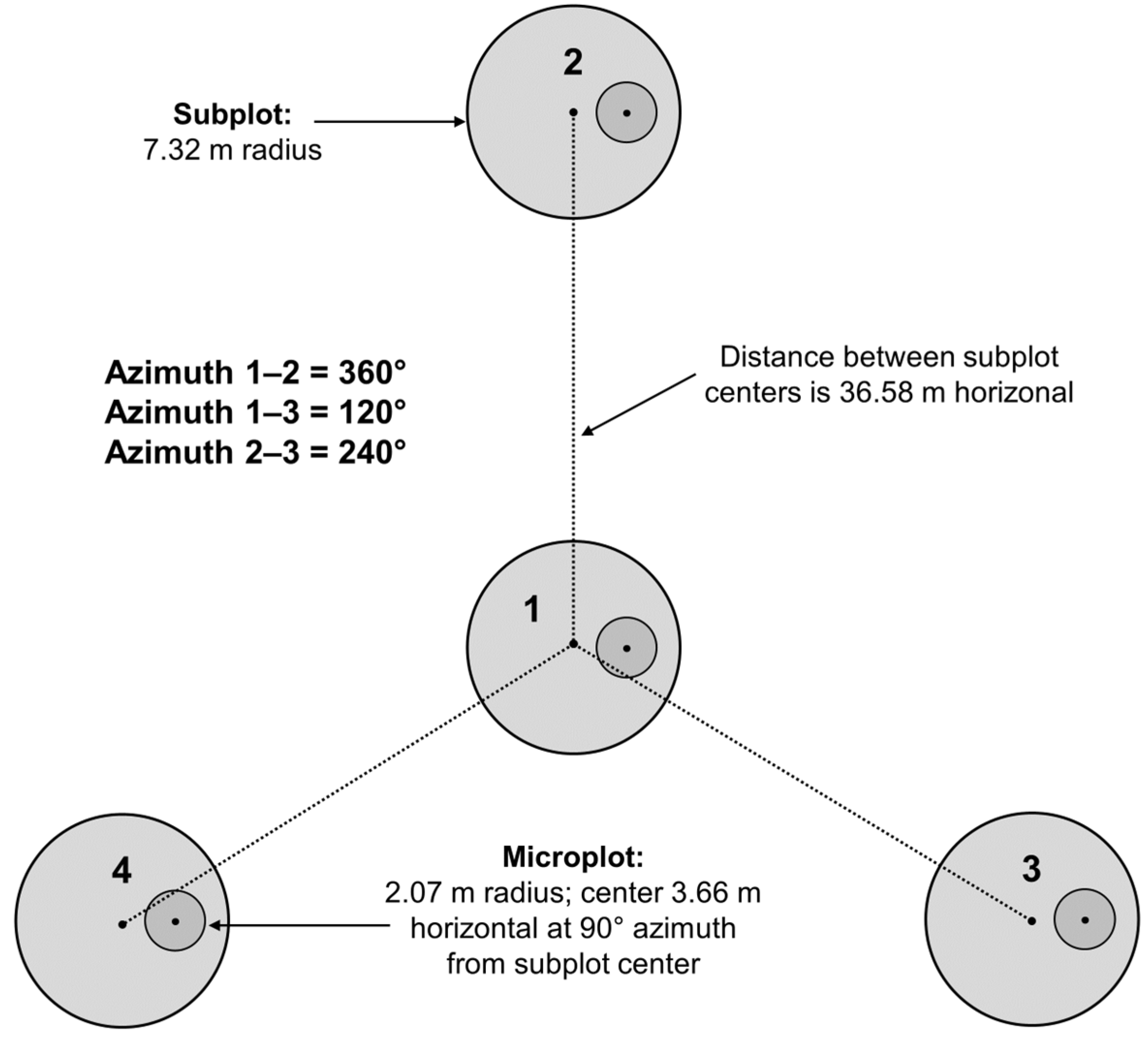
2.2. Count and Areal Estimates
2.3. Plot-Level Analyses
2.4. Regeneration within Seed Zones
3. Results
3.1. Count and Areal Estimates
3.2. Plot-Level Analyses
3.3. Regeneration within Seed Zones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frost, C. History and future of the longleaf pine ecosystem. In The Longleaf Ecosystem: Ecology, Silviculture and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer Science: New York, NY, USA, 2006; pp. 9–48. [Google Scholar]
- Stambaugh, M.C.; Varner, J.M.; Jackson, S.T. Biogeography: An interweave of climate, fire, and humans. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Bota Raton, FL, USA, 2017; pp. 17–38. [Google Scholar]
- Boyer, W.D. Longleaf pine. In Silvics of North America: 1. Conifers; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 1. [Google Scholar]
- Walker, J.; Peet, R.K. Composition and species diversity of pine-wiregrass savannas of the Green Swamp, North Carolina. Vegetatio 1984, 55, 163–179. [Google Scholar] [CrossRef]
- Carr, S.C.; Robertson, K.M.; Peet, R.K. A Vegetation Classification of Fire-Dependent Pinelands of Florida. Castanea 2010, 75, 153–189. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Carroll, W.D.; Kapeluck, P.R.; Johnson, R. History and restoration.of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Platt, W.J. Conservation and restoration of the Pinus palustris ecosystem. Appl. Veg. Sci. 2006, 9, 7–10. [Google Scholar] [CrossRef]
- Outcalt, K.W.; Sheffield, R.M. The Longleaf Pine Forest: Trends and Current Conditions; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 1996; p. 28. [Google Scholar]
- Noss, R.F.; LaRoe, E.T.; Scott, J.M. Endangered Ecosystems of the United States: A Preliminary Assessment of Loss and Degradation; U.S. Department of Interior, National Biological Service: Washington, DC, USA, 1995; p. 59. [Google Scholar]
- Farjon, A. Pinus palustris. IUCN Red List Threat. Species 2013, e.T39068A2886222. [Google Scholar] [CrossRef]
- Engstrom, R.T.; Sanders, F.J. Red-cockaded Woodpecker foraging ecology in an old-growth longleaf pine forest. Wilson Bull. 1997, 109, 203–217. [Google Scholar]
- Mitchell, R.; Engstrom, T.; Sharitz, R.R.; De Steven, D.; Hiers, K.; Cooper, R.; Kirkman, L.K. Old forests and endangered woodpeckers: Old-growth in the Southern coastal plain. Nat. Areas J. 2009, 29, 301–310. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Jack, S.B.; McIntyre, R.K. The fire forest of the past and present. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Bota Raton, FL, USA, 2017; pp. 3–16. [Google Scholar]
- America’s Longleaf Restoration Initiative. Range-Wide Conservation Plan for Longleaf Pine; America’s Longleaf Restoration Initiative: Atlanta, GA, USA, 2009; p. 42. [Google Scholar]
- America’s Longleaf Restoration Initiative. Range-Wide Conservation Plan for Longleaf Pine: 2025–2040; America’s Longleaf Restoration Initiative: Atlanta, GA, USA, 2023; p. 43. [Google Scholar]
- Oswalt, C.M.; Guldin, J.M. Status of Longleaf Pine in the South: An FIA Update; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2021; p. 25. [Google Scholar]
- Oswalt, C.M.; Cooper, J.A.; Brockway, D.G.; Brooks, H.W.; Walker, J.L.; Connor, K.F.; Oswalt, S.N.; Conner, R.C. History and Current Condition of Longleaf Pine in the Southern United States; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2012; p. 51. [Google Scholar]
- Oswalt, C.M.; Woodall, C.W.; Brooks, H.W. Is the footprint of longleaf pine in the Southeastern United States still shrinking? In Proceedings of the 17th Biennial Southern Silvicultural Research Conference, Shreveport, LA, USA, 5–7 March 2015; pp. 503–504. [Google Scholar]
- Platt, W.J.; Evans, G.W.; Rathbun, S.L. The population dynamics of a long-lived conifer (Pinus palustris). Am. Nat. 1988, 131, 491–525. [Google Scholar] [CrossRef]
- Ojha, S.K.; Tadesse, W.; Oswalt, C.M.; Gyawali, B. Occurrence, density, and distribution of longleaf pine regeneration in southeastern forests: An assessment by forest type, disturbance and site quality. For. Ecol. Manag. 2021, 481, 15. [Google Scholar] [CrossRef]
- Potter, K.M.; Riitters, K. A national multi-scale assessment of regeneration deficit as an indicator of potential risk of forest genetic variation loss. Forests 2021, 13, 19. [Google Scholar] [CrossRef]
- Manion, P.D.; Griffin, D.H. Large landscape scale analysis of tree death in the Adirondack Park, New York. For. Sci. 2001, 47, 542–549. [Google Scholar] [CrossRef]
- Burrill, E.A.; DiTommaso, A.M.; Turner, J.A.; Pugh, S.A.; Christiansen, G.; Perry, C.J.; Conkling, B.L. The Forest Inventory and Analysis Database: Database Description and User Guide Version 9.01 for Phase 2; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2021; p. 1026. [Google Scholar]
- Bechtold, W.A.; Scott, C.T. The Forest Inventory and Analysis plot design. In The Enhanced Forest Inventory and Analysis Program—National Sampling Design and Estimation Procedures; Bechtold, W.A., Patterson, P.L., Eds.; United States Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2005; Volume General Technical Report SRS-80; pp. 27–42. [Google Scholar]
- SAS Institute Inc. The SAS System for Windows, Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Smith, R.L.; Smith, T.M. Ecology and Field Biology, 6th ed.; Addison Wesley Longman: San Francisco, CA, USA, 2001; p. 771. [Google Scholar]
- Heath, L.S.; Hansen, M.; Smith, J.E.; Miles, P.D. Investigation into calculating tree biomass and carbon in the FIADB using a biomass expansion factor approach. In Proceedings of the Forest Inventory and Analysis (FIA) Symposium 2008, Park City, Utah, 21–23 October 2018. [Google Scholar]
- Woodall, C.W.; Heath, L.S.; Domke, G.M.; Nichols, M.C. Methods and Equations for Estimating Aboveground Volume, Biomass, and Carbon for Trees in the U.S. Forest Inventory, 2010; United States Department of Agriculture Forest Service, Northern Research Station: Newtown Square, PA, USA, 2011; p. 30. [Google Scholar]
- Pike, C.; Potter, K.M.; Berrang, P.; Crane, B.; Baggs, J.; Leites, L.; Luther, T. New seed-collection zones for the Eastern United States: The Eastern Seed Zone Forum. J. For. 2020, 118, 444–451. [Google Scholar] [CrossRef]
- Miller, S.A.; Bartow, A.; Gisler, M.; Ward, K.; Young, A.S.; Kaye, T.N. Can an ecoregion serve as a seed transfer zone? Evidence from a common garden study with five native species. Restor. Ecol. 2010, 19, 268–276. [Google Scholar] [CrossRef]
- Bower, A.D.; St Clair, B.; Erickson, V. Generalized provisional seed zones for native plants. Ecol. Appl. 2014, 24, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Cleland, D.T.; Freeouf, J.A.; Keys, J.E.; Nowacki, G.J.; Carpenter, C.A.; McNab, W.H. Ecological Subregions: Sections and Subsections for the conterminous United States. In General Technical Report WO-76; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2007. [Google Scholar]
- Janowiak, M.K.; Nagel, L.M.; Webster, C.R. Spatial scale and stand structure in northern hardwood forests: Implications for quantifying diameter distributions. For. Sci. 2008, 54, 497–506. [Google Scholar] [CrossRef]
- Rubin, B.D.; Manion, P.D.; Faber-Langendoen, D. Diameter distributions and structural sustainability in forests. For. Ecol. Manag. 2006, 222, 427–438. [Google Scholar] [CrossRef]
- Guldin, J.M. Restoration of Native Fire-Adapted Southern Pine-Dominated Forest Ecosystems: Diversifying the Tools in the Silvicultural Toolbox. For. Sci. 2019, 65, 508–518. [Google Scholar] [CrossRef]
- Ojha, S.K.; Dimov, L.D.; Tadesse, W. Growth, proportion, and distribution pattern of longleaf pine across southeastern forests and disturbance types: A change assessment for the period 1997–2018. PLoS ONE 2021, 16, 24. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Jack, S.B.; Engstrom, R.T. Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. For. Res.-Rev. Can. Rech. For. 2006, 36, 2724–2736. [Google Scholar] [CrossRef]
- Krugman, S.L.; Jenkinson, J.L. Pinus L. Pine. In The Woody Seed Plant Manual, Agricultural Handbook No. 727 ed.; Bonner, F.T., Karrfalt, R.P., Eds.; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2008; pp. 809–847. [Google Scholar]
- Croker, T.C., Jr.; Boyer, W.D. Regenerating Longleaf Pine Naturally; United States Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1975; p. 21. [Google Scholar]
- Boyer, W.D. Regenerating the natural longleaf pine forest. J. For. 1979, 77, 572–575. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W.; Boyer, W.D. Longleaf Pine Regeneration Ecology and Methods. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 95–133. [Google Scholar]
- Croker, T.C., Jr. Can the shelterwood method successfully regenerate longleaf pine? J. For. 1956, 54, 258–260. [Google Scholar]
- Carey, J.H. Pinus palustris. Available online: https://www.fs.usda.gov/database/feis/plants/tree/pinpal/all.html (accessed on 20 March 2024).
- Boyer, W.D.; Peterson, D.W. Longleaf pine. In Silvicultural Systems for the Major Forest Types of the United States; Burns, R.M., Ed.; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1983; Volume Agricultural Handbook 445; pp. 153–156. [Google Scholar]
- Curtin, P.J.; Knapp, B.O.; Jack, S.B.; Vickers, L.A.; Larsen, D.R.; Guldin, J.M. Effects of overstory competition on canopy recruitment patterns of naturally regenerated longleaf pine on two site types. Can. J. For. Res. 2020, 50, 624–635. [Google Scholar] [CrossRef]
- Guldin, J.M. Uneven-aged silviculture of longleaf pine. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer Science: New York, NY, USA, 2006; pp. 217–241. [Google Scholar]
- Kitchens, R.N. Alternative silvicultural systems on southern National Forests: A status report. In Proceedings of the Silvicultural Challenges and Opportunities in the 1990’s: Proceedings of the National Silvicultural Workshop, Petersburg, Alaska, 10–13 July 1989; pp. 46–50. [Google Scholar]
- Halpin, C.R.; Lorimer, C.G.; Hanson, J.J.; Palik, B.J. Predicted long-term effects of group selection on species composition and stand structure in northern hardwood forests. For. Ecol. Manag. 2017, 400, 677–691. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| First Evaluation | Second Evaluation | |||
|---|---|---|---|---|
| State | Years | Plots | Years | Plots |
| Alabama | 2006–2014 | 345 | 2015–2022 | 360 |
| Florida | 2009–2013 | 481 | 2014–2019 | 539 |
| Georgia | 2010–2014 | 300 | 2015–2021 | 326 |
| Louisiana | 2001–2010 | 133 | 2011–2019 | 91 |
| Mississippi | 2009–2015 | 184 | 2016–2021 | 289 |
| North Carolina | 2009–2015 | 141 | 2016–2021 | 152 |
| South Carolina | 2012–2016 | 246 | 2017–2021 | 229 |
| Texas | 2004–2014 | 38 | 2015–2021 | 54 |
| Virginia | 2012–2016 | 0 | 2017–2021 | 2 |
| Forest w. Longleaf Trees | Forest w. Longleaf Seedlings | Forest w. Any Longleaf | |||||||
| 2009–2015 | 2016–2021 | Diff. | 2009–2015 | 2016–2021 | Diff. | 2009–2015 | 2016–2021 | Diff. | |
| Total | 3,732,870 | 3,452,281 | (280,590) | 1,016,689 | 790,995 | (225,693) | 3,989,344 | 3,710,503 | (278,841) |
| Forest type | |||||||||
| Longleaf pine | 1,201,830 | 1,477,531 | 275,700 | 493,504 | 389,222 | (104,282) | 1,279,533 | 1,554,154 | 274,621 |
| Longleaf pine–oak | 358,715 | 290,847 | (67,868) | 155,551 | 104,673 | (50,877) | 387,048 | 327,579 | (59,468) |
| Sand pine | 41,589 | 35,039 | (6550) | 4056 | 6730 | 2673 | 45,645 | 36,891 | (8754) |
| Slash pine | 446,880 | 360,578 | (86,302) | 63,842 | 43,658 | (20,184) | 459,717 | 382,643 | (77,075) |
| Southern scrub oak | 150,330 | 87,927 | (62,402) | 54,219 | 42,860 | (11,358) | 160,551 | 100,488 | (60,063) |
| Other | 1,533,526 | 1,200,359 | (333,167) | 245,517 | 203,852 | (41,664) | 1,656,850 | 1,308,749 | (348,101) |
| State | |||||||||
| Alabama | 681,425 | 659,739 | (21,686) | 231,877 | 161,951 | (69,925) | 738,019 | 724,822 | (13,197) |
| Florida | 1,008,431 | 924,504 | (83,927) | 284,826 | 234,970 | (49,856) | 1,069,826 | 978,683 | (91,143) |
| Georgia | 584,017 | 593,689 | 9672 | 155,262 | 154,966 | (296) | 636,731 | 661,444 | 24,713 |
| Louisiana | 296,713 | 187,512 | (109,201) | 60,227 | 16,619 | (43,608) | 316,838 | 193,736 | (123,102) |
| Mississippi | 389,163 | 332,238 | (56,925) | 46,609 | 57,575 | 10,966 | 403,439 | 343,578 | (59,861) |
| North Carolina | 266,733 | 280,363 | 13,630 | 76,442 | 59,117 | (17,325) | 284,584 | 299,732 | 15,148 |
| South Carolina | 427,519 | 389,986 | (37,533) | 151,455 | 91,606 | (59,849) | 460,403 | 418,747 | (41,655) |
| Texas | 78,868 | 80,075 | 1207 | 9991 | 14,190 | 4199 | 79,504 | 85,588 | 6084 |
| Virginia | - | 4174 | 4174 | - | - | - | - | 4174 | 4174 |
| Ownership | |||||||||
| US Forest Service | 556,980 | 499,569 | (57,410) | 132,627 | 88,180 | (44,447) | 578,899 | 509,493 | (69,406) |
| Other federal | 325,055 | 365,413 | 40,358 | 121,692 | 114,445 | (7,247) | 354,075 | 379,125 | 25,051 |
| State/local | 314,843 | 347,940 | 33,097 | 98,953 | 83,973 | (14,980) | 324,162 | 371,174 | 47,013 |
| Private | 2,535,992 | 2,239,358 | (296,635) | 663,416 | 504,396 | (159,020) | 2,732,209 | 2,450,711 | (281,498) |
| Stand origin | |||||||||
| Planted | 372,417 | 733,348 | 360,931 | 265,707 | 298,007 | 32,300 | 495,985 | 887,675 | 391,690 |
| Natural | 3,360,453 | 2,718,932 | (641,521) | 750,982 | 492,988 | (257,994) | 3,493,359 | 2,822,829 | (670,530) |
| Longleaf Pine Trees | Longleaf Pine Seedlings | |||||
| 2009–2015 | 2016–2021 | Diff. | 2009–2015 | 2016–2021 | Diff. | |
| Total | 849,365,838 | 1,076,035,524 | 226,669,686 | 873,738,658 | 762,597,568 | (111,141,090) |
| Forest type | ||||||
| Longleaf pine | 587,763,975 | 838,628,811 | 250,864,836 | 513,204,378 | 466,011,442 | (47,192,936) |
| Longleaf pine–oak | 106,566,711 | 68,643,383 | (37,923,328) | 150,363,604 | 111,686,978 | (38,676,626) |
| Sand pine | 3,890,053 | 6,166,582 | 2,276,529 | 894,634 | 3,545,699 | 2,651,065 |
| Slash pine | 33,980,134 | 31,035,931 | (2,944,203) | 51,368,336 | 51,635,537 | 267,201 |
| Soutdern scrub oak | 13,900,984 | 10,055,168 | (3,845,816) | 35,120,160 | 29,846,280 | (5,273,880) |
| Otder | 103,263,982 | 121,505,650 | 18,241,668 | 122,787,546 | 99,871,632 | (22,915,914) |
| State | ||||||
| Alabama | 173,281,807 | 226,563,837 | 53,282,030 | 252,174,168 | 145,826,710 | (106,347,458) |
| Florida | 210,628,965 | 252,874,091 | 42,245,126 | 241,042,409 | 304,188,108 | 63,145,699 |
| Georgia | 128,540,953 | 175,635,129 | 47,094,176 | 94,323,502 | 114,678,563 | 20,355,061 |
| Louisiana | 46,201,822 | 30,459,653 | (15,742,169) | 37,495,017 | 8,931,903 | (28,563,114) |
| Mississippi | 60,265,210 | 71,499,713 | 11,234,503 | 21,044,587 | 52,919,285 | 31,874,698 |
| Nortd Carolina | 74,856,173 | 97,142,343 | 22,286,170 | 71,913,367 | 41,093,302 | (30,820,065) |
| Soutd Carolina | 143,780,505 | 197,722,995 | 53,942,490 | 139,051,212 | 87,416,243 | (51,634,969) |
| Texas | 12,138,545 | 20,074,562 | 7,936,017 | 16,694,396 | 7,543,454 | (9,150,942) |
| Virginia | - | 4,063,200 | 4,063,200 | - | - | - |
| Ownership | ||||||
| US Forest Service | 105,633,731 | 116,763,998 | 11,130,267 | 112,043,178 | 62,706,090 | (49,337,088) |
| Otder federal | 71,138,687 | 98,333,275 | 27,194,588 | 116,812,765 | 211,470,053 | 94,657,288 |
| State/local | 89,923,705 | 102,341,331 | 12,417,626 | 99,139,226 | 133,866,455 | 34,727,229 |
| Private | 582,669,715 | 758,596,919 | 175,927,204 | 545,743,488 | 354,554,970 | (191,188,518) |
| Stand origin | ||||||
| Planted | 224,744,876 | 494,323,030 | 269,578,154 | 192,371,057 | 234,457,884 | 42,086,827 |
| Natural | 624,620,963 | 581,712,493 | (42,908,470) | 681,367,600 | 528,139,684 | (153,227,916) |
| Importance Value (IV) | C Tonnes/ha | |||||||||
| Plots | 2009–2015 | 2016–2021 | 2009–2015 | 2016–2021 | ||||||
| 2009–2015 | 2016–2021 | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| All | 1754 | 1914 | 30.7 | 0.7 | 38.0 | 0.8 | 2.20 | 0.07 | 3.11 | 0.08 |
| Forest type | ||||||||||
| Longleaf pine | 520 | 745 | 62.4 | 1.3 | 65.9 | 1.1 | 4.38 | 0.17 | 5.26 | 0.16 |
| Longleaf pine–oak | 151 | 136 | 36.4 | 2.0 | 33.1 | 2.0 | 1.95 | 0.11 | 2.29 | 0.15 |
| Sand pine | 18 | 17 | 9.4 | 1.6 | 13.9 | 2.7 | 0.59 | 0.20 | 0.70 | 0.22 |
| Slash pine | 209 | 215 | 14.9 | 1.1 | 17.8 | 1.1 | 1.44 | 0.13 | 1.83 | 0.13 |
| Soutdern scrub oak | 65 | 45 | 22.0 | 2.8 | 22.4 | 3.5 | 0.63 | 0.09 | 0.98 | 0.17 |
| Otder | 791 | 756 | 14.0 | 0.7 | 18.5 | 0.8 | 1.17 | 0.06 | 1.68 | 0.09 |
| State | ||||||||||
| Alabama | 319 | 328 | 27.6 | 1.6 | 32.9 | 1.8 | 2.28 | 0.15 | 2.85 | 0.18 |
| Florida | 476 | 512 | 36.7 | 1.6 | 42.5 | 1.5 | 1.95 | 0.12 | 2.57 | 0.13 |
| Georgia | 275 | 292 | 31.4 | 1.9 | 39.6 | 2.1 | 1.96 | 0.14 | 2.51 | 0.17 |
| Louisiana | 130 | 89 | 26.9 | 2.7 | 31.4 | 3.4 | 3.08 | 0.37 | 4.19 | 0.53 |
| Mississippi | 188 | 281 | 19.6 | 1.8 | 33.3 | 1.9 | 2.01 | 0.23 | 4.11 | 0.26 |
| Nortd Carolina | 131 | 142 | 32.3 | 2.8 | 40.3 | 2.8 | 2.83 | 0.30 | 3.45 | 0.33 |
| Soutd Carolina | 198 | 216 | 33.9 | 2.2 | 42.6 | 2.4 | 2.22 | 0.23 | 3.58 | 0.26 |
| Texas | 37 | 52 | 19.1 | 3.8 | 26.1 | 4.0 | 1.95 | 0.44 | 3.31 | 0.66 |
| Virginia | 0 | 2 | - | 65.8 | 34.2 | - | 0.66 | 0.34 | ||
| Ownership group | ||||||||||
| US Forest Service | 248 | 477 | 37.8 | 2.2 | 41.3 | 1.5 | 3.63 | 0.26 | 4.62 | 0.21 |
| Otder federal | 144 | 162 | 40.2 | 2.7 | 44.5 | 2.6 | 3.20 | 0.28 | 3.37 | 0.28 |
| State/local | 144 | 157 | 41.2 | 2.9 | 41.2 | 2.7 | 2.48 | 0.21 | 2.92 | 0.26 |
| Private | 1218 | 1118 | 26.8 | 0.8 | 35.1 | 1.0 | 1.75 | 0.07 | 2.54 | 0.09 |
| Stand origin | ||||||||||
| Planted | 158 | 313 | 64.7 | 2.6 | 65.3 | 1.9 | 2.28 | 0.26 | 3.19 | 0.20 |
| Natural | 1596 | 1601 | 27.3 | 0.7 | 32.6 | 0.8 | 2.19 | 0.07 | 3.09 | 0.09 |
| Mean Plot-Level Change | ||
| IV | C/ha | |
| Latitude | 0.144 | 0.189 |
| Longitude | 0.089 | 0.039 |
| Elevation | 0.183 | 0.103 |
| Slope | 0.070 | 0.135 |
| Aspect (nortdness) | 0.055 | 0.077 |
| Aspect (eastness) | −0.039 | 0.125 |
| Stand age | −0.120 | 0.124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potter, K.M.; Oswalt, C.M.; Guldin, J.M. Range-Wide Assessment of Recent Longleaf Pine (Pinus palustris Mill.) Area and Regeneration Trends. Forests 2024, 15, 1255. https://doi.org/10.3390/f15071255
Potter KM, Oswalt CM, Guldin JM. Range-Wide Assessment of Recent Longleaf Pine (Pinus palustris Mill.) Area and Regeneration Trends. Forests. 2024; 15(7):1255. https://doi.org/10.3390/f15071255
Chicago/Turabian StylePotter, Kevin M., Christopher M. Oswalt, and James M. Guldin. 2024. "Range-Wide Assessment of Recent Longleaf Pine (Pinus palustris Mill.) Area and Regeneration Trends" Forests 15, no. 7: 1255. https://doi.org/10.3390/f15071255
APA StylePotter, K. M., Oswalt, C. M., & Guldin, J. M. (2024). Range-Wide Assessment of Recent Longleaf Pine (Pinus palustris Mill.) Area and Regeneration Trends. Forests, 15(7), 1255. https://doi.org/10.3390/f15071255