
The Distribution Range of Populus euphratica Oliv. (Salicaceae) Will Decrease Under Future Climate Change in Northwestern China
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Distribution Records of P. euphratica
2.2. Environmental Variables
2.3. Model Optimization and Accuracy Evaluation
2.4. Analysis of the Potential Suitable Area
3. Results
3.1. Contributions of Environmental Variables
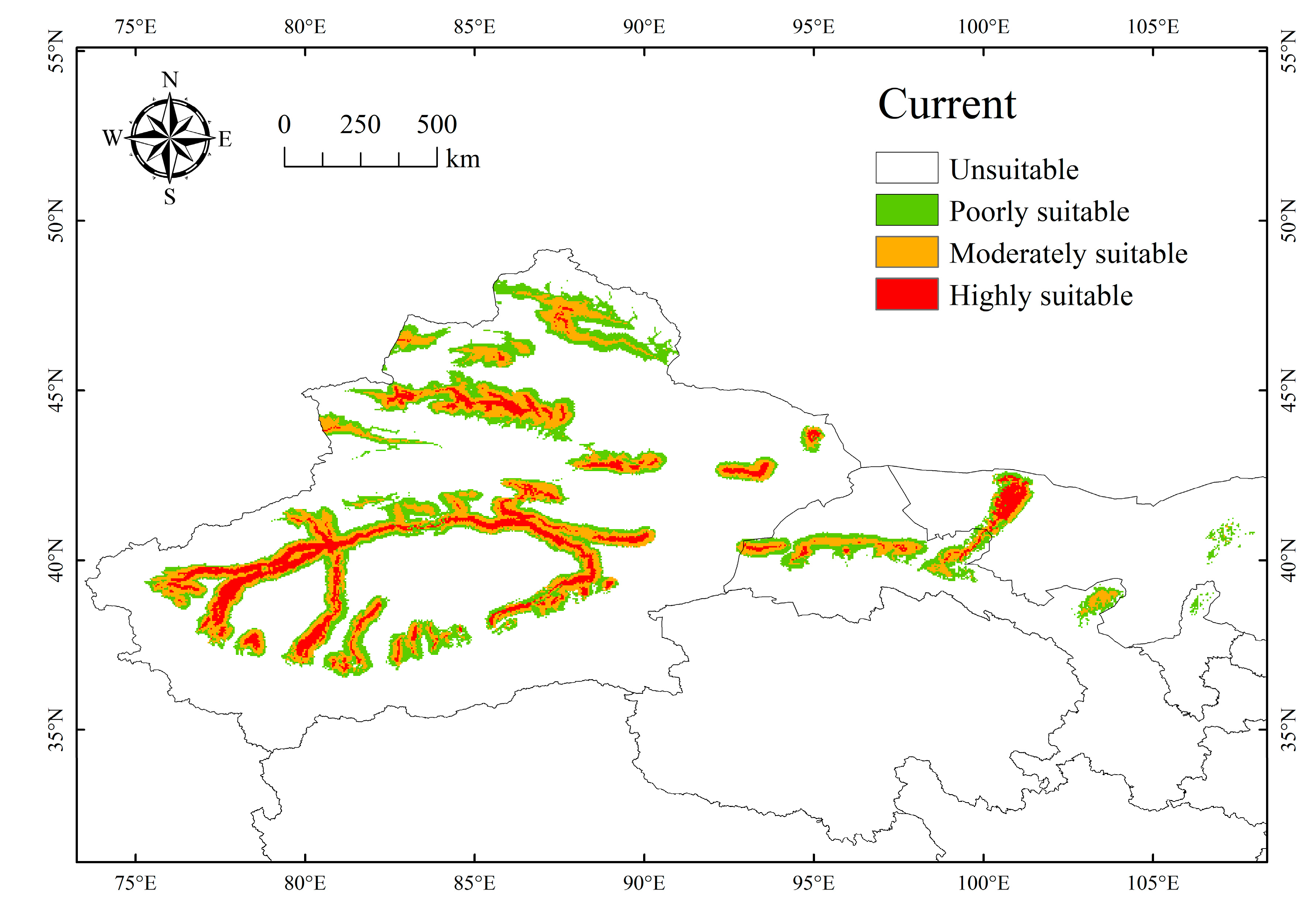
3.2. Current Distribution of the Suitable Habitat for P. euphratica
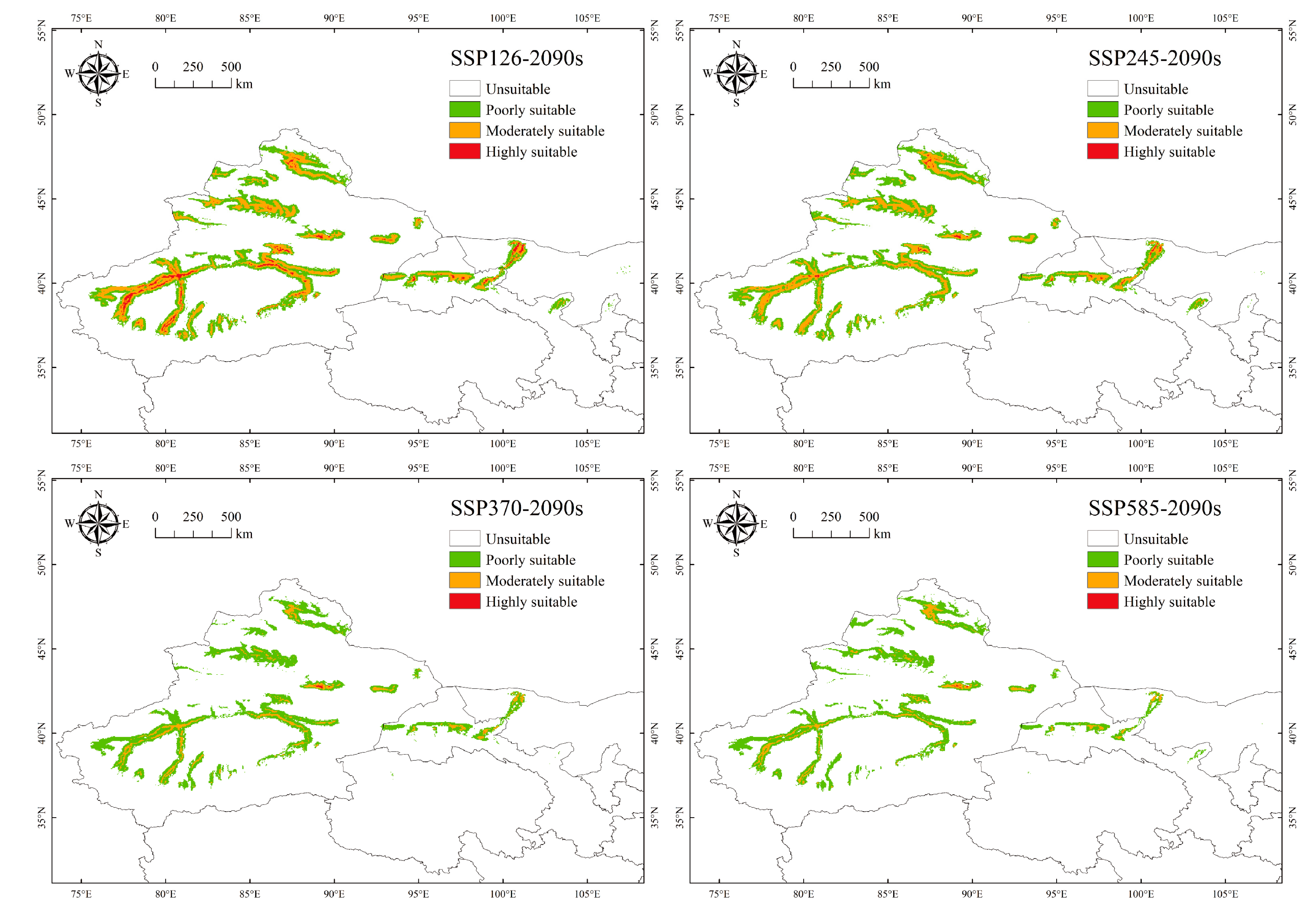
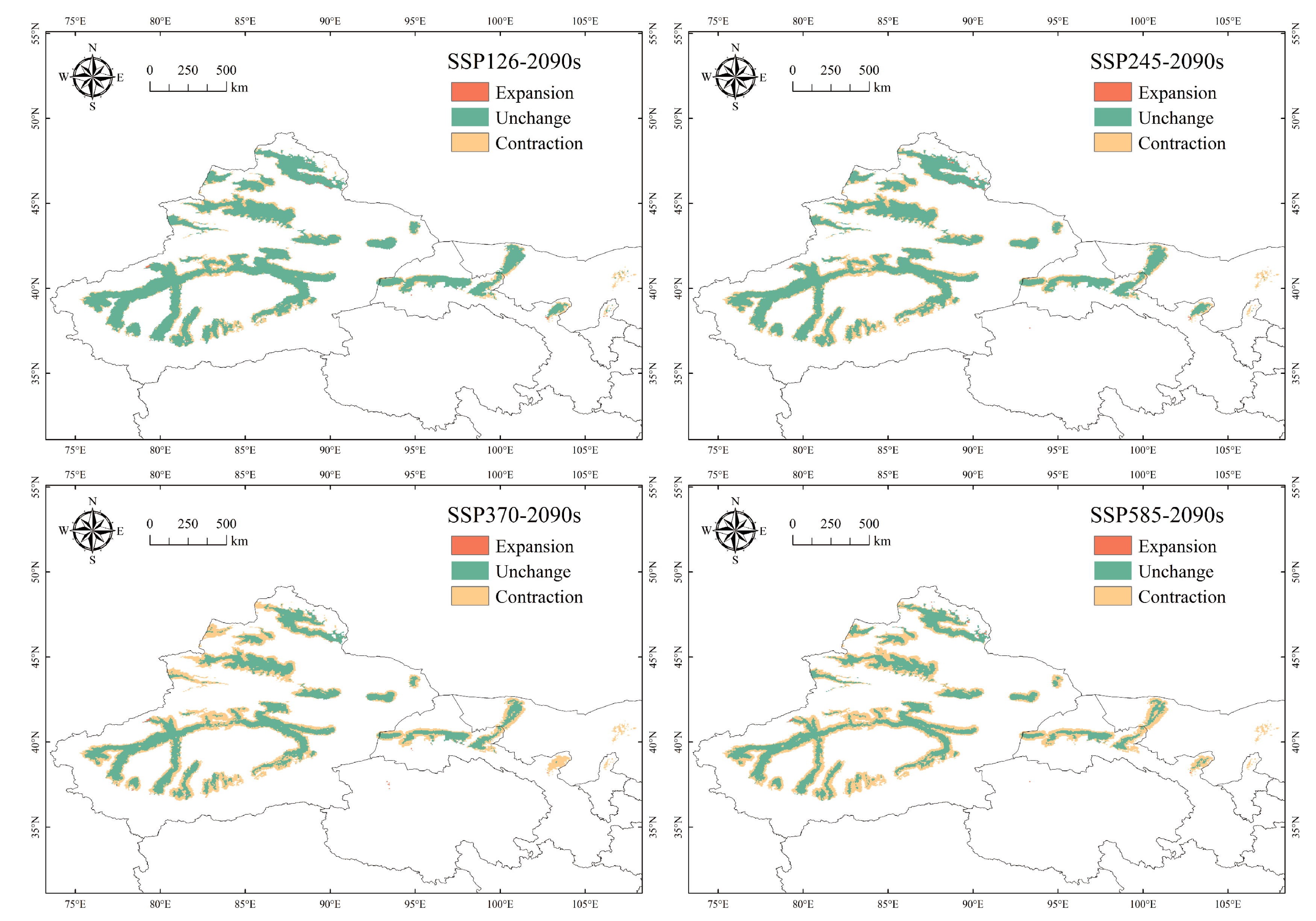
3.3. Changes in the Potential Suitable Habitat for P. euphratica under Future Climate Change Scenarios
4. Discussion
4.1. Key Predictor Variables Affecting the Distribution of P. euphratica
4.2. Prospective Changes in Suitable Habitat Distribution
4.3. Recommendations for Conservation and Limitations of the Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Scheffers, B.R.; Meester, L.D.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The Broad Footprint of Climate Change from Genes to Biomes to People. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef]
- Manthey, M.; Box, E.O. Realized Climatic Niches of Deciduous Trees: Comparing Western Eurasia and Eastern North America. J. Biogeogr. 2007, 34, 1028–1040. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; pp. 1–34. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavergne, S.; Roquet, C.; Boulangeat, I.; Lafourcade, B.; Araujo, M.B. Consequences of Climate Change on the Tree of Life in Europe. Nature 2011, 470, 531–534. [Google Scholar] [CrossRef]
- Lian, X.; Jiao, L.; Hu, Y.; Liu, Z. Future Climate Imposes Pressure on Vulnerable Ecological Regions in China. Sci. Total Environ. 2023, 858, 159995. [Google Scholar] [CrossRef]
- Ling, H.; Zhang, P.; Xu, H.; Zhao, X. How to Regenerate and Protect Desert Riparian Populus euphratica Forest in Arid Areas. Sci. Rep. 2015, 5, 15418. [Google Scholar] [CrossRef]
- Fang, Z.; Zhao, S.; Skvortsov, A.K. Salicaceae. In Flora of China; Science Press Beijing & Missouri Botanical GardenPress: Beijing, China; St. Louis, MO, USA, 1999; Volume 4, pp. 139–274. [Google Scholar]
- Wang, S. The status, conservation and recovery of global resources of Populus euphratica. World For. Res. 1996, 9, 38–45. (In Chinese) [Google Scholar] [CrossRef]
- Keram, A.; Halik, Ü.; Keyimu, M.; Aishan, T.; Mamat, Z.; Rouzi, A. Gap Dynamics of Natural Populus euphratica Floodplain Forests Affected by Hydrological Alteration along the Tarim River: Implications for Restoration of the Riparian Forests. For. Ecol. Manage. 2019, 438, 103–113. [Google Scholar] [CrossRef]
- Shi, H.; Shi, Q.; Zhou, X.; Imin, B.; Li, H.; Zhang, W.; Kahaer, Y. Effect of the Competition Mechanism of between Co-Dominant Species on the Ecological Characteristics of Populus euphratica under a Water Gradient in a Desert Oasis. Glob. Ecol. Conserv. 2021, 27, e01611. [Google Scholar] [CrossRef]
- Peng, Y.; He, G.; Wang, G. Spatial-Temporal Analysis of the Changes in Populus euphratica Distribution in the Tarim National Nature Reserve over the Past 60 Years. Int. J. Appl. Earth Obs. Geoinf. 2022, 113, 103000. [Google Scholar] [CrossRef]
- Qi, G.; Hu, Y.; Owens, J.R.; Dai, Q.; Hou, R.; Yang, Z.; Qi, D. Habitat Suitability for Chiru (Pantholops hodgsonii): Implications for Conservation Management across the Tibetan Region of Chang Tang. J. Wildl. Manage. 2015, 79, 384–392. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting Species Distributions for Conservation Decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Li, X.; Wu, K.; Hao, S.; Kang, L.; Ma, J.; Zhao, R.; Zhang, Y. Mapping of Suitable Habitats for Earthworms in China. Soil Biol. Biochem. 2023, 184, 109081. [Google Scholar] [CrossRef]
- Ma, Q.; Li, X.; Wu, S.; Zeng, F. Potential Geographical Distribution of Stipa purpurea across the Tibetan Plateau in China under Climate Change in the 21st Century. Glob. Ecol. Conserv. 2022, 35, e02064. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Feng, J.; Chen, C.; Chen, J.; Long, T.; Li, J.; Zang, R.; Li, J. Differential Responses to Climate and Land-Use Changes in Threatened Chinese Taxus Species. Forests 2019, 10, 766. [Google Scholar] [CrossRef]
- Cassini, M.H. Ecological Principles of Species Distribution Models: The Habitat Matching Rule. J. Biogeogr. 2011, 38, 2057–2065. [Google Scholar] [CrossRef]
- Yang, J.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R. Potential Geographical Distribution of the Edangred Plant Isoetes under Human Activities Using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The First Species Distribution Modelling Package, Its Early Applications and Relevance to Most Current MaxEnt Studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Carpenter, G.; Gillison, A.N.; Winter, J. DOMAIN: A Flexible Modelling Procedure for Mapping Potential Distributions of Plants and Animals. Biodivers. Conserv. 1993, 2, 667–680. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Yackulic, C.B.; Chandler, R.; Zipkin, E.F.; Royle, J.A.; Nichols, J.D.; Campbell Grant, E.H.; Veran, S. Presence-only Modelling Using MAXENT : When Can We Trust the Inferences? Methods Ecol. Evol. 2013, 4, 236–243. [Google Scholar] [CrossRef]
- He, P.; Li, J.; Li, Y.; Xu, N.; Gao, Y.; Guo, L.; Huo, T.; Peng, C.; Meng, F. Habitat Protection and Planning for Three Ephedra Using the MaxEnt and Marxan Models. Ecol. Indic. 2021, 133, 108399. [Google Scholar] [CrossRef]
- Xu, W.; Du, Q.; Yan, S.; Cao, Y.; Liu, X.; Guan, D.-X.; Ma, L.Q. Geographical Distribution of As-Hyperaccumulator Pteris Vittata in China: Environmental Factors and Climate Changes. Sci. Total Environ. 2022, 803, 149864. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, J.; Zhang, L.; Chen, S.; Zhao, A.; Ning, X.; Fan, G.; Wu, N.; Zhang, L.; Wang, Z. Prediction of the Potentially Suitable Areas of Litsea Cubeba in China Based on Future Climate Change Using the Optimized MaxEnt Model. Ecol. Indic. 2023, 148, 110093. [Google Scholar] [CrossRef]
- Xie, C.; Huang, B.; Jim, C.Y.; Han, W.; Liu, D. Predicting Differential Habitat Suitability of Rhodomyrtus tomentosa under Current and Future Climate Scenarios in China. For. Ecol. Manage. 2021, 501, 119696. [Google Scholar] [CrossRef]
- Shi, F.; Liu, S.; An, Y.; Sun, Y.; Zhao, S.; Liu, Y.; Li, M. Climatic Factors and Human Disturbance Influence Ungulate Species Distribution on the Qinghai-Tibet Plateau. Sci. Total Environ. 2023, 869, 161681. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Ning, Y.; Zheng, P.; Wang, R. The Effect of Climate Change on the Richness Distribution Pattern of Oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef]
- Zhang, P.; Deng, X.; Long, A.; Xu, H.; Ye, M.; Li, J. Change in Spatial Distribution Patterns and Regeneration of Populus euphratica under Different Surface Soil Salinity Conditions. Sci. Rep. 2019, 9, 9123. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, H.; Wang, H. Potential geographical distribution of populus euphratica in China under future climate change scenarios based on Maxent model. Acta Ecologica Sinica 2020, 40, 6552–6563. (In Chinese) [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H. Modeling the Distribution of Populus euphratica in the Heihe River Basin, an Inland River Basin in an Arid Region of China. Sci. China (Earth Sci.) 2018, 61, 1669–1684. [Google Scholar] [CrossRef]
- Liu, T.; Cao, J.; Qi, R.; Li, B.; Gao, B. Research of Potential Geographical Distribution of Picea purpurea Based on GIS and MaxEnt under Different Climate Conditions. Acta Bot. Boreali-Occident. Sin. 2022, 42, 481–491. (In Chinese) [Google Scholar]
- Ling, H.; Guo, B.; Xu, H.; Fu, J. Configuration of Water Resources for a Typical River Basin in an Arid Region of China Based on the Ecological Water Requirements (EWRs) of Desert Riparian Vegetation. Glob. Planet. Change 2014, 122, 292–304. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between Optimized MaxEnt and Random Forest Modeling in Predicting Potential Distribution: A Case Study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]
- Zhang, L.; Huettmann, F.; Liu, S.; Sun, P.; Yu, Z.; Zhang, X.; Mi, C. Classification and Regression with Random Forests as a Standard Method for Presence-Only Data SDMs: A Future Conservation Example Using China Tree Species. Ecol. Inform. 2019, 52, 46–56. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Yang, X.-Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent Modeling for Predicting the Potential Distribution of Medicinal Plant, Justicia Adhatoda L. in Lesser Himalayan Foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Bowen, A.K.M.; Stevens, M.H.H. Temperature, Topography, Soil Characteristics, and NDVI Drive Habitat Preferences of a Shade-tolerant Invasive Grass. Ecol. Evol. 2020, 10, 10785–10797. [Google Scholar] [CrossRef]
- Shen, Y.; Tu, Z.; Zhang, Y.; Zhong, W.; Xia, H.; Hao, Z.; Zhang, C.; Li, H. Predicting the Impact of Climate Change on the Distribution of Two Relict Liriodendron Species by Coupling the MaxEnt Model and Actual Physiological Indicators in Relation to Stress Tolerance. J. Environ. Manage. 2022, 322, 116024. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Ren, G.; Zhao, K.; Wang, X. Global Potential Distribution Prediction of Xanthium italicum Based on Maxent Model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the Maxent Model’s Complexity on the Prediction of Species Potential Distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q. Comparison of Akaike Information Criterion (AIC) and Bayesian Information Criterion (BIC) in Selection of Stock–Recruitment Relationships. Fish Res. 2006, 77, 220–225. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Vanagas, G. Receiver Operating Characteristic Curves and Comparison of Cardiac Surgery Risk Stratification Systems. Interact Cardiovasc. Thorac. Surg. 2004, 3, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Swets, J.A. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Zhao, X.; Lei, M.; Wei, C.; Guo, X. Assessing the Suitable Regions and the Key Factors for Three Cd-Accumulating Plants (Sedum alfredii, Phytolacca americana, and Hylotelephium spectabile) in China Using MaxEnt Model. Sci. Total Environ. 2022, 852, 158202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent Modeling for Predicting the Potential Geographical Distribution of Two Peony Species under Climate Change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Han, X.; Deng, Y. Higher Precipitation Strengthens the Microbial Interactions in Semi-Arid Grassland Soils. Glob. Ecol. Biogeogr. 2018, 27, 570–580. [Google Scholar] [CrossRef]
- Chen, Y.; Li, W.; Zhou, H.; Chen, Y.; Hao, X.; Fu, A.; Ma, J. Experimental Study on Water Transport Observations of Desert Riparian Forests in the Lower Reaches of the Tarim River in China. Int. J. Biometeorol. 2017, 61, 1055–1062. [Google Scholar] [CrossRef]
- Li, M.; Ma, Z. Decadal Changes in Summer Precipitation over Arid Northwest China and Associated Atmospheric Circulations. Int. J. Climatol. 2018, 38, 4496–4508. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Chen, Y.; LI, Y.; Ren, Z. Response of Populus euphratica at Different Ages to Rainfall Pulses in the Desert Riparian Forest of the Lower Reaches of the Heihe River. Arid Zone Research 2016, 33, 172–178. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, K.; Lu, L.; Zhou, S.; Zhang, H. Seedling recruitment in desert riparian forest following river flooding in the middle reaches of the Tarim River. Acta Ecol. Sin. 2011, 31, 3322–3329. (In Chinese) [Google Scholar]
- Wang, Y.; Wang, J.; Qu, M.; Li, J. Plant community assembly processes and key drivers in an arid inland river basin. Biodivers. Sci. 2022, 30, 5–20. (In Chinese) [Google Scholar] [CrossRef]
- Schöb, C.; Armas, C.; Guler, M.; Prieto, I.; Pugnaire, F.I. Variability in Functional Traits Mediates Plant Interactions along Stress Gradients. J. Ecol. 2013, 101, 753–762. [Google Scholar] [CrossRef]
- Zhao, L.; Xie, C.; Liu, X.; Wang, N.; Yu, Z.; Dong, X.; Wang, L. Water Sources of Major Plant Species along a Strong Climatic Gradient in the Inland Heihe River Basin. Plant Soil 2020, 455, 439–466. [Google Scholar] [CrossRef]
- Zhou, H.H.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Photosynthesis of Populus euphratica in Relation to Groundwater Depths and High Temperature in Arid Environment, Northwest China. Photosynthetica 2010, 48, 257–268. [Google Scholar] [CrossRef]
- Yao, Y.; Zheng, C.; Liu, J.; Cao, G.; Xiao, H.; Li, H.; Li, W. Conceptual and Numerical Models for Groundwater Flow in an Arid Inland River Basin. Hydrol. Process. 2015, 29, 1480–1492. [Google Scholar] [CrossRef]
- Hua, P. Studies on Seed Germination and Seeding Early Growth of Populus euphratica on The Flood Plain. Xinjiang Environ. Prot. 2003, 25, 14–17. (In Chinese) [Google Scholar] [CrossRef]
- Taub, D.R.; Seemann, J.R.; Coleman, J.S. Growth in Elevated CO2 Protects Photosynthesis against High-Temperature Damage. Plant Cell Environ. 2000, 23, 649–656. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, Y.; Li, W.; Chen, Y. Photosynthesis of Populus euphratica olive and its response to CO2 concentration and high temperature in arid environment. Acta Ecol. Sin. 2009, 29, 2797–2810. (In Chinese) [Google Scholar] [CrossRef]
- Berry, J.; Bjorkman, O. Photosynthetic Response and Adaptation to Temperature in Higher Plants. Ann. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Mahmoodi, S.; Ahmadi, K.; Heydari, M.; Karami, O.; Esmailzadeh, O.; Heung, B. Elevational Shift of Endangered European Yew under Climate Change in Hyrcanian Mountain Forests: Rethinking Conservation-Restoration Strategies and Management. For. Ecol. Manage. 2023, 529, 120693. [Google Scholar] [CrossRef]
- Ni, J. Impacts of Climate Change on Chinese Ecosystems: Key Vulnerable Regions and Potential Thresholds. Reg. Envir. Chang. 2011, 11, 49–64. [Google Scholar] [CrossRef]
- Shi, J.; Xia, M.; He, G.; Gonzalez, N.C.T.; Zhou, S.; Lan, K.; Ouyang, L.; Shen, X.; Jiang, X.; Cao, F.; et al. Predicting Quercus Gilva Distribution Dynamics and Its Response to Climate Change Induced by GHGs Emission through MaxEnt Modeling. J. Environ. Manage. 2024, 357, 120841. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ma, J.; Chen, G. Potential Geographical Distribution and Its Multi-Factor Analysis of Pinus massoniana in China Based on the Maxent Model. Ecol. Indic. 2023, 154, 110790. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, J.; Leng, G.; Zhao, J.; Wang, L.; Ma, W. Future Potential Distribution and Expansion Trends of Highland Barley under Climate Change in the Qinghai-Tibet Plateau (QTP). Ecol. Indic. 2022, 136, 108702. [Google Scholar] [CrossRef]
- Yang, W.; Sun, S.; Wang, N.; Fan, P.; You, C.; Wang, R.; Zheng, P.; Wang, H. Dynamics of the Distribution of Invasive Alien Plants (Asteraceae) in China under Climate Change. Sci. Total Environ. 2023, 903, 166260. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Tian, C.; Ma, Q.; Lu, G.; Zeng, F. Variation characteristics of potential distribution patterns of Alhagi sparsifolia Shap. under climate change and human disturbance. Acta Ecol. Sin. 2022, 42, 7349–7361. (In Chinese) [Google Scholar]
- Zhu, N. Predicting the Geographic Distribution of Calligonum mongolicum under Climate Change. J. Desert Res. 2019, 39, 136–144. (In Chinese) [Google Scholar]
- Xiao, F.; Liu, Q.; Qin, Y. Predicting the Potential Distribution of Haloxylon ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model. Biology 2024, 13, 3. [Google Scholar] [CrossRef]
- Su, B.; Huang, J.; Fischer, T.; Wang, Y.; Kundzewicz, Z.W.; Zhai, J.; Sun, H.; Wang, A.; Zeng, X.; Wang, G.; et al. Drought Losses in China Might Double between the 1.5 °C and 2.0 °C Warming. Proc. Natl. Acad. Sci. USA 2018, 115, 10600–10605. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, Y.; Zhu, C.; Li, Z.; Fang, G.; Li, Y.; Fu, A. Climate Change May Accelerate the Decline of Desert Riparian Forest in the Lower Tarim River, Northwestern China: Evidence from Tree-Rings of Populus euphratica. Ecol. Indic. 2020, 111, 105997. [Google Scholar] [CrossRef]
- Aishan, T.; Betz, F.; Halik, Ü.; Cyffka, B.; Rouzi, A. Biomass Carbon Sequestration Potential by Riparian Forest in the Tarim River Watershed, Northwest China: Implication for the Mitigation of Climate Change Impact. Forests 2018, 9, 196. [Google Scholar] [CrossRef]
- Lei, S.; Wang, W.; Wang, Y.; Chen, L.; Wu, X.; Deng, Z.; Zhang, T.; Du, Z.; Li, J. Effects of Tamarix ramosissima seedlings on Populus euphratica seedlings under various water and salinity conditions. Acta Ecol. Sin. 2020, 40, 7638–7647. (In Chinese) [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Variables | Description | Units |
|---|---|---|---|
| Bioclimatic variables | Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range | °C | |
| Bio3 | Isothermality ((BIO2/BIO7) (*100)) | / | |
| Bio4 | Temperature seasonality | / | |
| Bio5 | Max temperature of the warmest month | °C | |
| Bio6 | Min temperature of the coldest month | °C | |
| Bio7 | Temperature annual range | °C | |
| Bio8 | Mean temperature of the wettest quarter | °C | |
| Bio9 | Mean temperature of the driest quarter | °C | |
| Bio10 | Mean temperature of the warmest quarter | °C | |
| Bio11 | Mean temperature of the coldest quarter | °C | |
| Bio12 | Annual precipitation | mm | |
| Bio13 | Precipitation of the wettest month | mm | |
| Bio14 | Precipitation of the driest month | mm | |
| Bio15 | Precipitation seasonality | / | |
| Bio16 | Precipitation of the wettest quarter | mm | |
| Bio17 | Precipitation of the driest quarter | mm | |
| Bio18 | Precipitation of the warmest quarter | mm | |
| Bio19 | Precipitation of the coldest quarter | mm | |
| Topography variables | Elev | Elevation | m |
| Asp | Aspect | ° | |
| Slope | Slope | ° | |
| Water sources | Disriver | Distance from river | m |
| Variable | Percent Contribution/% | Permutation Importance/% |
|---|---|---|
| Bio13 | 44.1 | 70.8 |
| Disriver | 23.3 | 20.4 |
| Bio4 | 13.9 | 0.8 |
| Bio5 | 11.6 | 0.6 |
| Bio15 | 3.1 | 0.9 |
| Asp | 1.4 | 0.7 |
| Slope | 1.2 | 0.7 |
| Elev | 1.1 | 4.9 |
| Bio2 | 0.2 | 0 |
| Bio6 | 0.1 | 0.1 |
| Bio14 | 0 | 0 |
| Suitable Classes | Current | 2090s | |||
|---|---|---|---|---|---|
| SSP126 | SSP245 | SSP370 | SSP585 | ||
| Poorly suitable | 21.23 | 23.93 | 24.12 | 22.52 | 21.15 |
| +12.7% | +13.57% | +6.05% | −0.39% | ||
| Moderately suitable | 19.16 | 13.99 | 11.19 | 3.85 | 3.24 |
| −27.01% | −41.62% | −79.91% | −83.1% | ||
| Highly suitable | 8.74 | 1.92 | 0.43 | 0.06 | 0.04 |
| −78.08% | −95.04% | −99.31% | −99.56% | ||
| Total | 49.14 | 39.84 | 35.74 | 26.43 | 24.43 |
| −18.93% | −27.28% | −46.22% | −50.29% |
| Current | 2090s | ||||
|---|---|---|---|---|---|
| SSP126 | SSP245 | SSP370 | SSP585 | ||
| Disriver | 10,973.28 | 9526.39 | 8604.12 | 7373.02 | 6282.92 |
| Elev | 954.46 | 936.79 | 926.35 | 910.46 | 896.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, X.; Qu, M.; Wang, J.; Hou, J.; Wang, Y.; Li, G.; Luo, M.; Li, Z.; Li, J. The Distribution Range of Populus euphratica Oliv. (Salicaceae) Will Decrease Under Future Climate Change in Northwestern China. Forests 2024, 15, 1288. https://doi.org/10.3390/f15081288
Lei X, Qu M, Wang J, Hou J, Wang Y, Li G, Luo M, Li Z, Li J. The Distribution Range of Populus euphratica Oliv. (Salicaceae) Will Decrease Under Future Climate Change in Northwestern China. Forests. 2024; 15(8):1288. https://doi.org/10.3390/f15081288
Chicago/Turabian StyleLei, Xun, Mengjun Qu, Jianming Wang, Jihua Hou, Yin Wang, Guanjun Li, Meiwen Luo, Zhijun Li, and Jingwen Li. 2024. "The Distribution Range of Populus euphratica Oliv. (Salicaceae) Will Decrease Under Future Climate Change in Northwestern China" Forests 15, no. 8: 1288. https://doi.org/10.3390/f15081288