
Predicting Potential Suitable Areas of Dendrocalamus brandisii under Global Climate Change
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection and Processing
2.2. Acquisition and Selection of Environmental Data
2.3. Model Optimization
2.4. Model Construction and Accuracy Assessment
2.5. Changes in Spatial Patterns and Centroid Shift of D. brandisii’s Suitable Areas
3. Results
3.1. Model Optimization Results and Accuracy Evaluation
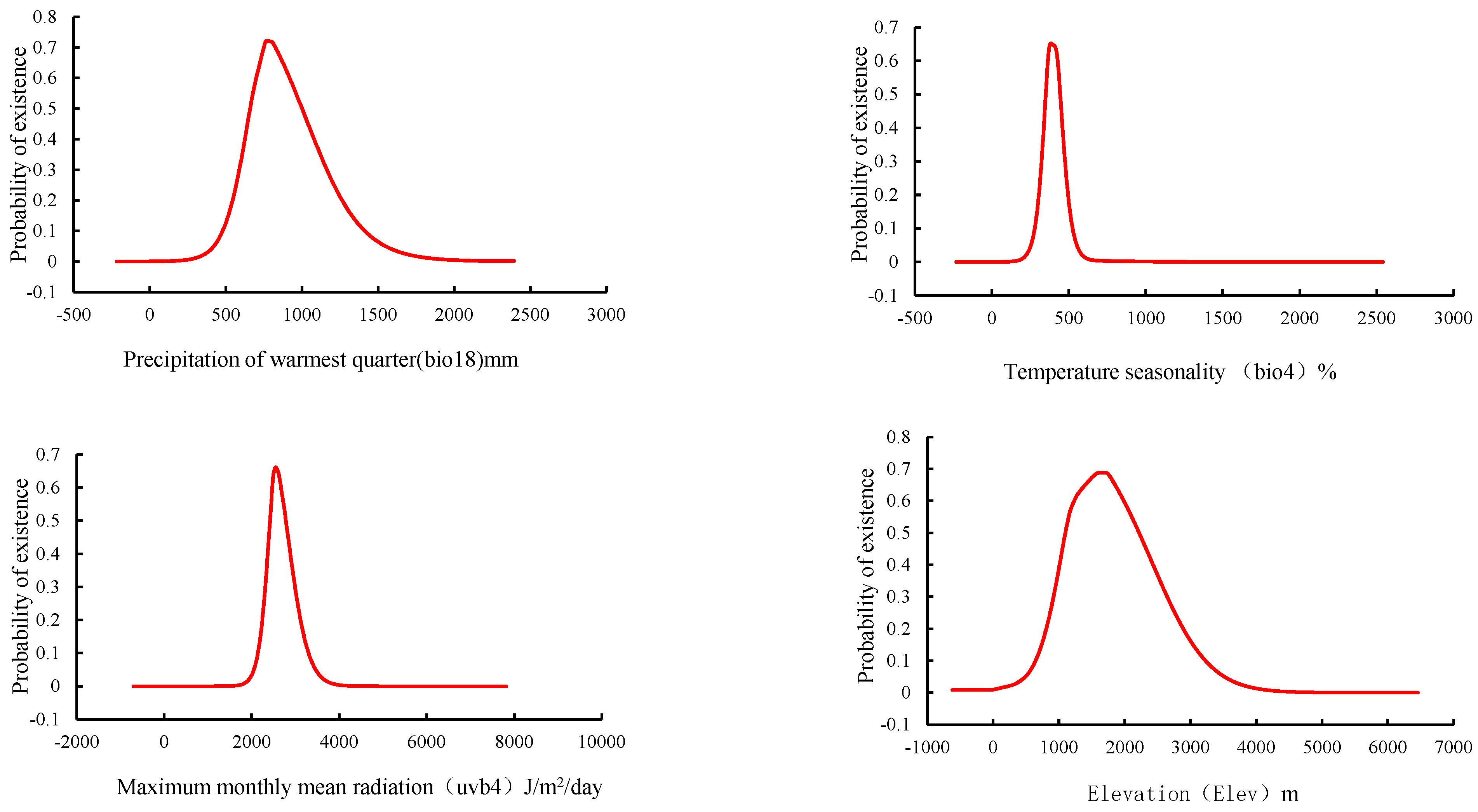
3.2. Key Factors Affecting the Potential Distribution of D. brandisii
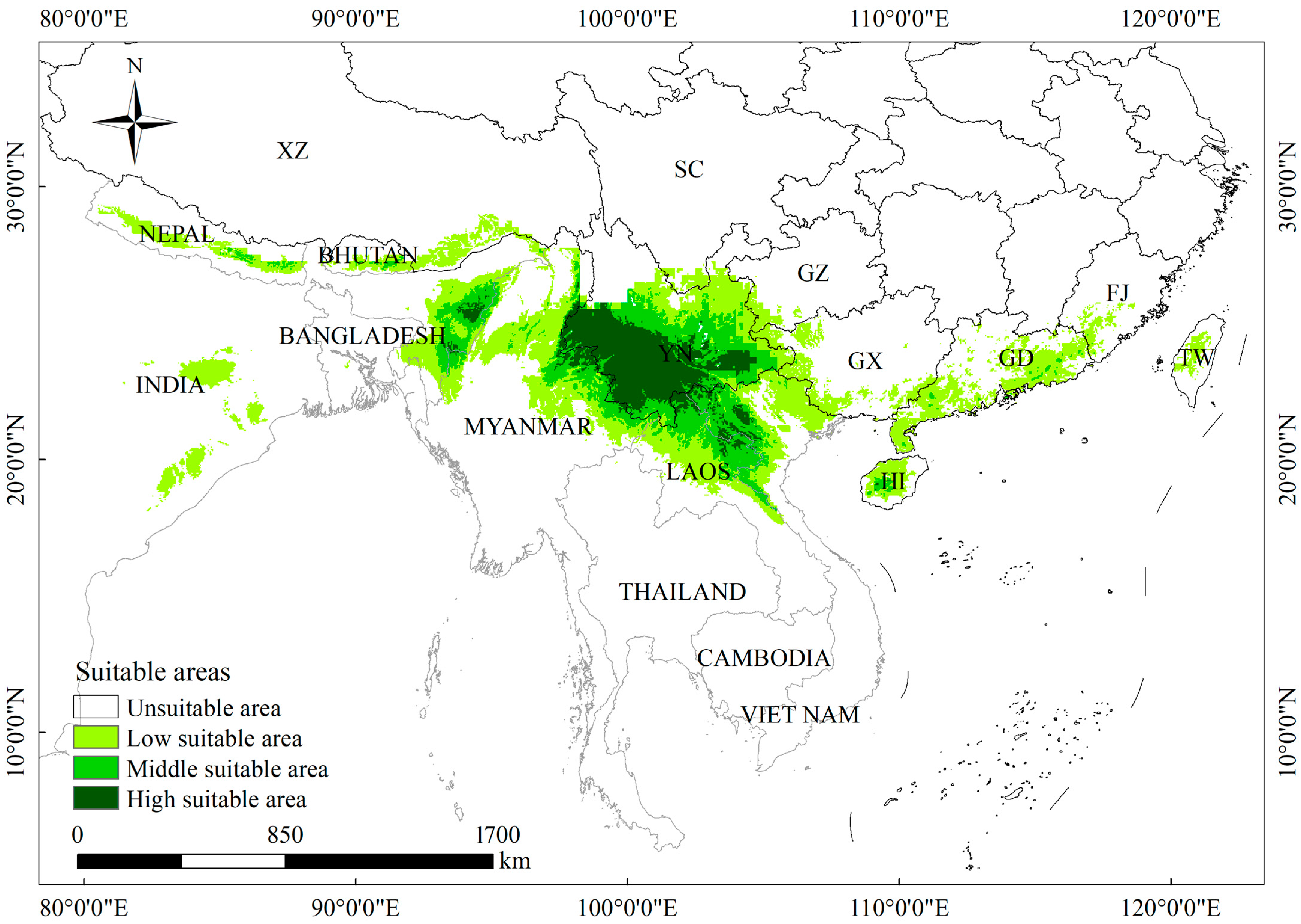
3.3. Current Climate Potential Distribution of D. brandisii
3.4. Simulation of Future Potential Suitable Areas for D. brandisii
3.5. Changes in the Spatial Pattern of D. brandisii’s Potential Suitable Habitat
3.6. Shift of Suitable Habitat Centroid
4. Discussion
4.1. Evaluation of Prediction Results
4.2. Impact of Environmental Variables on the Habitability Zone of D. brandisii
4.3. Spatial Distribution Pattern of the Potential Habitability Zone of D. brandisii
4.4. Conservation and Cultivation of D. brandisii
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Menezes-Silva, P.E.; Loram-Lourenço, L.; Alves, R.D.F.B.; Sousa, L.F.; da Silva Almeida, S.E.; Farnese, F.S. Different ways to die in a changing world: Consequences of climate change for tree species performance and survival through an ecophysiological perspective. Ecol. Evol. 2019, 9, 11979–11999. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. (Eds.) The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-B.; Svenning, J.-C.; Chen, G.-K.; Zhang, M.-G.; Huang, J.-H.; Chen, B.; Ordonez, A.; Ma, K.-P. Human activities have opposing effects on distributions of narrow-ranged and widespread plant species in China. Proc. Natl. Acad. Sci. USA 2019, 116, 26674–26681. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Yadav, M.; Mathur, A. Bamboo as a sustainable material in the construction industry: An overview. Mater. Today Proc. 2021, 43, 2872–2876. [Google Scholar] [CrossRef]
- Gupta, A.; Kumar, A. Potential of bamboo in sustainable development. Asia Pac. Bus. Rev. 2008, 4, 100–107. [Google Scholar] [CrossRef]
- Sawarkar, A.D.; Shrimankar, D.D.; Kumar, A.; Kumar, A.; Singh, E.; Singh, L.; Kumar, S.; Kumar, R. Commercial clustering of sustainable bamboo species in India. Ind. Crops Prod. 2020, 154, 112693. [Google Scholar] [CrossRef]
- Zhang, W.J.; Liu, W.Y.; Zhu, L.Y.; Tu, D.D.; Zhu, S.H.; Zhao, X.T.; Hui, C.M. Effects of Different Cultivation Modes of Dendrocalamus brandisii. World Bamboo Ratt. 2022, 20, 36–42. [Google Scholar]
- Wang, Y.; Zhan, H.; Ding, Y.; Wang, S.; Lin, S. Variability of anatomical and chemical properties with age and height in Dendrocalamus brandisii. BioResources 2016, 11, 1202–1213. [Google Scholar] [CrossRef]
- Liu, W.Y.; Hui, C.M.; Zou, X.M.; Wang, Y. Effects of Soil Temperature and Water Content on the Early Germination and Yield Improvement of Dendrocalamus brandisii Bamboo Shoots. J. Bamboo Res. 2019, 38, 39–48. [Google Scholar] [CrossRef]
- Hua, M.; Chen, J.; Bi, W.; Kong, J.J.; Hu, Y.L.; LI, Y.Q.; Yang, Y.M.; Wang, J. Determination of Total Flavonoids and Vitexin, Isovitexin, Orientin, Isoorientin in Cluster Bamboo by High-performance Liquid Chromatography. J. West China For. Sci. 2018, 47, 86–90. [Google Scholar] [CrossRef]
- Zhu, S.H.; Zhao, X.T.; Hui, C.M.; Zhang, Z.F.; Zhang, R.L.; Su, W.H.; Liu, W.Y. Effects of different planting durations of Dendrocalamus brandisii on the soil bacterial community. J. Soils Sediments 2023, 23, 3891–3902. [Google Scholar] [CrossRef]
- Meng, L.Y.; Huang, X.F.; Huang, D.Y.; Tang, W.; Pan, Y.G.; Deng, L.M.; Yu, H.L.; Wei, Y.N. The bamboo shoot yield and correlation analysis of six large clustered bamboos after afforestation. J. Cent. South Univ. For. Technol. 2022, 42, 46–54. [Google Scholar] [CrossRef]
- Zheng, S.Q. Research on Introduction and Planting Technology for Dendrocalamus brandisii. World Bamboo Ratt. 2016, 14, 18–21. [Google Scholar] [CrossRef]
- Zhao, X.T.; Hui, C.M.; Zhu, S.H.; Liu, W.Y.; Cai, C.L.; Zhang, W.J.; Tu, D.D.; Zhu, L.Y. Response of Soil Organic Matter Content and Its Infrared Spectral Characteristics to Different Planting Durations of Dendrocalamus brandisii. Acta Agric. Univ. Jiangxiensis 2022, 44, 1448–1456. [Google Scholar] [CrossRef]
- Huang, L.; Chu, C.H.; Deng, L.; Zhan, H.; Wang, S.G. Leaf Anatomy and Photosynthetic Characteristics in Dendrocalamus brandisii Seedlings under Different Cultivations. J. West China For. Sci. 2020, 49, 68–74. [Google Scholar] [CrossRef]
- Pei, J.L.; Li, P.C.; Wang, Q.; Wang, S.G. Comparison of Bamboo Shoot Nutrients of Dendrocalamus brandisii among Different Provenances. J. Northwest For. Univ. 2018, 33, 156–161. [Google Scholar] [CrossRef]
- Yufan, D.; Qin, L.; Yihe, Z.; Jianxin, C. Effects of ultrasonic assisted high-temperature cooking method on the physicochemical structure characteristics and in vitro antioxidant capacities of dietary fiber from Dendrocalamus brandisii Munro shoots. Ultrason. Sonochem. 2023, 97, 106462. [Google Scholar] [CrossRef]
- Zhang, X.F.; Zhao, Y.H.; Li, Q. Effects of Three Biological Preservatives on the Lignification of Postharvest Dendrocalamus brandisii Bamboo Shoots. J. West China For. Sci. 2022, 51, 43–49. [Google Scholar] [CrossRef]
- Zhang, H.J.; Chen, Y.; Huang, J.L.; Lie, H.W. Predicting potential geographic distribution of Mikania micrantha planting based on ecological niche models in China. Trans. Chin. Soc. Agric. Eng. 2011, 27, 413–418. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Xu, J.; Cao, B.; Bai, C.K. Prediction of potential suitable distribution of endangered plant Kingdonia uniflora in China with MaxEnt. Chin. J. Ecol. 2015, 34, 3354–3359. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.Y.; Li, Z.H.; Ouyang, S.L.; Cheng, Y.; Zhou, Z.C.; Wu, J.Y. Prediction of the potential distribution of Cyclobalanopsis gilva in China based on the Maxent and ArcGIS model. J. Cent. South Univ. For. Technol. 2023, 43, 19–26. [Google Scholar] [CrossRef]
- Li, X.; Mao, F.; Du, H.; Zhou, G.; Xing, L.; Liu, T.; Han, N.; Liu, Y.; Zheng, J.; Dong, L. Spatiotemporal evolution and impacts of climate change on bamboo distribution in China. J. Environ. Manag. 2019, 248, 109265. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sanchez, E.; Mendoza-Gonzalez, G.; Rojas-Soto, O. Mexican priority bamboo species under scenarios of climate change. Bot. Sci. 2018, 96, 11–23. [Google Scholar] [CrossRef]
- Song, H.F.; Dang, X.H.; Gao, Y.; Meng, Z.J.; Sun, Y.L. Potential Distribution of Pinus sylvestris in Inner Mongolia Autonomous Region Based on MaxEnt Model. J. Sichuan Agric. Univ. 2023, 41, 203–208. [Google Scholar] [CrossRef]
- Shi, X.; Wang, J.; Zhang, L.; Chen, S.; Zhao, A.; Ning, X.; Fan, G.; Wu, N.; Zhang, L.; Wang, Z. Prediction of the potentially suitable areas of Litsea cubeba in China based on future climate change using the optimized MaxEnt model. Ecol. Indic. 2023, 148, 110093. [Google Scholar] [CrossRef]
- Zhou, T.J.; Zou, L.W.; Chen, X.L. Commentary on the Coupled Model Intercomparison Project Phase 6 (CMIP6). Clim. Change Res. 2019, 15, 445–456. [Google Scholar] [CrossRef]
- Liu, J.Q.; Wei, G.K.; Shi, C.Q.; Zhao, T.N.; Qian, Y.K. Suitable distribution area of drought-resistant afforestation tree species in north China based on MaxEnt model. J. Beijing For. Univ. 2022, 44, 63–77. [Google Scholar] [CrossRef]
- Beckmann, M.; Václavík, T.; Manceur, A.M.; Šprtová, L.; von Wehrden, H.; Welk, E.; Cord, A.F. glUV: A global UV-B radiation data set for macroecological studies. Methods Ecol. Evol. 2014, 5, 372–383. [Google Scholar] [CrossRef]
- Chen, H.J.; Nie, Y.; Liu, X.H.; Liu, B.; Zhang, H. Research on prediction of potential suitable areas of Populus euphratica based on MaxEnt model. China Agric. Inform. 2021, 33, 46–55. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Xu, J.; Shrestha, K.K.; Kindt, R. Ensemble forecast of climate suitability for the Trans-Himalayan Nyctaginaceae species. Ecol. Model. 2014, 282, 18–24. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Z.; Zhang, M.Z.; Lai, W.F.; Yang, M.M.; Fan, H.H.; Zhang, G.F.; Chen, S.P.; Liu, B. Prediction of potential suitable distribution of Phoebe bournei based on MaxEnt optimization model. Acta Ecol. Sin. 2021, 41, 8135–8144. [Google Scholar] [CrossRef]
- Kong, W.Y.; Li, X.H.; Zou, H.F. Optimizing MaxEnt model in the prediction of species distribution. Chin. J. Appl. Ecol. 2019, 30, 2116–2128. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Alkishe, A.; Peterson, A.T. Climate change influences on the geographic distributional potential of the spotted fever vectors Amblyomma maculatum and Dermacentor andersoni. PeerJ 2022, 10, e13279. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.W.; Fang, Y.M. Responses of the distribution pattern of Quercus chenii to climate change following the Last Glacial Maximum. Chin. J. Plant Ecol. 2016, 40, 1164–1178. [Google Scholar] [CrossRef]
- Ning, Y.; Lei, J.R.; Song, X.Q.; Han, S.M.; Zhong, Y.F. Modeling the potential suitable habitat of Impatiens hainanensis, a limestone-endemic plant. Chin. J. Plant Ecol. 2018, 42, 946–954. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Liu, Y.L.; Qin, H.; Meng, Q.X. Prediction on spatial migration of suitable distribution of Elaeagnus mollis under climate change conditions in Shanxi Province, China. Chin. J. Appl. Ecol. 2019, 30, 496–502. [Google Scholar] [CrossRef]
- Pan, S.; Peng, D.-L.; Li, Y.-M.; Chen, Z.-J.; Zhai, Y.-Y.; Liu, C.; Hong, B. Potential global distribution of the guava root-knot nematode Meloidogyne enterolobii under different climate change scenarios using MaxEnt ecological niche modeling. J. Integr. Agric. 2023, 22, 2138–2150. [Google Scholar] [CrossRef]
- He, X.G.; Liu, H.; Zhang, J.; Cheng, W.; Ding, P.; Jia, F.M.; Li, Q.; Liu, C. Predicting potential suitable distribution areas for Juniperus przewalskii in QinghaiProvince of northwestern China based on the optimized MaxEnt model. J. Beijing For. Univ. 2023, 45, 19–31. [Google Scholar] [CrossRef]
- Porfirio, L.L.; Harris, R.M.B.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the Use of Species Distribution Models in Conservation Planning and Management under Climate Change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, B.; Liu, C.; Lai, J.; Zhang, J.; Ma, K. Identifying hotspots of endemic woody seed plant diversity in China. Divers. Distrib. 2012, 18, 673–688. [Google Scholar] [CrossRef]
- Dou, P.; Dong, Y.; Chen, L.; Yang, H.-Q. Modeling the potential distribution of different types of Dendrocalamus sinicus, the strongest woody bamboo in the world, with MaxEnt model. PeerJ 2022, 10, e13847. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Zheng, X.; Wang, Y.; Wang, W.; Li, M.; Wang, S.; Wang, J.; Wang, C.; Zhan, H. Effects of Climate Change and Environmental Factors on Bamboo (Ferrocalamus strictus), a PSESP Unique to China. Forests 2022, 13, 2108. [Google Scholar] [CrossRef]
- Kong, F.; Tang, L.; He, H.; Yang, F.; Tao, J.; Wang, W. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environ. Sci. Pollut. Res. 2021, 28, 34655–34663. [Google Scholar] [CrossRef]
- Lai, Y.Y.; Feng, J.M.; Yuan, Y.Y. The Influences of Climate Change on the Elevational Patterns of Tropical Plants in Nepal. J. Xinyang Norm. Univ. (Nat. Sci. Ed.) 2018, 31, 233–239. [Google Scholar] [CrossRef]
- Gao, Y.Q.; Cai, T.T.; Yang, Y.X.; Zhang, G.C.; Wan, X.L.; Liu, S.P.; Li, X.J.; Tie, X.J.; Huang, S.K.; Shi, D.H.; et al. Effects of Different Types of Mulch on Growth and Quality of Dendrocalamus brandisii Shoots. World Bamboo Ratt. 2023, 21, 45–49. [Google Scholar]
- Hui, C.M.; He, X.H.; Liu, W.Y.; Shi, Z.J.; Sun, M.S. Promoting the Development of Dendrocalamus brandisii Industry for Rural Vitalization. World Bamboo Ratt. 2022, 20, 66–69. [Google Scholar]
- Wu, Y.X.; Wang, J.L.; Han, B.M.; Yan, X.R. Forest classification for Motuo County: A method based on spatio-temporal-spectral characteristics. Remote Sens. Nat. Resour. 2023, 35, 180–188. [Google Scholar] [CrossRef]
- Enquist, B.J.; Feng, X.; Boyle, B.; Maitner, B.; Newman, E.A.; Jørgensen, P.M.; Roehrdanz, P.R.; Thiers, B.M.; Burger, J.R.; Corlett, R.T. The commonness of rarity: Global and future distribution of rarity across land plants. Sci. Adv. 2019, 5, eaaz0414. [Google Scholar] [CrossRef]
- Darabant, A.; Rai, P.B.; Staudhammer, C.L.; Dorji, T. Designing and evaluating bamboo harvesting methods for local needs: Integrating local ecological knowledge and science. Environ. Manag. 2016, 58, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.J.; Dong, S.K. Analyses on arbor distribution pattern and its environmental influence factors in the middle and lower reaches of Lancang River Basin. J. Plant Resour. Environ. 2018, 27, 65–71. [Google Scholar] [CrossRef]
- Wang, X.F.; Duan, Y.X.; Jin, L.L.; Wang, C.Y.; Peng, M.C.; Li, Y.; Wang, X.H.; Ma, Y.F. Prediction of historical, present and future distribution of Quercus sect. Heterobalanus based on the optimized MaxEnt model in China. Acta Ecol. Sin. 2023, 43, 6590–6604. [Google Scholar]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Bartlein, P.J.; Harrison, S.; Brewer, S.; Connor, S.; Davis, B.; Gajewski, K.; Guiot, J.; Harrison-Prentice, T.; Henderson, A.; Peyron, O. Pollen-based continental climate reconstructions at 6 and 21 ka: A global synthesis. Clim. Dyn. 2011, 37, 775–802. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Zhao, H.; Huang, Y.; Mou, Q.; Liu, K.; Chen, T.; Wang, X.; Zhang, Y.; Wei, B. Modeling habitat suitability of Houttuynia cordata Thunb (Ceercao) using MaxEnt under climate change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Ye, X.Z.; Li, J.H.; Liu, Y.P.; Chen, S.P.; Liu, B. Prediction of potential suitable area of Ulmus elongata in China under climate change scenario. Chin. J. Ecol. 2021, 40, 3822–3835. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Variable | Percent Contribution | Permutation Importance |
|---|---|---|---|
| 1 | Precipitation of warmest quarter (bio18) | 43.8 | 4.5 |
| 2 | Temperature seasonality (bio4) | 18.4 | 17.2 |
| 3 | Maximum monthly mean radiation (uvb-4) | 16.6 | 74.1 |
| 4 | Elevation (Elev) | 14.8 | 0.8 |
| 5 | Precipitation of coldest quarter (bio19) | 2.5 | 0.7 |
| 6 | Seasonal radiation (uvb-2) | 2.4 | 2 |
| 7 | Precipitation of wettest quarter (bio16) | 1.5 | 0.7 |
| Nation | Uninhabitable Area (×104 km2) | Low-Suitability Area (×104 km2) | Medium-Suitability Area (×104 km2) | High-Suitability Area (×104 km2) | Total Suitable Area (×104 km2) | The Proportion of High Suitability Area (%) | Total Suitable Area Proportion (%) |
|---|---|---|---|---|---|---|---|
| China | 918.89 | 23.94 | 8.36 | 11.84 | 44.14 | 78.33 | 47.88 |
| Bangladesh | 11.97 | 0.13 | 0.00 | 0.00 | 0.13 | 0.00 | 0.14 |
| Nepal | 10.64 | 2.63 | 0.28 | 0.00 | 2.91 | 0.00 | 3.16 |
| Laos | 11.36 | 4.42 | 3.21 | 0.70 | 8.33 | 4.62 | 9.03 |
| Burma | 42.72 | 10.62 | 3.74 | 1.08 | 15.44 | 7.14 | 16.75 |
| Thailand | 43.37 | 0.01 | 0.00 | 0.00 | 0.01 | 0.00 | 0.01 |
| Vietnam | 18.59 | 5.23 | 2.98 | 1.02 | 9.24 | 6.78 | 10.02 |
| India | 261.49 | 7.99 | 2.44 | 0.47 | 10.90 | 3.14 | 11.82 |
| Bhutan | 2.36 | 0.93 | 0.15 | 0.00 | 1.08 | 0.00 | 1.17 |
| Period | Current | 2050s (2041–2060) | 2070s (2061–2080) | 2090s (2081–2100) | |||
|---|---|---|---|---|---|---|---|
| SSPs2.6 | SSPs8.5 | SSPs2.6 | SSPs8.5 | SSPs2.6 | SSPs8.5 | ||
| Low-Suitability Area | 55.9 | 68.78 | 64.92 | 60.08 | 56.35 | 65.08 | 54.25 |
| Medium-Suitability Area | 21.16 | 19.81 | 18.81 | 18.69 | 20.19 | 17.70 | 19.75 |
| High-Suitability Area | 15.11 | 15.42 | 13.65 | 12.88 | 14.84 | 14.66 | 16.00 |
| Total Suitable Area | 92.17 | 104.01 | 97.38 | 91.65 | 91.38 | 97.44 | 90.00 |
| Period | Area/(×104 km2) | Change Rate/% | ||||||
|---|---|---|---|---|---|---|---|---|
| Increase | Reserved | Lost | Change | Increase | Reserved | Lost | Change | |
| 2050sSSPs2.6 | 16.50 | 87.51 | 4.66 | 11.84 | 17.90 | 94.94 | 5.06 | 12.85 |
| 2050sSSPs8.5 | 16.69 | 80.68 | 11.48 | 5.21 | 18.11 | 87.53 | 12.46 | 5.65 |
| 2070sSSPs2.6 | 12.91 | 78.74 | 13.43 | −0.52 | 14.00 | 85.43 | 14.57 | −0.56 |
| 2070sSSPs8.5 | 11.55 | 79.83 | 12.34 | −0.79 | 12.53 | 86.61 | 13.39 | −0.86 |
| 2090sSSPs2.6 | 18.47 | 78.97 | 13.20 | 5.27 | 20.04 | 85.68 | 14.32 | 5.72 |
| 2090sSSPs8.5 | 9.54 | 80.46 | 11.71 | −2.17 | 10.35 | 87.30 | 12.70 | −2.35 |
| Period | Current | 2041–2060s SSPs2.6 | 2041–2060s SSPs8.5 | 2061–2080s SSPs2.6 | 2061–2080s SSPs8.5 | 2081–2100s SSPs2.6 | 2081–2100s SSPs8.5 |
|---|---|---|---|---|---|---|---|
| longitude/(°) | 100.81 | 99.78 | 99.72 | 99.13 | 98.84 | 98.55 | 101.30 |
| latitude/(°) | 23.48 | 23.24 | 23.15 | 23.26 | 23.26 | 23.32 | 23.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, H.; Kingston, K.; Xu, Z.; Hosseini Bai, S.; Guo, L.; Liu, G.; Hui, C.; Liu, W. Predicting Potential Suitable Areas of Dendrocalamus brandisii under Global Climate Change. Forests 2024, 15, 1301. https://doi.org/10.3390/f15081301
Tao H, Kingston K, Xu Z, Hosseini Bai S, Guo L, Liu G, Hui C, Liu W. Predicting Potential Suitable Areas of Dendrocalamus brandisii under Global Climate Change. Forests. 2024; 15(8):1301. https://doi.org/10.3390/f15081301
Chicago/Turabian StyleTao, Hang, Kate Kingston, Zhihong Xu, Shahla Hosseini Bai, Lei Guo, Guanglu Liu, Chaomao Hui, and Weiyi Liu. 2024. "Predicting Potential Suitable Areas of Dendrocalamus brandisii under Global Climate Change" Forests 15, no. 8: 1301. https://doi.org/10.3390/f15081301