
Soil-Moisture-Dependent Temperature Sensitivity of Soil Respiration in a Poplar Plantation in Northern China

Abstract
:1. Introduction
2. Materials and Methods
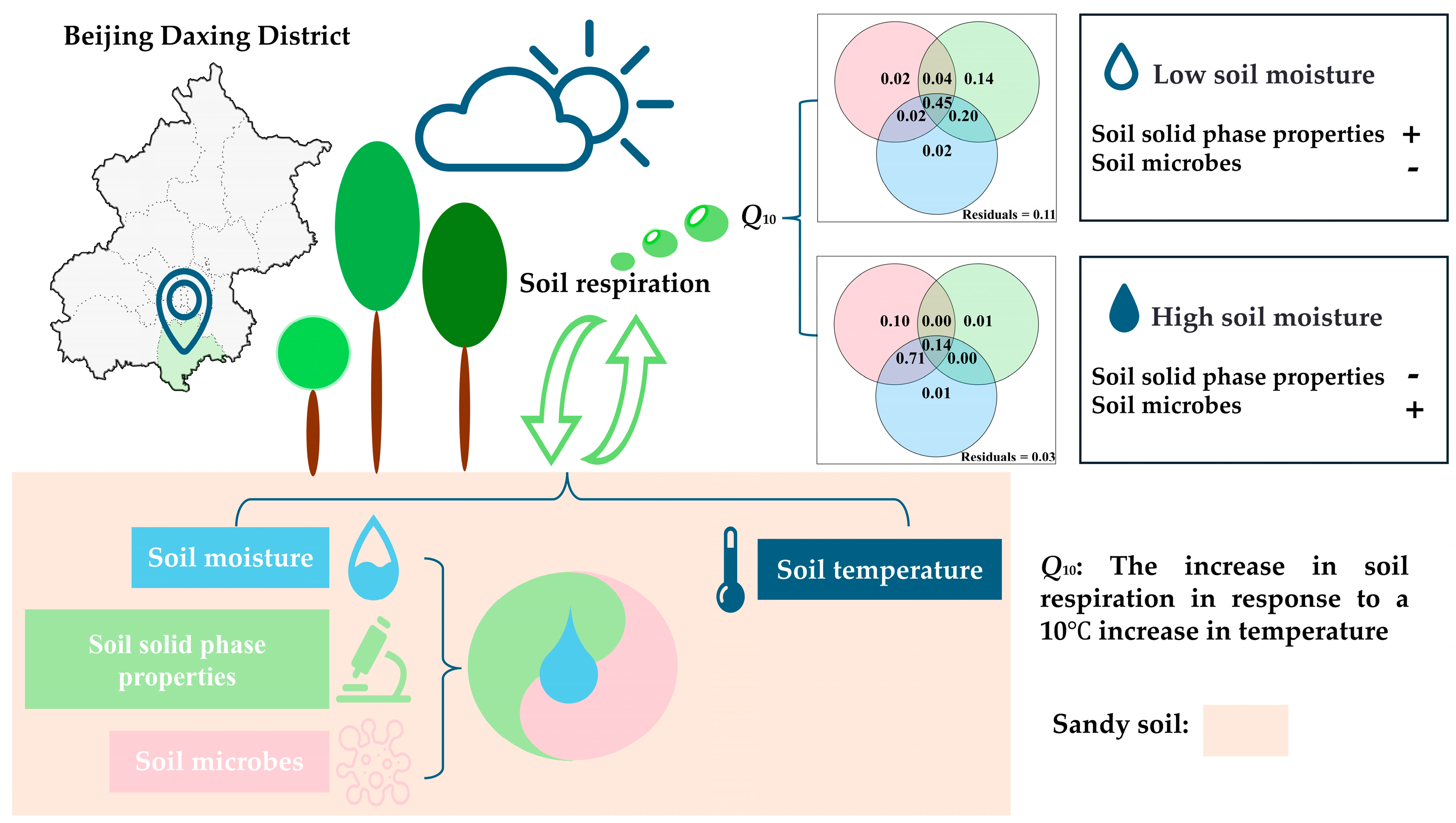
2.1. Study Area
2.2. Sampling and Pretreatment
2.3. Analysis Methods
2.3.1. Soil Physical and Chemical Properties
2.3.2. Soil Microbial Biomass Carbon and Microbial Community Composition
2.3.3. Soil Water Retention Curve
2.3.4. Soil Respiration, Soil Temperature, and Moisture
2.4. Temperature Sensitivity of Rs
2.5. Data Analysis
3. Results
3.1. Soil Properties and Soil Water Retention Curve
3.2. Soil Respiration
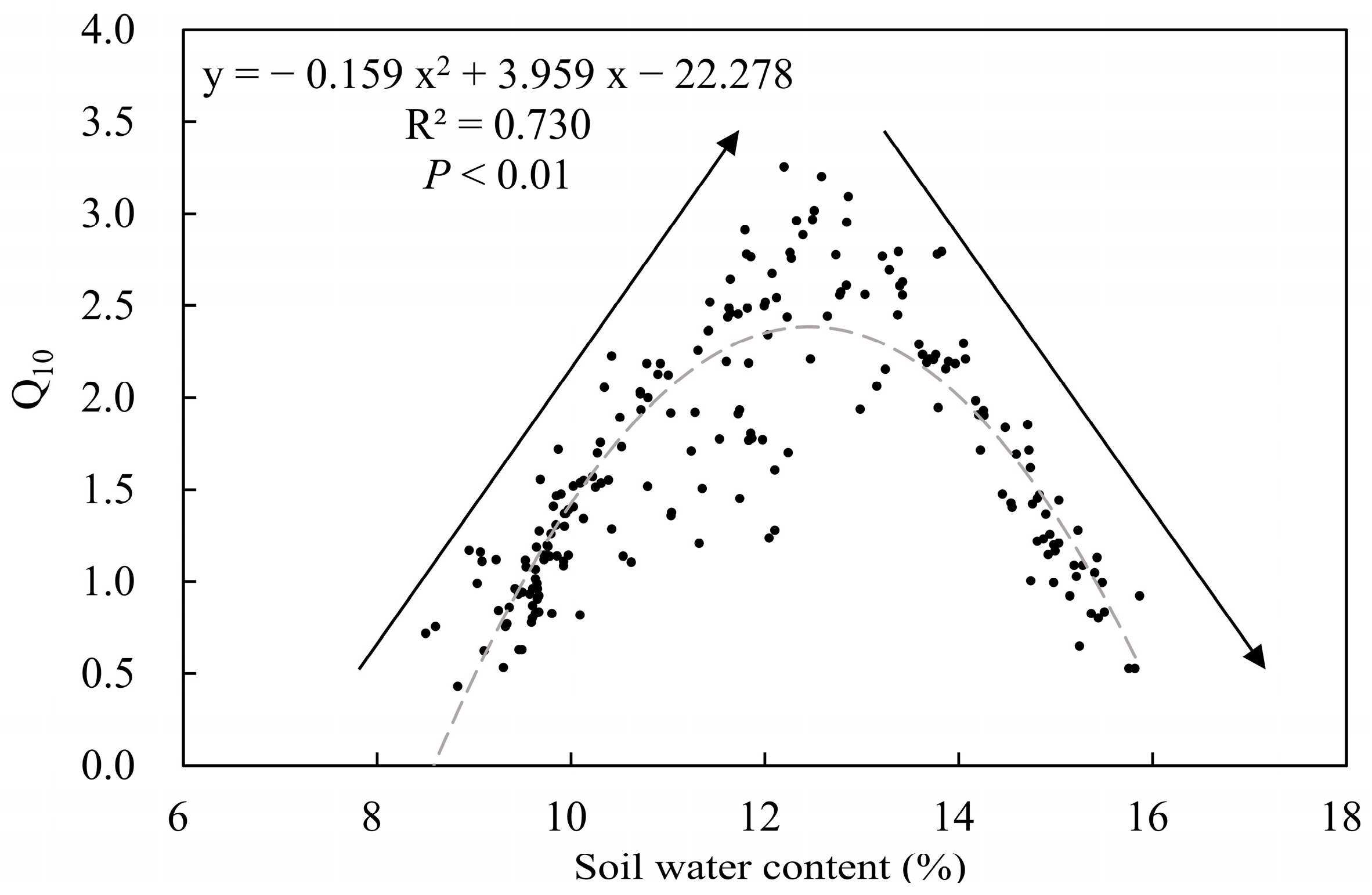
3.3. Effects of Soil Moisture on Q10
3.4. Correlations of Q10 with Soil Properties and Microbial Characteristics in Response to Soil Moisture Variability
3.5. Effects of Soil Moisture, Soil Solid-Phase Properties, and Microbial Characteristics on Q10
4. Discussion
4.1. Changes in Q10 Vary with Soil Moisture
4.2. Variability of Influencing Factors on Q10 with Soil Moisture
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.T.; Yang, J.Y.; Drury, C.F.; Yang, X.M.; Reynolds, W.D.; Li, X.G.; Hu, C.S. Evaluation of the DNDC model for simulating soil temperature, moisture and respiration from monoculture and rotational corn, soybean and winter wheat in Canada. Ecol. Model. 2017, 360, 230–243. [Google Scholar] [CrossRef]
- Fang, C.; Li, F.M.; Pei, J.Y.; Ren, J.; Gong, Y.H.; Yuan, Z.Q.; Ke, W.B.; Zheng, Y.; Bai, X.K.; Ye, J.-S. Impacts of warming and nitrogen addition on soil autotrophic and heterotrophic respiration in a semi-arid environment. Agric. For. Meteorol. 2018, 248, 449–457. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Zhou, X.H.; Liu, R.Q.; Du, Z.G.; Zhou, L.Y.; Li, S.S.; Liu, H.Y.; Shao, J.J.; Wang, J.W.; Nie, Y.Y.; et al. Soil fungi and fine root biomass mediate drought-induced reductions in soil respiration. Funct. Ecol. 2020, 34, 2634–2643. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Q.Y.; Han, Y.C.; Zhang, D.L.; Zhang, C.-C.; Hu, C.X. Carbon cycle in the microbial ecosystems of biological soil crusts. Soil Biol. Biochem. 2022, 171, 108729. [Google Scholar] [CrossRef]
- Yang, Y.; Li, T.; Pokharel, P.; Liu, L.X.; Qiao, J.B.; Wang, Y.Q.; An, S.S.; Chang, S.X. Global effects on soil respiration and its temperature sensitivity depend on nitrogen addition rate. Soil Biol. Biochem. 2022, 174, 108814. [Google Scholar] [CrossRef]
- Liu, Y.; He, N.P.; Zhu, J.X.; Xu, L.; Yu, G.R.; Niu, S.L.; Sun, X.M.; Wen, X.F. Regional variation in the temperature sensitivity of soil organic matter decomposition in China’s forests and grasslands. Glob. Change Biol. 2017, 23, 3393–3402. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.Q.; He, F.P.; Wang, W. Seasonality, rather than nutrient addition or vegetation types, influenced short-term temperature sensitivity of soil organic carbon decomposition. PLoS ONE 2016, 11, e0153415. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.Q.; Chen, L.Y.; Fang, K.; Zhang, Q.W.; Wang, J.; Liu, F.T.; Yu, J.C.; Yang, Y.H. Temperature sensitivity of SOM decomposition governed by aggregate protection and microbial communities. Sci. Adv. 2019, 5, eaau1218. [Google Scholar] [CrossRef] [PubMed]
- Haaf, D.; Six, J.; Doetterl, S. Global patterns of geo-ecological controls on the response of soil respiration to warming. Nat. Clim. Change 2021, 11, 623–627. [Google Scholar] [CrossRef]
- Wang, Q.K.; Liu, S.G.; Tian, P. Carbon quality and soil microbial property control the latitudinal pattern in temperature sensitivity of soil microbial respiration across Chinese forest ecosystems. Glob. Change Biol. 2018, 24, 2841–2849. [Google Scholar] [CrossRef] [PubMed]
- Janssens, I.A.; Pilegaard, K. Large seasonal changes in Q10 of soil respiration in a beech forest. Glob. Change Biol. 2003, 9, 911–918. [Google Scholar] [CrossRef]
- Meyer, N.; Welp, G.; Amelung, W. The temperature sensitivity (Q10) of soil respiration: Controlling factors and spatial prediction at regional scale based on environmental soil classes. Glob. Biogeochem. Cycles 2018, 32, 306–323. [Google Scholar] [CrossRef]
- Wang, C.K.; Yang, J.Y.; Zhang, Q.Z. Soil respiration in six temperate forests in China. Glob. Change Biol. 2006, 12, 2103–2114. [Google Scholar] [CrossRef]
- Yan, J.X.; Chen, L.F.; Li, J.J.; Li, H.J. Five-year soil respiration reflected soil quality evolution in different forest and grassland vegetation types in the eastern Loess Plateau of China. CLEAN Soil Air Water 2013, 41, 680–689. [Google Scholar] [CrossRef]
- Di, N.; Yang, S.J.; Liu, Y.; Fan, Y.X.; Duan, J.; Nadezhdina, N.; Li, X.M.; Xi, B.Y. Soil-moisture-dependent nocturnal water use strategy and its responses to meteorological factors in a seasonal-arid poplar plantation. Agric. Water Manag. 2022, 274, 107984. [Google Scholar] [CrossRef]
- Chen, L.H.; Mi, J.X.; Hao, L.T.; He, F.; Yang, H.B.; Wan, X.Q.; Zhang, F.; Liu, Y.; Lin, T.T. Effects of simulated nitrogen deposition on the ecophysiological responses of Populus beijingensis and P. cathayana under intra- and interspecific competition. Plant Soil 2022, 481, 127–146. [Google Scholar] [CrossRef]
- Rogers, E.R.; Zalesny, R.S.; Lin, C.-H.; Vinhal, R.A. Intrinsic and extrinsic factors influencing Populus water use: A literature review. J. Environ. Manag. 2023, 348, 119180. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Liu, Y.; Liu, Y.J.; Lu, M.Z.; Kang, X.H.; Mansfield, S.D.; Zeng, W.; Zhang, J. Opportunities and barriers for biofuel and bioenergy production from poplar. GCB Bioenergy 2021, 13, 905–913. [Google Scholar] [CrossRef]
- McIvor, I.; Douglas, G.; Dymond, J.; Eyles, G.; Marden, M. Pastoral hill slope erosion in New Zealand and the role of poplar and willow trees in its reduction. In Soil Erosion Issues in Agriculture; Godone, D., Stanchi, S., Eds.; InTech: Rijeka, Croatia, 2011; pp. 257–278. [Google Scholar] [CrossRef]
- Navarro, A.; Portillo-Estrada, M.; Ceulemans, R. Identifying the best plant water status indicator for bio-energy poplar genotypes. GCB Bioenergy 2020, 12, 426–444. [Google Scholar] [CrossRef]
- Wu, X.Y.; Xu, H.; Zha, T.G.; Zhao, Y.; Zhang, Z.Q. Soil water availability induces divergent ecosystem water-use strategies to dry-heat conditions in two poplar plantations in North China. Agric. For. Meteorol. 2024, 353, 110074. [Google Scholar] [CrossRef]
- Kooch, Y.; Rostayee, F.; Hosseini, S.M. Effects of tree species on topsoil properties and nitrogen cycling in natural forest and tree plantations of northern Iran. Catena 2016, 144, 65–73. [Google Scholar] [CrossRef]
- Man, Z.H.; Xie, C.K.; Jiang, R.Y.; Liang, A.Z.; Wu, H.; Che, S.Q. Effects of revetments on soil ecosystems in the urban river-riparian interface. iScience 2022, 25, 105277. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 30–34. [Google Scholar]
- Gee, G.W.; Or, D. Methods of soil analysis, part 4: Physical methods. In Particle-Size Analysis; Dane, J.H., Topp, G.C., Eds.; SSSA Book Ser. 5; SSSA: Madison, WI, USA, 2002; pp. 255–293. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass carbon. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Hobbs, P.J.; Frostegård, A. Changes in soil fungal: Bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol. Fert. Soils 1996, 22, 261–264. [Google Scholar] [CrossRef]
- Rinnan, R.; Bååth, E. Differential utilization of carbon substrates by bacteria and fungi in tundra soil. Appl. Environ. Microbiol. 2009, 75, 3611–3620. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef]
- Dong, H.Y.; Hu, Y.; Qian, L.W.; Yan, J.F.; Gao, L.Y.; Mei, W.X.; Zhang, J.L.; Chen, X.Z.; Wu, P.F.; Sun, Y.; et al. Preliminary manifestation of the Yangtze River Protection Strategy in improving the carbon sink function of estuary wetlands. iScience 2024, 27, 108974. [Google Scholar] [CrossRef]
- Li, J.Q.; Pei, J.M.; Pendall, E.; Fang, C.M.; Nie, M. Spatial heterogeneity of temperature sensitivity of soil respiration: A global analysis of field observations. Soil Biol. Biochem. 2020, 141, 107675. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix. 2021, R Package Version 0.92. Available online: https://github.com/taiyun/corrplot (accessed on 20 April 2023).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. In Community Ecology Package Version 2; R Software Inc.: Indianapolis, IN, USA, 2013; Volume 2, pp. 1–295. [Google Scholar]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Lilienfein, J.; Wilcke, W.; Vilela, L.; Do Carmo Lima, S.; Thomas, R.; Zech, W. Effect of no-tillage and conventional tillage systems on the chemical composition of soil solid phase and soil solution of Brazilian savanna oxisols. J. Plant Nutr. Soil Sci. 2000, 163, 411–419. [Google Scholar] [CrossRef]
- Janssen, R.P.T.; Peijnenburg, W.J.G.M.; Posthuma, L.; Van Den Hoop, M.A.G.T. Equilibrium partitioning of heavy metals in Dutch field soils. I. Relationship between metal partition coefficients and soil characteristics. Environ. Toxicol. Chem. 1997, 16, 2470–2478. [Google Scholar] [CrossRef]
- Zaidel’man, F.R.; Ustinov, M.T.; Pakhomova, E.Y. Solods of the Baraba Lowland and the Priobskoe Plateau: Their properties and genesis and the methods of their diagnostics. Eurasian Soil Sci. 2010, 43, 1069–1082. [Google Scholar] [CrossRef]
- da Costa, A.; Albuquerque, J.A.; Almeida, J.A.D.; da Costa, A.; Luciano, R.V. Pedotransfer functions to estimate retention and availability of water in soils of the state of Santa Catarina, Brazil. Rev. Bras. De Ciênc. Do Solo 2013, 37, 889–910. [Google Scholar] [CrossRef]
- Zhu, M.X.; De Boeck, H.J.; Xu, H.; Chen, Z.S.N.; Lv, J.; Zhang, Z.Q. Seasonal variations in the response of soil respiration to rainfall events in a riparian poplar plantation. Sci. Total Environ. 2020, 747, 141222. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, X.J.; Wang, S.Z.; Wang, G.D.; Zhang, J.T. Patterns and controls of temperature sensitivity of soil respiration in a meadow steppe of the Songnen Plain, Northeast China. PLoS ONE 2018, 13, e0204053. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wu, H.; Wang, Z.R.; Semenov, M.V.; Ye, J.; Yin, L.M.; Wang, X.; Kravchenko, I.; Semenov, V.; Kuzyakov, Y.; et al. Linkages between the temperature sensitivity of soil respiration and microbial life strategy are dependent on sampling season. Soil Biol. Biochem. 2022, 172, 108758. [Google Scholar] [CrossRef]
- Smith, A.P.; Bond-Lamberty, B.; Benscoter, B.W.; Tfaily, M.M.; Hinkle, C.R.; Liu, C.X.; Bailey, V.L. Shifts in pore connectivity from precipitation versus groundwater rewetting increases soil carbon loss after drought. Nat. Commun. 2017, 8, 1335. [Google Scholar] [CrossRef] [PubMed]
- Han, G.X.; Sun, B.Y.; Chu, X.J.; Xing, Q.H.; Song, W.M.; Xia, J.Y. Precipitation events reduce soil respiration in a coastal wetland based on four-year continuous field measurements. Agric. For. Meteorol. 2018, 256–257, 292–303. [Google Scholar] [CrossRef]
- Tan, J.R. The Dynamic of Soil Respiration and Its Dependence on Soil Temperature and Water Content in a Poplar Plantation in Daxing District of Beijing, China. Master’s Thesis, Beijing Forestry University, Beijing, China, 2010. (In Chinese). [Google Scholar]
- Yang, C.; Zhang, Y.J.; Rong, Y.P.; Bei, Y.X.; Wei, Y.Q.; Liu, N. Temporal variation of Q10 values in response to changes in soil physiochemical properties caused by fairy rings. Eur. J. Soil Biol. 2018, 86, 42–48. [Google Scholar] [CrossRef]
- Zhuang, W.L.; Liu, M.Y.; Wu, Y.L.; Ma, J.T.; Zhang, Y.J.; Su, L.; Liu, Y.C.; Zhao, C.C.; Fu, S.L. Litter inputs exert greater influence over soil respiration and its temperature sensitivity than roots in a coniferous forest in north-south transition zone. Sci. Total Environ. 2023, 886, 164009. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Berhe, A.A.; Khoi, C.M.; van Asperen, H.; Gillabel, J.; Six, J. Differential effects of wetting and drying on soil CO2 concentration and flux in near-surface vs. deep soil layers. Biogeochemistry 2020, 148, 255–269. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Liu, X.; Cui, Y.; Zhang, Y.; Zheng, X.; Zhang, W.; Fan, Y.; Zou, J. Effects of Drying and Rewetting Cycles on Carbon Dioxide Emissions and Soil Microbial Communities. Forests 2022, 13, 1916. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, G.L.; Yang, J.L.; Li, D.C.; Zhao, Y.G.; Liu, F.; Yang, R.M.; Yang, F. Organic matter controls of soil water retention in an alpine grassland and its significance for hydrological processes. J. Hydrol. 2014, 519, 3086–3093. [Google Scholar] [CrossRef]
- Moyano, F.E.; Manzoni, S.; Chenu, C. Responses of soil heterotrophic respiration to moisture availability: An exploration of processes and models. Soil Biol. Biochem. 2013, 59, 72–85. [Google Scholar] [CrossRef]
- Ni, H.W.; Liu, C.Y.; Sun, B.; Liang, Y.T. Response of global farmland soil organic carbon to nitrogen application over time depends on soil type. Geoderma 2022, 406, 115542. [Google Scholar] [CrossRef]
- Grandy, A.S.; Strickland, M.S.; Lauber, C.L.; Bradford, M.A.; Fierer, N. The influence of microbial communities, management, and soil texture on soil organic matter chemistry. Geoderma 2009, 150, 278–286. [Google Scholar] [CrossRef]
- Zheng, Z.M.; Yu, G.R.; Fu, Y.L.; Wang, Y.S.; Sun, X.M.; Wang, Y.H. Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study. Soil Biol. Biochem. 2009, 41, 1531–1540. [Google Scholar] [CrossRef]
- Xia, Q.; Rufty, T.; Shi, W. Soil microbial diversity and composition: Links to soil texture and associated properties. Soil Biol. Biochem. 2020, 149, 107953. [Google Scholar] [CrossRef]
- Bölscher, T.; Ågren, G.I.; Herrmann, A.M. Land-use alters the temperature response of microbial carbon-use efficiency in soils –a consumption-based approach. Soil Biol. Biochem. 2020, 140, 107639. [Google Scholar] [CrossRef]
- Zhou, W.P.; Hui, D.F.; Shen, W.J. Effects of soil moisture on the temperature sensitivity of soil heterotrophic respiration: A laboratory incubation study. PLoS ONE 2014, 9, e92531. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.M.; Liu, T.X.; Ma, L.Q.; Lei, H.M.; Singh, V.P. Analysis of soil respiration and influencing factors in a semiarid dunemeadow cascade ecosystem. Sci. Total Environ. 2021, 796, 148993. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.B.; Classen, A.T.; Kardol, P.; Yermakov, Z.; Miller, R.M. Multiple climate change factors interact to alter soil microbial community structure in an old-field ecosystem. Soil Sci. Soc. Am. J. 2011, 75, 2217–2226. [Google Scholar] [CrossRef]
- Zeng, X.M.; Feng, J.; Chen, J.; Delgado-Baquerizo, M.; Zhang, Q.G.; Zhou, X.Q.; Yuan, Y.S.; Feng, S.H.; Zhang, K.X.; Liu, Y.R.; et al. Microbial assemblies associated with temperature sensitivity of soil respiration along an altitudinal gradient. Sci. Total Environ. 2022, 820, 153257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Z.; Xue, K.; Xie, J.P.; Deng, Y.; Wu, L.Y.; Cheng, X.L.; Fei, S.F.; Deng, S.P.; He, Z.L.; Van Nostrand, J.; et al. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Change 2012, 2, 106–110. [Google Scholar] [CrossRef]
- Ma, X.L.; Jiang, S.J.; Zhang, Z.Q.; Wang, H.; Song, C.; He, J.-S. Long-term collar deployment leads to bias in soil respiration measurements. Methods Ecol. Evol. 2023, 14, 981–990. [Google Scholar] [CrossRef]
- Weiland-Brauer, N. Friends or foes—Microbial interactions in nature. Biology 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | pH | TN (g kg−1) | TP (g kg−1) | TK (g kg−1) | SOC (g kg−1) | Clay (%) | MBC (mg kg−1) | F:B | GP:GN | PLFAs (nmol g−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | 6.97 | 0.45 | 0.68 | 17.67 | 15.76 | 3.99 | 166.18 | 0.35 | 0.66 | 20.88 |
| SE | 0.06 | 0.01 | 0.02 | 0.85 | 0.58 | 0.14 | 5.13 | 0.01 | 0.02 | 0.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Zha, T.; Tan, J. Soil-Moisture-Dependent Temperature Sensitivity of Soil Respiration in a Poplar Plantation in Northern China. Forests 2024, 15, 1466. https://doi.org/10.3390/f15081466
He H, Zha T, Tan J. Soil-Moisture-Dependent Temperature Sensitivity of Soil Respiration in a Poplar Plantation in Northern China. Forests. 2024; 15(8):1466. https://doi.org/10.3390/f15081466
Chicago/Turabian StyleHe, Huan, Tonggang Zha, and Jiongrui Tan. 2024. "Soil-Moisture-Dependent Temperature Sensitivity of Soil Respiration in a Poplar Plantation in Northern China" Forests 15, no. 8: 1466. https://doi.org/10.3390/f15081466