
Changes in Soil Total and Microbial Biomass Nitrogen in Deforested and Eroded Areas in the Western Black Sea Region of Turkey


Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Data Collection
2.3. Determination of Soil Total N and Microbial Biomass N
2.4. Statistical Analysis
3. Results
3.1. Soil Total N and Microbial Biomass N Content
3.2. Nmic/Ntotal Percentage
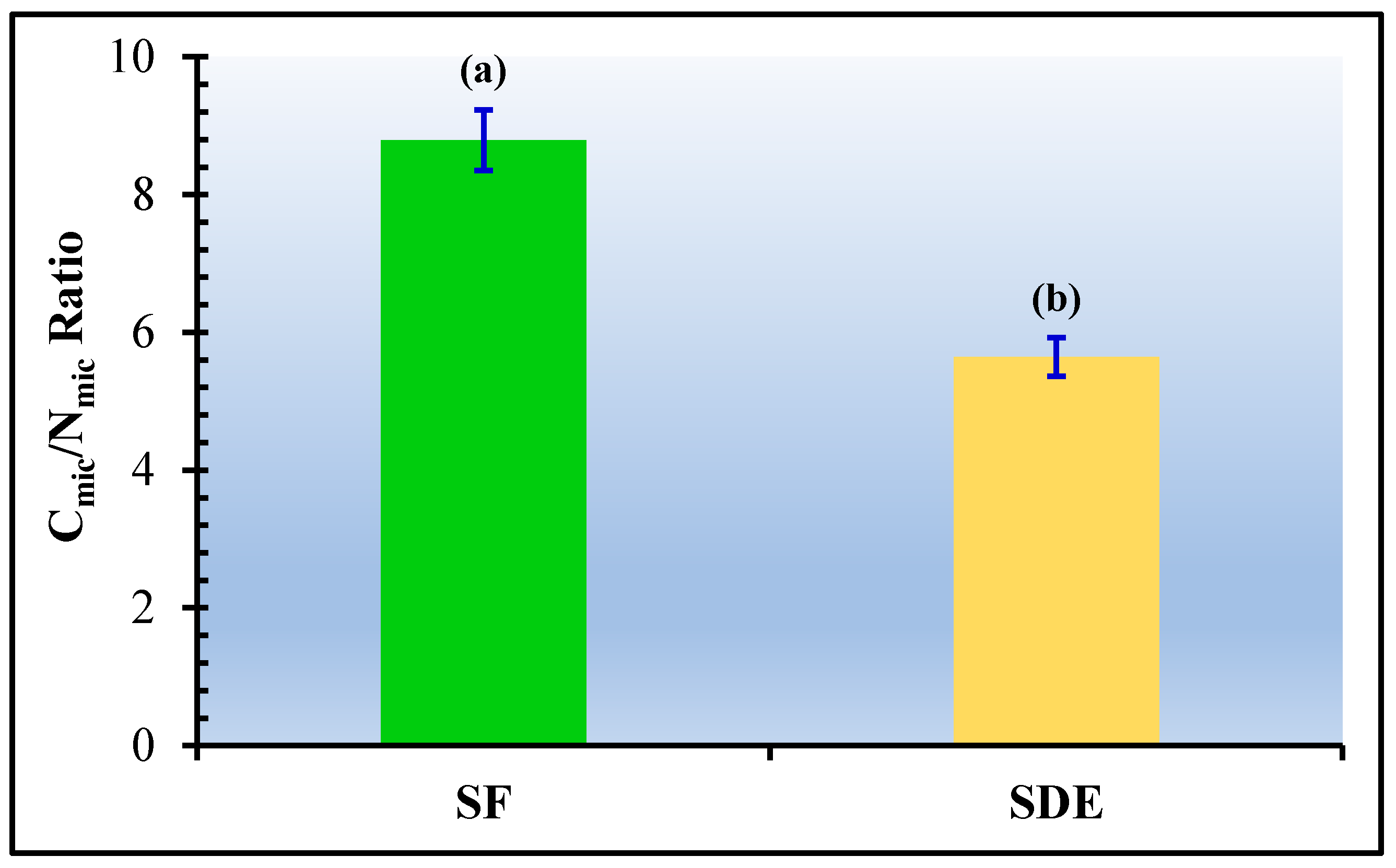
3.3. Cmic/Nmic Ratio
3.4. Relationships between Some Physical–Chemical Soil Characteristics and Ntotal and Nmic
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bargali, K.; Manral, V.; Padalia, K.; Bargali, S.S.; Upadhyay, V.P. Effect of vegetation type and season on microbial biomass carbon in Central Himalayan forest soils, India. Catena 2018, 171, 125–135. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.R.; Weil, R.R. Soil quality indicator properties in mid-Atlantic soils as influenced by conservation management. J. Water Soil Conserv. 2000, 55, 69–78. [Google Scholar]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar] [CrossRef]
- Allen, K.; Corre, M.D.; Tjoa, A.; Veldkamp, E. Soil nitrogen-cycling responses to conversion of lowland forests to oil palm and rubber plantations in Sumatra, Indonesia. PLoS ONE 2015, 10, e0133325. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Gao, W.; Yan, D. Warming suppresses microbial biomass but enhances N recycling. Soil Biol. Biochem. 2019, 131, 111–118. [Google Scholar] [CrossRef]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.Y.H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Change Biol. 2019, 25, 1078–1088. [Google Scholar] [CrossRef]
- Bolat, I.; Sensoy, H.; Özer, D. Short-term changes in microbial biomass and activity in soils under black locust trees (Robinia pseudoacacia L.) in the northwest of Turkey. J. Soils Sediments 2015, 15, 2189–2198. [Google Scholar] [CrossRef]
- Bolat, I.; Kara, Ö.; Sensoy, H.; Yüksel, K. Influences of Black Locust (Robinia pseudoacacia L.) afforestation on soil microbial biomass and activity. Iforest—Biogeosciences For. 2016, 9, 171–177. [Google Scholar] [CrossRef]
- Bolat, I.; Sensoy, H. Microbial biomass soil content and activity under black alder and sessile oak in the western Black Sea Region of Turkey. Int. J. Environ. Res. 2019, 13, 781–791. [Google Scholar] [CrossRef]
- Bhanwaria, R.; Singh, B.; Musarella, C.M. Effect of organic manure and moisture regimes on soil physiochemical properties, microbial biomass Cmic:Nmic:Pmic turnover and yield of mustard grains in arid climate. Plants 2022, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xie, W.; Ding, L.; Zhuo, Y.; Gao, Y.; Li, J.; Zhao, L. Effects of maize–crop rotation on soil physicochemical properties, enzyme activities, microbial biomass and microbial community structure in Southwest China. Microorganisms 2023, 11, 2621. [Google Scholar] [CrossRef] [PubMed]
- Junior, C.R.P.; Pereira, M.G.; Filho, J.S.O.; Beutler, S.J. Can topography affect the restoration of soil properties after deforestation in a semiarid ecosystem? J. Arid Environ. 2019, 162, 45–52. [Google Scholar] [CrossRef]
- Zeraatpisheh, M.; Bakhshandeh, E.; Hosseini, M.; Alavi, S.M. Assessing the effects of deforestation and intensive agriculture on the soil quality through digital soil mapping. Geoderma 2020, 363, 114139. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Z.; Yao, Y.; Kong, W.; Zhao, Z.; Shao, M.; Wei, X. Effects of slope morphology and position on soil nutrients after deforestation in the hilly loess region of China. Agric. Ecosyst. Environ. 2021, 321, 107615. [Google Scholar] [CrossRef]
- Rezapour, S.; Alipour, O. Degradation of Mollisols quality after deforestation and cultivation on a transect with Mediterranean condition. Environ. Earth Sci. 2017, 76, 755. [Google Scholar] [CrossRef]
- Li, M.; Zhou, X.; Zhang, Q.; Cheng, X. Consequences of afforestation for soil nitrogen dynamics in central China. Agric. Ecosyst. Environ. 2014, 183, 40–46. [Google Scholar] [CrossRef]
- Ferreira, A.C.C.; Leite, L.F.C.; De Araujo, A.S.F.; Eisenhauer, N. Land-use type effects on soil organic carbon and microbial properties in a semi-arid region of northeast Brazil. Land Degrad. Dev. 2016, 27, 171–178. [Google Scholar] [CrossRef]
- Kassa, H.; Dondeyne, S.; Poesen, J.; Frankl, A.; Nyssen, J. Impact of deforestation on soil fertility, soil carbon and nitrogen stocks: The case of the Gacheb catchment in the White Nile Basin, Ethiopia. Agric. Ecosyst. Environ. 2017, 247, 273–282. [Google Scholar] [CrossRef]
- Sirajul Haque, S.M.; Gupta, S.D.; Miah, S. Deforestation effects on biological and other important soil properties in an upland watershed of Bangladesh. J. For. Res. 2014, 25, 877–885. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Hosseini, M.; Zeraatpisheh, M.; Francaviglia, R. Land use change effects on soil quality and biological fertility: A case study in northern Iran. Eur. J. Soil Biol. 2019, 95, 103119. [Google Scholar] [CrossRef]
- De Souza, T.A.F.; Da Silva, L.J.R.; Nascimento, G.S. Amazonian deforestation and its influence on soil biotic factors and abiotic properties. Pedobiologia 2023, 97–98, 150865. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, K.; Jiang, P.; Wu, J.; Fu, W. Revealing horizontal and vertical variation of soil organic carbon, soil total nitrogen and C:N ratio in subtropical forests of southeastern China. J. Environ. Manag. 2021, 289, 112483. [Google Scholar] [CrossRef]
- Kara, O.; Sensoy, H.; Bolat, I. Slope length effects on microbial biomass and activity of eroded sediments. J. Soils Sediments 2010, 10, 434–439. [Google Scholar] [CrossRef]
- Liu, R.; Pan, Y.; Bao, H.; Liang, S.; Jiang, Y.; Tu, H.; Nong, J.; Huang, W. Variations in soil physico-chemical properties along slope position gradient in secondary vegetation of the hilly region, Guilin, southwest China. Sustainability 2020, 12, 1303. [Google Scholar] [CrossRef]
- Shi, W.; Huang, M.; Wu, L. Prediction of storm-based nutrient loss incorporating the estimated runoff and soil loss at a slope scale on the Loess Plateau. Land Degrad. Dev. 2018, 29, 2899–2910. [Google Scholar] [CrossRef]
- Feng, S.; Wen, H.; Ni, S.; Wang, J.; Cai, C. Degradation characteristics of soil-quality-related physical and chemical properties affected by collapsing gully: The case of subtropical hilly region, China. Sustainability 2019, 11, 3369. [Google Scholar] [CrossRef]
- Merlo, M.N.; Avanzi, J.C.; da Silva, L.C.M.; da Silva Aragao, O.O.; Borghi, E.; de Souza Moreira, F.M.; Thebaldi, M.S.; de Resende, A.V.; Silva, M.L.N.; Silva, B.M. Microbiological properties in cropping systems and their relationship with water erosion in the Brazilian Cerrado. Water 2022, 14, 614. [Google Scholar] [CrossRef]
- Qiu, L.; Zhu, H.; Liu, J.; Yao, Y.; Wang, X.; Rong, G.; Zhao, X.; Shao, M.; Wei, X. Soil erosion significantly reduces organic carbon and nitrogen mineralization in a simulated experiment. Agric. Ecosyst. Environ. 2021, 307, 107232. [Google Scholar] [CrossRef]
- Sensoy, H. Impact of deforestation and erosion on some soil physicochemical properties and microbial activity on steep slopes. Croat. J. For. Eng. 2024, 45, 305–318. [Google Scholar] [CrossRef]
- Karakaya, K. Numerical Simulation of the Kirazlıköprü Dam Failure on the Gökırmak River. Master’s Thesis, Middle East Technical University, Ankara, Turkey, 2005; 117p. [Google Scholar]
- Sensoy, H.; Atesoglu, A. A review of climate type variability from Bartın region. J. Bartin Fac. For. 2018, 20, 576–582. [Google Scholar]
- Sensoy, H.; Kara, Ö. Slope shape effect on runoff and soil erosion under natural rainfall conditions. iForest 2014, 7, 110–114. [Google Scholar] [CrossRef]
- Kara, Ö.; Bolat, İ.; Çakıroğlu, K.; Öztürk, M. Plant canopy effects on litter accumulation and soil microbial biomass in two temperate forests. Biol. Fertil. Soils 2008, 45, 193–198. [Google Scholar] [CrossRef]
- Devi, N.B.; Yadava, P.S. Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India. Appl. Soil Ecol. 2006, 31, 220–227. [Google Scholar] [CrossRef]
- Rowell, D.L. Soil Science: Methods and Applications, 1st ed.; Longman Group: London, UK, 1994. [Google Scholar]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods, 2nd ed.; CAB International: Wallingford, UK, 1996; pp. 68–70. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Lopes, E.L.N.; Fernandes, A.R.; Ruivo, L.P.; Cattanio, J.H.; Souza, G.F.D. Microbial biomass and soil chemical properties under different land use systems in northeastern Pará. Rev. Bras. Cienc. Solo 2011, 35, 1127–1139. [Google Scholar] [CrossRef]
- Bolat, I.; Sensoy, H. Changes in soil microbial biomass and activity under different land-use types in Bartın, northwestern Türkiye. Contemp. Probl. Ecol. 2023, 16, 886–889. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, W.; Ye, Y.; Zhao, J.; Wang, K. Soil aggregate mediates the impacts of land uses on organic carbon, total nitrogen, and microbial activity in a Karst ecosystem. Sci. Rep. 2017, 7, 41402. [Google Scholar] [CrossRef]
- Razzaghi, S.; Islam, K.R.; Ahmed, I.A.M. Deforestation impacts soil organic carbon and nitrogen pools and carbon lability under Mediterranean climates. J. Soils Sediments 2022, 22, 2381–2391. [Google Scholar] [CrossRef]
- Kara, O.; Bolat, I. The effect of different land uses on soil microbial biomass carbon and nitrogen in Bartın province. Turk. J. Agric. For. 2008, 32, 281–288. [Google Scholar]
- Tiwari, S.; Singh, C.; Boudh, S.; Rai, P.K.; Gupta, V.K.; Singh, J.S. Land use change: A key ecological disturbance declines soil microbial biomass in dry tropical uplands. J. Environ. Manag. 2019, 242, 1–10. [Google Scholar] [CrossRef]
- Gao, F.; Cui, X.; Sang, Y.; Song, J. Changes in soil organic carbon and total nitrogen as affected by primary forest conversion. For. Ecol. Manag. 2020, 463, 118013. [Google Scholar] [CrossRef]
- Groppo, J.D.; Lins, S.R.M.; Camargo, P.B.; Assad, E.D.; Pinto, H.S.; Martins, S.C.; Salgado, P.R.; Evangelista, B.; Vasconcellos, E.; Sano, E.E.; et al. Changes in soil carbon, nitrogen, and phosphorus due to land-use changes in Brazil. Biogeosciences 2015, 12, 4765–4780. [Google Scholar] [CrossRef]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activities. Catena 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Haghverdi, K.; Kooch, Y. Soil carbon and nitrogen fractions in response to land use/cover changes. Acta Oecologica 2020, 109, 103659. [Google Scholar] [CrossRef]
- Zhu, B.; Li, Z.; Li, P.; Liu, G.; Xue, S. Soil erodibility, microbial biomass, and physical–chemical property changes during long-term natural vegetation restoration: A case study in the Loess Plateau, China. Ecol. Res. 2010, 25, 531–541. [Google Scholar] [CrossRef]
- Berhe, A.A.; Torn, M.S. Erosional redistribution of topsoil controls soil nitrogen dynamics. Biogeochemistry 2017, 132, 37–54. [Google Scholar] [CrossRef]
- Restrepo, J.D.; Kettner, A.J.; Syvitski, J.P.M. Recent deforestation causes rapid increase in river sediment load in the Colombian Andes. Anthropocene 2015, 10, 13–28. [Google Scholar] [CrossRef]
- Parhizkar, M.; Shabanpour, M.; Zema, D.A.; Lucas-Borja, M.E. Rill erosion and soil quality in forest and deforested ecosystems with different morphological characteristics. Resources 2020, 9, 129. [Google Scholar] [CrossRef]
- Chuma, G.B.; Mondo, J.M.; Ndeko, A.B.; Mugumaarhahama, Y.; Bagula, E.M.; Blaise, M.; Valerie, M.; Jacques, K.; Karume, K.; Mushagalusa, G.N. Forest cover affects gully expansion at the tropical watershed scale: Case study of Luzinzi in Eastern DR Congo. Trees For. People 2021, 4, 100083. [Google Scholar] [CrossRef]
- Bauhus, J.D.; Pare, D.; Cote, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Khan, K.S.; Joergensen, R.G. Microbial C, N, and P relationships in moisture-stressed soils of Potohar, Pakistan. J. Plant Nutr. Soil Sci. 2006, 169, 494–500. [Google Scholar] [CrossRef]
- Wen, L.; Lei, P.; Xiang, W.; Yan, W.; Liu, S. Soil microbial biomass carbon and nitrogen in pure and mixed stands of Pinus massoniana and Cinnamomum camphora differing in stand age. For. Ecol. Manag. 2014, 328, 150–158. [Google Scholar] [CrossRef]
- Foote, J.A.; Boutton, T.W.; Scott, D.A. Soil C and N storage and microbial biomass in US southern pine forests: Influence of forest management. For. Ecol. Manag. 2015, 355, 48–57. [Google Scholar] [CrossRef]
- Singh, K.P.; Mandal, T.N.; Tripathi, S.K. Patterns of restoration of soil physciochemical properties and microbial biomass in different landslide sites in the sal forest ecosystem of Nepal Himalaya. Ecol. Eng. 2001, 17, 385–401. [Google Scholar] [CrossRef]
- Nsabimana, D.; Haynes, R.J.; Wallis, F.M. Size, activity and catabolic diversity of the soil microbial biomass as affected by land use. Appl. Soil Ecol. 2004, 26, 81–92. [Google Scholar] [CrossRef]
- Evangelou, E.; Tsadilas, C.; Giourga, C. Seasonal variation of soil microbial biomass carbon and nitrogen as affected by land use in a mediterranean agro ecosystem. Commun. Soil Sci. Plant Anal. 2021, 52, 222–234. [Google Scholar] [CrossRef]
- Moore, J.M.; Klose, S.; Tabatabai, M.A. Soil microbial biomass carbon and nitrogen as affected by cropping systems. Biol. Fertil. Soils 2000, 31, 200–210. [Google Scholar] [CrossRef]
- Kara, O.; Bolat, I. Soil microbial biomass C and N changes in relation to forest conversion in the Northwestern Turkey. Land Degrad. Dev. 2008, 19, 421–428. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Ladd, J.N. Microbial biomass in soil measurement and turnover. In Soil Biochemistry; Paul, E.A., Ladd, J.N., Eds.; Marcel Dekker Inc.: New York, NY, USA; Basel, Switzerland, 1981; Volume 5, pp. 415–471. [Google Scholar]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Kallenbach, C.; Grandy, A.S. Controls over soil microbial biomass responses to carbon amendments in agricultural systems: A meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 241–252. [Google Scholar] [CrossRef]
- Smith, L.C.; Orgiazzi, A.; Eisenhauer, N.; Cesarz, S.; Lochner, A.; Jones, A.; Bastida, F.; Patoine, G.; Reitz, T.; Buscot, F.; et al. Large-scale drivers of relationships between soil microbial properties and organic carbon across Europe. Glob. Ecol. Biogeogr. 2021, 30, 2070–2083. [Google Scholar] [CrossRef]
- Li, X.; Qu, Z.; Zhang, Y.; Ge, Y.; Sun, H. Soil fungal community and potential function in different forest ecosystems. Diversity 2022, 14, 520. [Google Scholar] [CrossRef]
- Bauhus, J.; Khanna, P.K. The significance of microbial biomass in forest soils. In Going Underground—Ecological Studies in Forest Soils; Rastin, N., Bauhus, J., Eds.; Research Signpost: Trivandrum, India, 1999; pp. 77–110. [Google Scholar]
- Högberg, M.N.; Högberg, P.; Myrold, D.D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 2007, 150, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Pietikäinen, J.; Pettersson, M.; Bååth, E. Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol. Ecol. 2005, 52, 49–58. [Google Scholar] [CrossRef]
- Dinesh, R.; Chaudhuri, S.G.; Ganeshamurthy, A.N.; Dey, C. Changes in soil microbial indices and their relationships following deforestation and cultivation in wet tropical forests. Appl. Soil Ecol. 2003, 24, 17–26. [Google Scholar] [CrossRef]
- Kooch, Y.; Tavakoli, M.; Akbarinia, M. Microbial/biochemical indicators showing perceptible deterioration in the topsoil due to deforestation. Ecol. Indic. 2018, 91, 84–91. [Google Scholar] [CrossRef]
- Nunes, J.S.; Araujo, A.S.F.; Nunes, L.A.P.L.; Lima, L.M.; Carneiro, R.F.V.; Salviano, A.A.C.; Tsai, S.M. Impact of land degradation on soil microbial biomass and activity in northeast Brazil. Pedosphere 2012, 22, 88–95. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Characteristics | SF | SDE |
|---|---|---|
| Sand (%) | 33.6 a | 46.9 b |
| Silt (%) | 18.1 a | 14.5 a |
| Clay (%) | 48.3 a | 38.6 a |
| Soil texture | Clay | Sandy clay |
| Bulk density (g cm−3) | 1.16 a | 1.53 b |
| Pore space (%) | 51.83 a | 41.69 b |
| Soil temperature in sites (°C) | 18.77 a | 21.90 b |
| Soil moisture content (%) | 21.74 a | 13.10 b |
| >2 mm/<2 mm (%) | 22.48 a | 40.16 b |
| pH (H2O) | 5.31 a | 7.72 b |
| Electrical conductivity (μS cm−1) | 39.45 a | 61.80 b |
| Soil organic C (g kg−1) | 22.53 a | 4.41 b |
| Soil Characteristics | Person Correlation and Significance Level | Nmic | Ntotal | Cmic/Nmic | OC | ST | SM | pH | EC | BD | C | Nmic/Ntotal |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nmic | Pearson correlation | 1 | 0.675 ** | 0.562 * | 0.715 ** | −0.341 | 0.688 ** | −0.796 ** | −0.350 | −0.699 ** | 0.281 | 0.641 * |
| Sig. (2-tailed) | 0.006 | 0.029 | 0.003 | 0.214 | 0.005 | 0.000 | 0.201 | 0.004 | 0.311 | 0.010 | ||
| Ntotal | Pearson correlation | 1 | 0.611 * | 0.913 ** | −0.397 | 0.821 ** | −0.677 ** | −0.403 | −0.800 ** | 0.224 | −0.053 | |
| Sig. (2-tailed) | 0.015 | 0.000 | 0.143 | 0.000 | 0.006 | 0.137 | 0.000 | 0.422 | 0.851 | |||
| Cmic/Nmic | Pearson correlation | 1 | 0.702 ** | −0.061 | 0.576 * | −0.749 ** | −0.477 | −0.673 ** | 0.498 | 0.197 | ||
| Sig. (2-tailed) | 0.004 | 0.829 | 0.025 | 0.001 | 0.072 | 0.006 | 0.059 | 0.481 | ||||
| OC | Pearson correlation | 1 | −0.228 | 0.775 ** | −0.733 ** | −0.365 | −0.857 ** | 0.150 | 0.034 | |||
| Sig. (2-tailed) | 0.414 | 0.001 | 0.002 | 0.180 | 0.000 | 0.594 | 0.905 | |||||
| ST | Pearson correlation | 1 | −0.447 | 0.488 | 0.580 * | 0.359 | −0.292 | −0.018 | ||||
| Sig. (2-tailed) | 0.095 | 0.065 | 0.024 | 0.189 | 0.292 | 0.950 | ||||||
| SM | Pearson correlation | 1 | −0.752 ** | −0.540 * | −0.742 ** | 0.222 | 0.062 | |||||
| Sig. (2-tailed) | 0.001 | 0.038 | 0.002 | 0.427 | 0.828 | |||||||
| pH | Pearson correlation | 1 | 0.726 ** | 0.855 ** | -0.559 * | −0.326 | ||||||
| Sig. (2-tailed) | 0.002 | 0.000 | 0.030 | 0.236 | ||||||||
| EC | Pearson correlation | 1 | 0.525 * | -0.751 ** | −0.018 | |||||||
| Sig. (2-tailed) | 0.044 | 0.001 | 0.950 | |||||||||
| BD | Pearson correlation | 1 | -0.327 | −0.014 | ||||||||
| Sig. (2-tailed) | 0.235 | 0.960 | ||||||||||
| C | Pearson correlation | 1 | 0.249 | |||||||||
| Sig. (2-tailed) | 0.370 | |||||||||||
| Nmic/Ntotal | Pearson correlation | 1 | ||||||||||
| Sig. (2-tailed) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolat, İ.; Sensoy, H. Changes in Soil Total and Microbial Biomass Nitrogen in Deforested and Eroded Areas in the Western Black Sea Region of Turkey. Forests 2024, 15, 1468. https://doi.org/10.3390/f15081468
Bolat İ, Sensoy H. Changes in Soil Total and Microbial Biomass Nitrogen in Deforested and Eroded Areas in the Western Black Sea Region of Turkey. Forests. 2024; 15(8):1468. https://doi.org/10.3390/f15081468
Chicago/Turabian StyleBolat, İlyas, and Huseyin Sensoy. 2024. "Changes in Soil Total and Microbial Biomass Nitrogen in Deforested and Eroded Areas in the Western Black Sea Region of Turkey" Forests 15, no. 8: 1468. https://doi.org/10.3390/f15081468