
Improved Branch Volume Prediction of Multi-Stemmed Shrubs: Implications in Shrub Volume Inventory and Fuel Characterization

 ,
,  ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Experimental Species
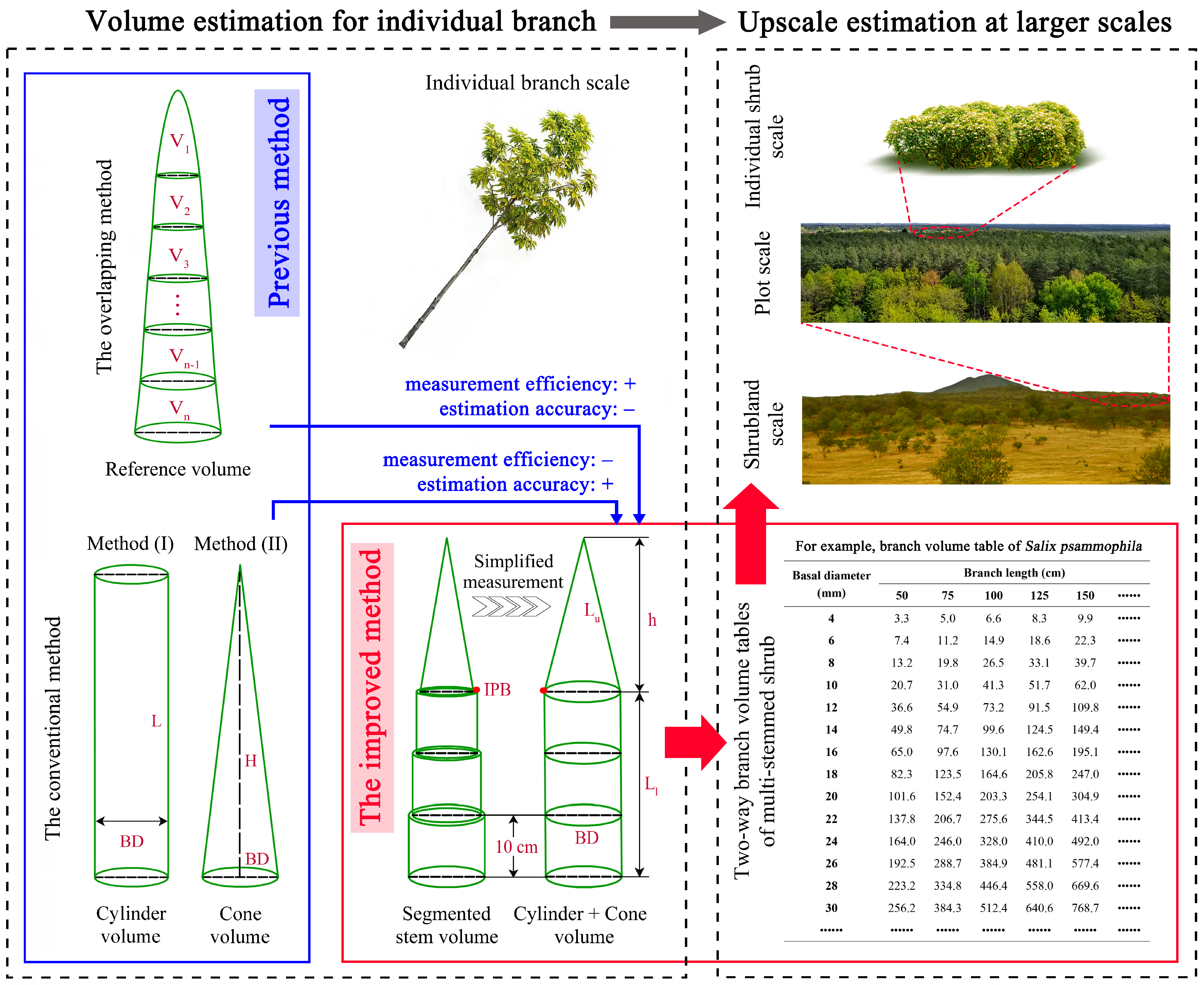
2.2. Methods of Estimating Branch Volume
2.2.1. Locating the Inflection Point of Branch Diameter
2.2.2. Predicting Branch Volume
2.3. Methods of Validating Branch Volume
2.4. Data Analysis
3. Results
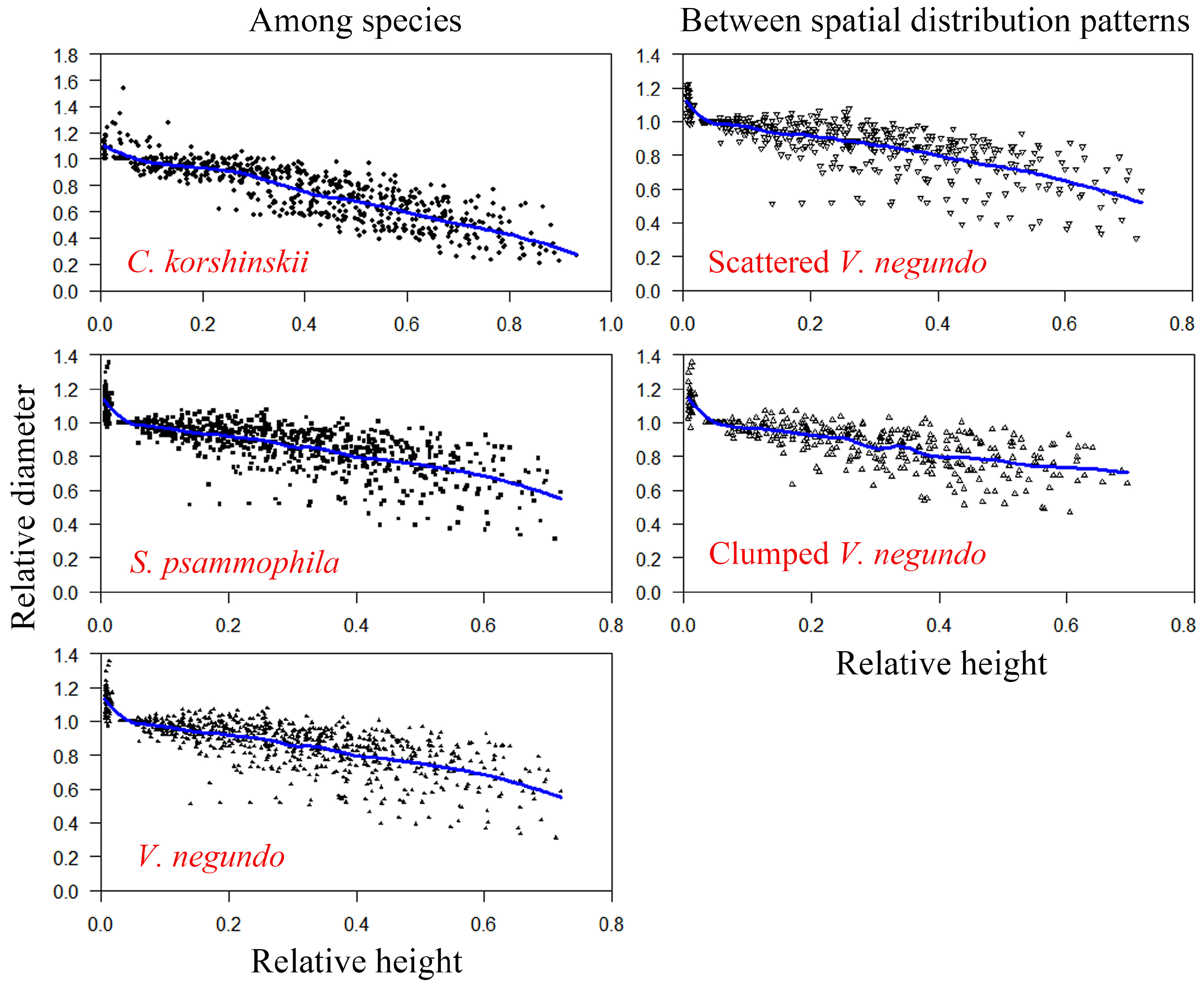
3.1. Comparison of Branch Morphology and Inflection Point
3.2. Comparison of Branch Volume Estimations and Validations
4. Discussion
4.1. Verification of Measurement Efficiency
4.2. Verification of Estimation Accuracy
4.3. Application for Shrub Volume Tables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morandini, F.; Santoni, P.A.; Tramoni, J.B.; Mell, W.E. Experimental investigation of flammability and numerical study of combustion of shrub of rockrose under severe drought conditions. Fire Safety J. 2019, 108, 102836. [Google Scholar] [CrossRef]
- Keeley, J.E.; Zedler, P.H. Large, high-intensity fire events in southern California shrublands: Debunking the fine-grain age patch model. Ecol. Appl. 2009, 19, 69–94. [Google Scholar] [CrossRef]
- Salvador, R.; Valeriano, J.; Pons, X.; Diaz-Delgado, R. A semi-automatic methodology to detect fire scars in shrubs and evergreen forests with Landsat MSS time series. Int. J. Remote Sens. 2000, 21, 655–671. [Google Scholar] [CrossRef]
- Chester, L. The 2019–2020 Australian bushfires: A potent mix of climate change, problematisation, indigenous disregard, a fractured federation, volunteerism, social media, and more. Rev. Evol. Political Econ. 2020, 1, 245–264. [Google Scholar] [CrossRef]
- Stephens, S.L.; Agee, J.K.; Fulé, P.Z.; North, M.P.; Romme, W.H.; Swetnam, T.W.; Turner, M.G. Managing forests and fire in changing climates. Science 2013, 342, 41–42. [Google Scholar] [CrossRef]
- Yin, J.; He, B.; Fan, C.; Chen, R.; Zhang, H.; Zhang, Y. Drought-related wildfire accounts for one-third of the forest wildfires in subtropical China. Agric. For. Meteorol. 2024, 346, 109893. [Google Scholar] [CrossRef]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin? Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Essaghi, S.; Hachmi, M.; Yessef, M.; Dehhaoui, M.; Sesbou, A. Litter and biomass traits of some dominant moroccan understorey fuels in five fire-prone forest regions. Bois For. Des Trop. 2019, 342, 3–16. [Google Scholar] [CrossRef]
- Harrison, G.R.; Shrestha, A.; Strand, E.K.; Karl, J.W. A comparison and development of methods for estimating shrub volume using drone-imagery-derived point clouds. Ecosphere 2024, 15, e4877. [Google Scholar] [CrossRef]
- Radeloff, V.C.; Hammer, R.B.; Stewart, S.I.; Fried, J.S.; Holcomb, S.S.; McKeefry, J.F. The Wildland–Urban Interface in the United States. Ecol. Appl. 2005, 15, 799–805. [Google Scholar] [CrossRef]
- Conti, G.; Gorné, L.D.; Zeballos, S.R.; Lipoma, M.L.; Gatica, G.; Kowaljow, E.; Whitworth-Hulse, J.I.; Cuchietti, A.; Poca, M.; Pestoni, S.; et al. Developing allometric models to predict the individual aboveground biomass of shrubs worldwide. Glob. Ecol. Biogeogr. 2019, 28, 961–975. [Google Scholar] [CrossRef]
- Yao, X.; Yang, G.; Wu, B.; Jiang, L.; Wang, F. Biomass estimation models for six shrub species in Hunshandake sandy land in inner Mongolia, Northern China. Forests 2021, 12, 167. [Google Scholar] [CrossRef]
- Ellsworth, L.M.; Newingham, B.A.; Shaff, S.E.; Williams, C.L.; Strand, E.K.; Reeves, M.; Pyke, D.A.; Schupp, E.W.; Chambers, J.C. Fuel reduction treatments reduce modeled fire intensity in the sagebrush steppe. Ecosphere 2022, 13, e4064. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Wang, Y.; Zheng, Z.; Zheng, S.; Zhao, D.; Bai, Y. UAV-based individual shrub aboveground biomass estimation calibrated against terrestrial LiDAR in a shrub-encroached grassland. Int. J. Appl. Earth Obs. Geoinf. 2021, 101, 102358. [Google Scholar] [CrossRef]
- Yépez-Rincón, F.D.; Luna-Mendoza, L.; Ramírez-Serrato, N.L.; Hinojosa-Corona, A.; Ferriño-Fierro, A.L. Assessing vertical structure of an endemic forest in succession using terrestrial laser scanning (TLS). Case study: Guadalupe Island. Remote Sens. Environ. 2021, 263, 112563. [Google Scholar] [CrossRef]
- Heinzel, J.; Huber, M.O. Detecting tree stems from volumetric TLS data in forest environments with rich understory. Remote Sens. 2017, 9, 9. [Google Scholar] [CrossRef]
- Vega, J.A.; Arellano-Pérez, S.; Álvarez-González, J.G.; Fernández, C.; Jiménez, E.; Fernández-Alonso, J.M.; Vega-Nieva, D.J.; Briones-Herrera, C.; Alonso-Rego, C.; Fontúrbel, T.; et al. Modelling aboveground biomass and fuel load components at stand level in shrub communities in NW Spain. For. Ecol. Manag. 2022, 505, 119926. [Google Scholar] [CrossRef]
- Verón, S.R.; Paruelo, J.M.; Oesterheld, M. Assessing desertification. J. Arid Environ. 2006, 66, 751–763. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Martin, A.J. Testing volume equation accuracy with water displacement techniques. For. Sci. 1984, 30, 41–50. [Google Scholar]
- Forslund, R.R. A geometrical tree volume model based on the location of the centre of gravity of the bole. Can. J. For. Res. 1982, 12, 215–221. [Google Scholar] [CrossRef]
- Wood, G.B.; Wiant, H.V.; Loy, R.J.; Miles, J.A. Centroid sampling: A variant of importance sampling for estimating the volume of sample trees of radiata pine. For. Ecol. Manag. 1990, 36, 233–243. [Google Scholar] [CrossRef]
- Özçelik, R.; Diamantopoulou, M.J.; Brooks, J.R.; Wiant, H.V. Estimating tree bole volume using artificial neural network models for four secies in Turkey. J. Environ. Manag. 2010, 91, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Behre, E.C. Preliminary notes on studies of tree form. J. For. 1923, 21, 507–511. [Google Scholar]
- Jacobs, M.; Rais, A.; Pretzsch, H. Analysis of stand density effects on the stem form of Norway spruce trees and volume miscalculation by traditional form factor equations using terrestrial laser scanning (TLS). Can. J. For. Res. 2020, 50, 51–64. [Google Scholar] [CrossRef]
- Osunkoya, O.O.; Sheng, T.K.; Mahmud, N.-A.; Damit, N. Variation in wood density, wood water content, stem growth and mortality among twenty-seven tree species in a tropical rainforest on Borneo island. Austral Ecol. 2007, 32, 191–201. [Google Scholar] [CrossRef]
- Kozak, A.; Smith, J.H.G. Standards for evaluating taper estimating systems. For. Chron. 1993, 69, 438–444. [Google Scholar] [CrossRef]
- Kozak, A. My last words on taper equations. For. Chron. 2004, 80, 507–515. [Google Scholar] [CrossRef]
- Bi, H. Trigonometric variable-form taper equations for Australian eucalypts. For. Sci. 2000, 46, 397–409. [Google Scholar] [CrossRef]
- Bailey, R.L. Upper stem volumes from stem analysis data: An overlapping bolts method. Can. J. For. Res. 1995, 25, 170–173. [Google Scholar] [CrossRef]
- Filho, A.F.; Machado, S.A.; Carneiro, M.R.A. Testing accuracy of log volume calculation procedures against water displacement techniques (Xylometer). Can. J. For. Res. 2000, 30, 990–997. [Google Scholar] [CrossRef]
- Phillips, D.R.; Taras, M.A. Accuracy of log volume estimates by density and formulas compared with water displacement. For. Prod. J. 1987, 37, 37–42. [Google Scholar]
- Özçelik, R.; Wiant, H.V.; Brooks, J.R. Accuracy using xylometry of log volume estimates for two tree species in turkey. Scand. J. For. Res. 2008, 23, 272–277. [Google Scholar] [CrossRef]
- Nicholls, D.L.; Halbrook, J.M.; Benedum, M.E.; Han, H.-S.; Lowell, E.C.; Becker, D.R.; Barbour, R.J. Socioeconomic constraints to biomass removal from forest lands for fire risk reduction in the western U.S. Forests 2018, 9, 264. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, G.G.; Liu, G.; Xu, S. Forest biomass of China: An estimate based on the biomass-volume relationship. Ecol. Appl. 1998, 8, 1084–1091. [Google Scholar]
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef]
- Yuan, C.; Guo, L.; Levia, D.F.; Rietkerk, M.; Fu, B.; Gao, G. Quantity or efficiency: Strategies of self-organized xerophytic shrubs to harvest rain. Water Resour. Res. 2022, 58, e2022WR032008. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, G.; Fu, B.; He, D.; Duan, X.; Wei, X. Temporally dependent effects of rainfall characteristics on inter- and intra-event branch-scale stemflow variability in two xerophytic shrubs. Hydrol. Earth Syst. Sci. 2019, 23, 4077–4095. [Google Scholar] [CrossRef]
- Jia, X.X.; Shao, M.A.; Wei, X.R.; Horton, R.; Li, X.Z. Estimating total net primary productivity of managed grasslands by a state-space modeling approach in a small catchment on the Loess Plateau, China. Geoderma 2011, 160, 281–291. [Google Scholar] [CrossRef]
- He, P.; Hussain, A.; Khurram Shahzad, M.; Jiang, L.; Li, F. Evaluation of four regression techniques for stem taper modeling of Dahurian Larch (Larix gmelinii) in northeastern China. For. Ecol. Manag. 2021, 494, 119336. [Google Scholar] [CrossRef]
- Husch, B.; Miller, C.I.; Beers, T.W. Forest Mensuration, 3rd ed.; Wiley: New York, NY, USA, 1982; pp. 1–402. [Google Scholar]
- Wiant, H.V.; Wood, G.B.; Furnival, G.M. Estimating log volume using the centroid position. For. Sci. 1992, 38, 187–191. [Google Scholar] [CrossRef]
- Fang, Z.; Borders, B.E.; Bailey, R.L. Compatible volume-taper models for loblolly and slash Pine based on a system with segmented-stem form factors. For. Sci. 2000, 46, 1–12. [Google Scholar] [CrossRef]
- Inoue, A. Can the relative stem taper curve be estimated theoretically only from tree height and diameter at breast height? J. For. Plan. 2001, 7, 89–94. [Google Scholar] [CrossRef]
- Bojórquez, A.; Martínez-Yrízar, A.; Búrquez, A.; Jaramillo, V.J.; Mora, F.; Balvanera, P.; Álvarez-Yépiz, J.C. Improving the accuracy of aboveground biomass estimations in secondary tropical dry forests. For. Ecol. Manag. 2020, 474, 118384. [Google Scholar] [CrossRef]
- Cleveland, W.S.; Devlin, S.J.; Grosse, E. Regression by local fitting: Methods, properties, and computational algorithms. J. Econom. 1988, 37, 87–114. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Jiang, L.C.; Brooks, J.R.; Wang, J.X. Compatible taper and volume equations for yellow-poplar in West Virginia. For. Ecol. Manag. 2005, 213, 399–409. [Google Scholar] [CrossRef]
- Özçelik, R.; Crecente-Campo, F. Stem taper equations for estimating merchantable volume of Lebanon cedar trees in the Taurus mountains, southern Turkey. For. Sci. 2016, 62, 78–91. [Google Scholar] [CrossRef]
- Clark, A.; Souter, R.A.; Schlaegel, B.E. Stem Profile for Southern Equations for Southern Tree Species; Research Paper SE-282; US Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1991; pp. 1–117.
- Cui, X.; Guo, L.; Chen, J.; Chen, X.; Zhu, X. Estimating tree-root biomass in different depths using ground-penetrating radar: Evidence from a controlled experiment. IEEE Trans. Geosci. Remote Sens. 2013, 51, 3410–3423. [Google Scholar] [CrossRef]
- Colgan, M.S.; Swemmer, T.; Asner, G.P. Structural Relationships between Form Factor, Wood Density, and Biomass in African Savanna Woodlands. Trees 2014, 28, 91–102. [Google Scholar] [CrossRef]
- Picard, N.; Rutishauser, E.; Ploton, P.; Ngomanda, A.; Henry, M. Should tree biomass allometry be restricted to power models? For. Ecol. Manag. 2015, 353, 156–163. [Google Scholar] [CrossRef]
- Dutca, I.; McRoberts, R.; Næsset, E.; Viorel, B. A practical measure for determining if diameter (D) and height (H) should be combined into D2H in allometric biomass models. Forestry 2019, 92, 627–634. [Google Scholar] [CrossRef]
- Diéguez-Aranda, U.; Castedo-Dorado, F.; Álvarez-González, J.F.; Rojo-Alboreca, A. Compatible taper function for Scots pine plantations in northwestern Spain. Can. J. For. Res. 2006, 36, 1190–1205. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Diéguez-Aranda, U.; Alvarez-Gonzalez, J.G. A growth model for Pinus radiata D. Don stands in northwestern Spain. Ann. For. Sci. 2007, 64, 453–465. [Google Scholar] [CrossRef]
- Pérez, D.H.; de Los Santos-Posadas, H.M.; Ángeles-Pérez, G.; Valdez-Lazalde, J.R.; Haller, V. Taper and Commercial volume system for Pinus patula Schltdl. et Cham. in Zacualtipán, Hidalgo State. Rev. Mex. De Cienc. For. 2013, 4, 34–45. [Google Scholar]
- Alkan, O.; Özçelik, R. Stem taper equations for diameter and volume predictions of Abies cilicica Carr. in the Taurus Mountains. Turkey J. Mt. Sci. 2020, 17, 3054–3069. [Google Scholar] [CrossRef]
- Shahzad, M.K.; Hussain, A.; Jiang, L.C. A model form for stem taper and volume estimates of Asian white birch (Betula platyphylla): A major commercial tree species of Northeast China. Can. J. For. Res. 2020, 50, 274–286. [Google Scholar] [CrossRef]
- Poudel, K.P.; Özçelik, R.; Yavuz, H. Differences in stem taper of black alder (Alnus glutinosa subsp. barbata) by origin. Can. J. For. Res. 2020, 50, 581–588. [Google Scholar] [CrossRef]
- Hussain, A.; Shahzad, M.K.; Burkhart, H.; Jiang, L.C. Stem taper functions for white birch (Betula platyphylla) and costata birch (Betula costata) in the Xiaoxing’an Mountains, northeast China. Forestry 2021, 94, 714–733. [Google Scholar] [CrossRef]
- Sánchez-Banda, M.B.; De los Santos-Posadas, H.M.; Valdez-Lazalde, J.R.; Roldán-Cortés, M.A. Heartwood functions and compatible taper and merchantable volume system for Acacia mangium Willd., in Tabasco, Mexico. Agrociencia 2022, 56, 88–107. [Google Scholar]
- Guo, F.; Su, Z.; Wang, G.; Sun, L.; Tigabu, M.; Yang, X.; Hu, H. Understanding fire drivers and relative impacts in different Chinese forest ecosystems. Sci. Total Environ. 2017, 605–606, 411–425. [Google Scholar] [CrossRef]
- Huff, S.; Ritchie, M.; Temesgen, H. Allometric equations for estimating aboveground biomass for common shrubs in northeastern California. For. Ecol. Manag. 2017, 398, 48–63. [Google Scholar] [CrossRef]
- Iqbal, S.; Zha, T.S.; Jia, X.; Hayat, M.; Qian, D.; Bourque, C.P.A.; Tian, Y.; Bai, Y.J.; Liu, P.; Yang, R.Z.; et al. Interannual variation in sap flow resprnse in three xeric shrub species to periodic drought. Agric. For. Meteorol. 2021, 297, 108276. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Rego, F.C. A new method to estimate fuel surface area-to-volume ratio using water immersion. Int. J. Wildland Fire 1998, 8, 121–128. [Google Scholar] [CrossRef]
- Hierro, J.L.; Branch, L.C.; Villarreal, D.; Clark, K.L. Predictive equations for biomass and fuel characteristics of argentine shrubs. J. Range Manag. 2000, 53, 617–621. [Google Scholar] [CrossRef]
- Li, J.; Mahalingam, S.; Weise, D.R. Experimental investigation of fire propagation in single live shrubs. Int. J. Wildland Fire 2017, 26, 58–70. [Google Scholar] [CrossRef]
- Larson, P.R. Stem form development of forest trees. For. Sci. 1963, 9, a0001–a0042. [Google Scholar] [CrossRef]
- Kumar, I.; Gupta, R.; Kumar, A. Construction of two-way volume table of Eucalyptus Tereticornis for mid-Himalayan region of Himachal Pradesh. Int. J. Bio-Resour. Stress Manag. 2020, 11, 046–050. [Google Scholar] [CrossRef]
- Norris, J.; Arnold, S.; Fairman, T. An indicative estimate of carbon stocks on Victoria’s publicly managed land using the FullCAM carbon accounting model. Aust. For. 2010, 73, 209–219. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and
Spatial Distribution Patterns | BD Categories (mm) | BD (mm) | BL (cm) | PBL (%) | |
|---|---|---|---|---|---|
| Different shrub species | C. korshinskii | <10 | 5.6 ± 3.0 | 118.0 ± 65.4 | 50.1 ± 11.6 |
| 10–20 | 14.5 ± 2.6 | 246.9 ± 30.2 | 28.9 ± 7.3 | ||
| >20 | 23.3 ± 1.5 | 301.5 ± 24.9 | 33.6 ± 6.1 | ||
| Average | 14.5 ± 0.7 | 222.1 ± 22.0 | 37.5 ± 2.9 | ||
| S. psammophila | <10 | 6.3 ± 2.2 | 168.2 ± 49.1 | 29.0 ± 8.0 | |
| 10–20 | 15.2 ± 3.3 | 284.4 ± 50.7 | 47.1 ± 10.0 | ||
| >20 | 26.0 ± 3.8 | 387.2 ± 38.3 | 58.8 ± 8.5 | ||
| Average | 15.8 ± 0.8 | 279.9 ± 6.8 | 45.0 ± 1.0 | ||
| V. negundo | <10 | 7.4 ± 1.2 | 156.5 ± 31.9 | 41.6 ± 7.6 | |
| 10–20 | 15.3 ± 2.9 | 234.1 ± 47.3 | 22.9 ± 8.3 | ||
| >20 | 22.2 ± 2.0 | 284.9 ± 47.1 | 24.6 ± 8.5 | ||
| Average | 15.0 ± 0.8 | 225.2 ± 8.8 | 29.7 ± 0.5 | ||
| Different spatial distribution patterns | Scattered V. negundo | <10 | 7.2 ± 1.2 | 158.9 ± 29.3 | 43.4 ± 8.1 |
| 10–20 | 15.6 ± 2.4 | 258.4 ± 41.1 | 18.5 ± 4.7 | ||
| >20 | 22.3 ± 2.2 | 295.3 ± 50.1 | 28.0 ± 6.5 | ||
| Average | 15.2 ± 0.6 | 237.5 ± 10.4 | 30.0 ± 1.7 | ||
| Clumped V. negundo | <10 | 7.2 ± 1.3 | 154.0 ± 35.9 | 39.7 ± 7.0 | |
| 10–20 | 15.0 ± 3.3 | 214.4 ± 43.4 | 26.5 ± 8.9 | ||
| >20 | 21.9 ± 1.4 | 253.9 ± 16.1 | 14.7 ± 5.8 | ||
| Average | 14.7 ± 1.1 | 207.5 ± 14.1 | 27.0 ± 1.6 | ||
| Species and
Spatial Distribution Patterns | Methods | Branch Volume (cm3) | RD (%) | RMSE | H | p | ||
|---|---|---|---|---|---|---|---|---|
| Median | Interquartile Range | |||||||
| Different shrub species | C. korshinskii | The reference | 104.1 ab | 8.8–214.7 | NA | NA | 8.9 | 0.031 |
| The improved | 111.5 ab | 10.4−247.0 | 15.8% | 72.6 | ||||
| Conventional cylinder | 183.4 a | 14.8−480.9 | 96.8% | 290.1 | ||||
| Conventional cone | 61.1 b | 4.9−160.3 | −34.4% | 57.3 | ||||
| S. psammophila | The reference | 120.8 ab | 35.9−438.8 | NA | NA | 10.9 | 0.012 | |
| The improved | 142.4 ab | 41.1−684.4 | 33.4% | 358.3 | ||||
| Conventional cylinder | 248.4 a | 73.0−933.0 | 115.9% | 628.4 | ||||
| Conventional cone | 82.8 b | 24.3−311.0 | −28.0% | 101.4 | ||||
| V. negundo | The reference | 197.9 b | 82.9−381.1 | NA | NA | 14.9 | 0.002 | |
| The improved | 182.1 bc | 67.6−335.7 | −8.5% | 65.8 | ||||
| Conventional cylinder | 387.8 a | 125.6−718.7 | 78.0% | 304.3 | ||||
| Conventional cone | 129.3 c | 41.9−239.6 | −40.7% | 122.5 | ||||
| Different spatial distribution patterns | Scattered V. negundo | The reference | 280.7 abc | 109.5−465.8 | NA | NA | 19.1 | <0.001 |
| The improved | 196.6 bc | 95.2−384.4 | −9.3% | 80.4 | ||||
| Conventional cylinder | 461.6 a | 171.1−859.5 | 77.9% | 353.5 | ||||
| Conventional cone | 153.9 c | 57.0−286.5 | −40.7% | 149.5 | ||||
| Clumped V. negundo | The reference | 149.5 ab | 80.6−305.4 | NA | NA | 17.2 | 0.001 | |
| The improved | 167.6 ab | 65.5−295.4 | −7.6% | 45.4 | ||||
| Conventional cylinder | 326.0 a | 129.9−585.1 | 78.1% | 241.4 | ||||
| Conventional cone | 108.7 b | 43.3−195.0 | −40.6% | 85.0 | ||||
| Species and
Spatial Distribution Patterns | Shrub ID | CH (m) | CA (m2) | BN (Unitless) | BD (mm) | BL (cm) | AV (cm3) | CV (cm3) | CV/AV (Unitless) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Different shrub species | C. korshinskii | CK1 | 2.2 | 4.1 | 34 | 8.6 | 141.8 | 2.0 × 103 | 3.0 × 106 | 1500 |
| CK2 | 2.3 | 4.4 | 24 | 8.8 | 128.0 | 1.6 × 103 | 3.3 × 106 | 2063 | ||
| CK3 | 1.9 | 3.9 | 35 | 6.6 | 118.3 | 1.2 × 103 | 2.4 × 106 | 2000 | ||
| CK4 | 2.4 | 3.7 | 37 | 8.1 | 129.2 | 2.1 × 103 | 2.9 × 106 | 1381 | ||
| Average | 2.2 | 4.0 | 33 | 8.0 | 129.3 | 1.7 × 103 a | 2.9 × 106 b | 1676 | ||
| S. psammophila | SP1 | 3.5 | 14.1 | 85 | 13.8 | 262.2 | 2.6 × 103 | 1.6 × 106 | 615 | |
| SP2 | 3.6 | 21.4 | 43 | 15.4 | 263.1 | 1.5 × 103 | 2.6 × 106 | 1733 | ||
| SP3 | 3.7 | 23.9 | 54 | 15.1 | 268.0 | 2.8 × 103 | 2.9 × 106 | 1036 | ||
| SP4 | 3.3 | 26.1 | 44 | 13.1 | 262.4 | 1.2 × 103 | 2.9 × 106 | 2417 | ||
| Average | 3.5 | 21.4 | 57 | 14.4 | 263.9 | 2.0 × 103 a | 2.5 × 106 b | 1250 | ||
| Different spatial distribution patterns | Scattered V. negundo | S-VN1 | 2.7 | 7.6 | 5 | 18.8 | 291.8 | 2.0 × 103 | 6.9 × 106 | 3450 |
| S-VN2 | 3.4 | 3.2 | 10 | 15.3 | 244.6 | 3.4 × 103 | 3.6 × 106 | 1059 | ||
| S-VN3 | 1.9 | 4.5 | 6 | 14.4 | 189.0 | 1.2 × 103 | 2.9 × 106 | 2417 | ||
| S-VN4 | 2.4 | 4.9 | 8 | 17.2 | 268.7 | 2.9 × 103 | 3.9 × 106 | 1345 | ||
| Average | 2.6 | 5.1 | 7 | 16.4 | 248.5 | 2.4 × 103 a | 4.3 × 106 b | 1819 | ||
| Clumped V. negundo | C-VN1 | 2.1 | 4.2 | 19 | 11.0 | 180.4 | 2.1 × 103 | 2.9 × 106 | 1381 | |
| C-VN2 | 2.5 | 2.4 | 16 | 11.8 | 212.7 | 2.7 × 103 | 2.0 × 106 | 741 | ||
| C-VN3 | 2.2 | 4.3 | 17 | 9.9 | 169.3 | 1.6 × 103 | 3.1 × 106 | 1938 | ||
| C-VN4 | 2.4 | 2.8 | 8 | 11.3 | 162.8 | 1.1 × 103 | 2.2 × 106 | 2000 | ||
| Average | 2.3 | 3.4 | 15 | 11.0 | 181.3 | 1.9 × 103 a | 2.6 × 106 b | 1356 | ||
| References | Country | Species | Samples | DBH (cm) | H (m) | PBL (%) | Actual Volume (m3) | Improved Method | Cylinder Method | Cone Method | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Volume (m3) | RD (%) | Volume (m3) | RD (%) | Volume (m3) | RD (%) | ||||||||
| Diéguez-Arand et al., 2006 [56] | Spain | Pinus sylvestris | 228 | 22.8 | 10.8 | 60.7 | 0.250 | 0.33 | 32.0 | 0.44 | 76.0 | 0.15 | −40.0 |
| Castedo-Dorado et al., 2007 [57] | Spain | Pinus radiata | 421 | 28.2 | 20.4 | 65.7 | 0.759 | 0.98 | 29.4 | 1.27 | 67.9 | 0.42 | −44.0 |
| Pérez et al., 2013 [58] | Mexico | Pinus patula | 78 | 27.8 | 10.3 | 82.3 | 0.517 | 0.55 | 6.4 | 0.62 | 19.9 | 0.21 | −59.4 |
| Özçelik and Crecente-Campo 2016 [50] | Turkey | Cedrus libani | 362 | 31.5 | 18.3 | 51.1 | 0.750 | 0.96 | 28.2 | 1.43 | 90.2 | 0.48 | −36.6 |
| Alkan and Özçelik 2020 [59] | Turkey | Abies cilicica | 244 | 38.6 | 17.9 | 9.1 | 1.066 | 0.83 | −22.4 | 2.10 | 96.9 | 0.70 | −34.4 |
| Shahzad et al., 2020 [60] | China | Betula platyphylla | 218 | 17.9 | 16.6 | 54.0 | 0.252 | 0.29 | 15.1 | 0.42 | 66.7 | 0.14 | −44.4 |
| Poudel et al., 2020 [61] | Turkey | Alnus glutinosa | 499 | 25.5 | 17.3 | 82.0 | 0.593 | 0.78 | 31.0 | 0.88 | 48.9 | 0.29 | −50.4 |
| 582 | 22.3 | 13.8 | 78.0 | 0.367 | 0.46 | 25.2 | 0.54 | 46.7 | 0.18 | −51.1 | |||
| 199 | 21.4 | 16.4 | 83.0 | 0.366 | 0.52 | 42.8 | 0.59 | 61.0 | 0.20 | −46.3 | |||
| Hussain et al., 2021 [62] | China | Betula costata | 108 | 31.1 | 19.3 | 54.4 | 0.900 | 1.02 | 13.4 | 1.47 | 62.9 | 0.49 | −45.7 |
| Sánchez-Banda et al., 2022 [63] | Mexico | Acacia mangium | 60 | 20.1 | 19.5 | 80.5 | 0.390 | 0.54 | 38.0 | 0.62 | 58.7 | 0.21 | −47.1 |
| 60 | 18.4 | 19.5 | 0.350 | 0.45 | 29.1 | 0.52 | 48.4 | 0.17 | −50.5 | ||||
| 31 | 20.4 | 20.3 | 0.410 | 0.58 | 40.4 | 0.66 | 61.4 | 0.22 | −46.2 | ||||
| 31 | 18.8 | 20.3 | 0.370 | 0.49 | 31.4 | 0.56 | 51.1 | 0.19 | −49.6 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, C.; Zhou, J.; Xiang, W.; Lu, N.; Hu, Y.; Guo, L.; Wang, Y.; Chen, W.; Gao, G.; Tang, Q.; et al. Improved Branch Volume Prediction of Multi-Stemmed Shrubs: Implications in Shrub Volume Inventory and Fuel Characterization. Forests 2024, 15, 1437. https://doi.org/10.3390/f15081437
Yuan C, Zhou J, Xiang W, Lu N, Hu Y, Guo L, Wang Y, Chen W, Gao G, Tang Q, et al. Improved Branch Volume Prediction of Multi-Stemmed Shrubs: Implications in Shrub Volume Inventory and Fuel Characterization. Forests. 2024; 15(8):1437. https://doi.org/10.3390/f15081437
Chicago/Turabian StyleYuan, Chuan, Jiayu Zhou, Wenhua Xiang, Nan Lu, Yanting Hu, Li Guo, Yi Wang, Weiliang Chen, Guangyao Gao, Qiang Tang, and et al. 2024. "Improved Branch Volume Prediction of Multi-Stemmed Shrubs: Implications in Shrub Volume Inventory and Fuel Characterization" Forests 15, no. 8: 1437. https://doi.org/10.3390/f15081437
APA StyleYuan, C., Zhou, J., Xiang, W., Lu, N., Hu, Y., Guo, L., Wang, Y., Chen, W., Gao, G., Tang, Q., Wang, S., Cheng, X., Gao, J., & Wei, X. (2024). Improved Branch Volume Prediction of Multi-Stemmed Shrubs: Implications in Shrub Volume Inventory and Fuel Characterization. Forests, 15(8), 1437. https://doi.org/10.3390/f15081437