


EIAGA-S: Rapid Mapping of Mangroves Using Geospatial Data without Ground Truth Samples

Abstract
1. Introduction
- (1)
- Stable and unsupervised segmentation: The EIAGA-S model eliminates the need for extensive field sampling and labeled data, achieving stable and accurate mapping results without relying on ground truth samples. This makes it a practical and efficient solution for large-scale mangrove monitoring.
- (2)
- Boundary and small object segmentation: The EIAGA-S model excels in boundary delineation and small object segmentation. It accurately identifies fine details such as small river tributaries and complex terrain features, outperforming other methods like GA, K-means, SVM, and U-Net in handling intricate and dispersed objects.
- (3)
- The development and application of MEVI: A new Mangrove Enhanced Vegetation Index (MEVI) was developed to better distinguish mangroves from other vegetation types. This index improves the discrimination within the spectral feature space, enhancing the overall segmentation accuracy and supporting effective monitoring in areas like the Hainan Dongzhai Port Mangrove Nature Reserve.
2. Materials
3. Methodology
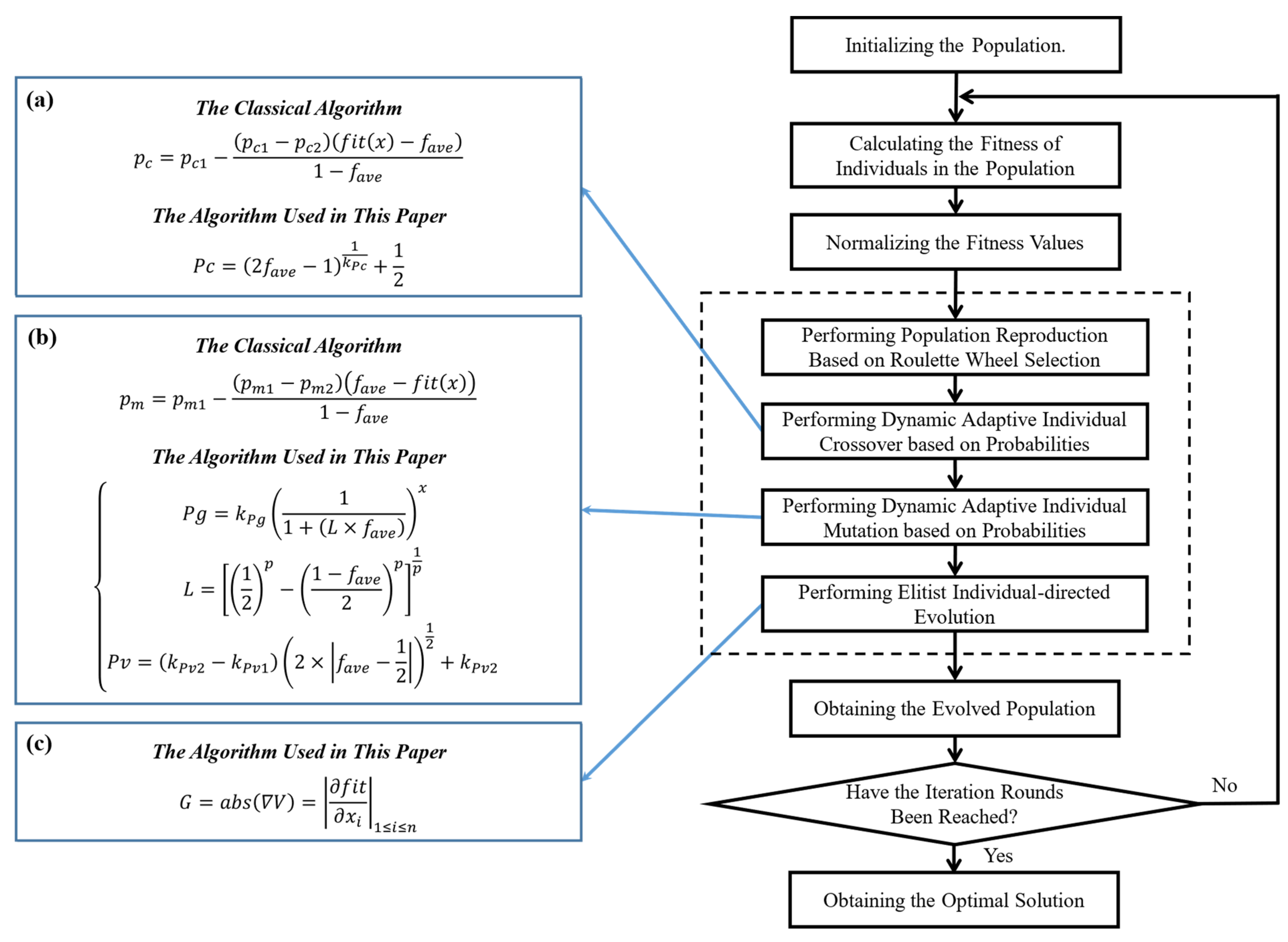
3.1. Elite Individual Adaptive Genetic Algorithm
3.1.1. Population Encoding and Fitness Function
3.1.2. Adaptive Crossover and Mutation Strategies
3.1.3. Elite-Individual-Oriented Evolutionary Strategy
3.2. Semantic Information Enhancement Method for Mangrove (MEVI)
3.3. Semantic Feature Decision
3.3.1. Connectivity Analysis
3.3.2. Classification Decision
3.4. Experimental Evaluation Indicators
4. Experimental Results and Analysis
4.1. Precision and Result Analysis of EIAGA-S
4.2. Analysis of Generalization Capability of the EIAGA-S Model
4.3. Analysis of EIAGA-S Model’s Effectiveness on Boundaries and Small Target Objects
5. Discussion
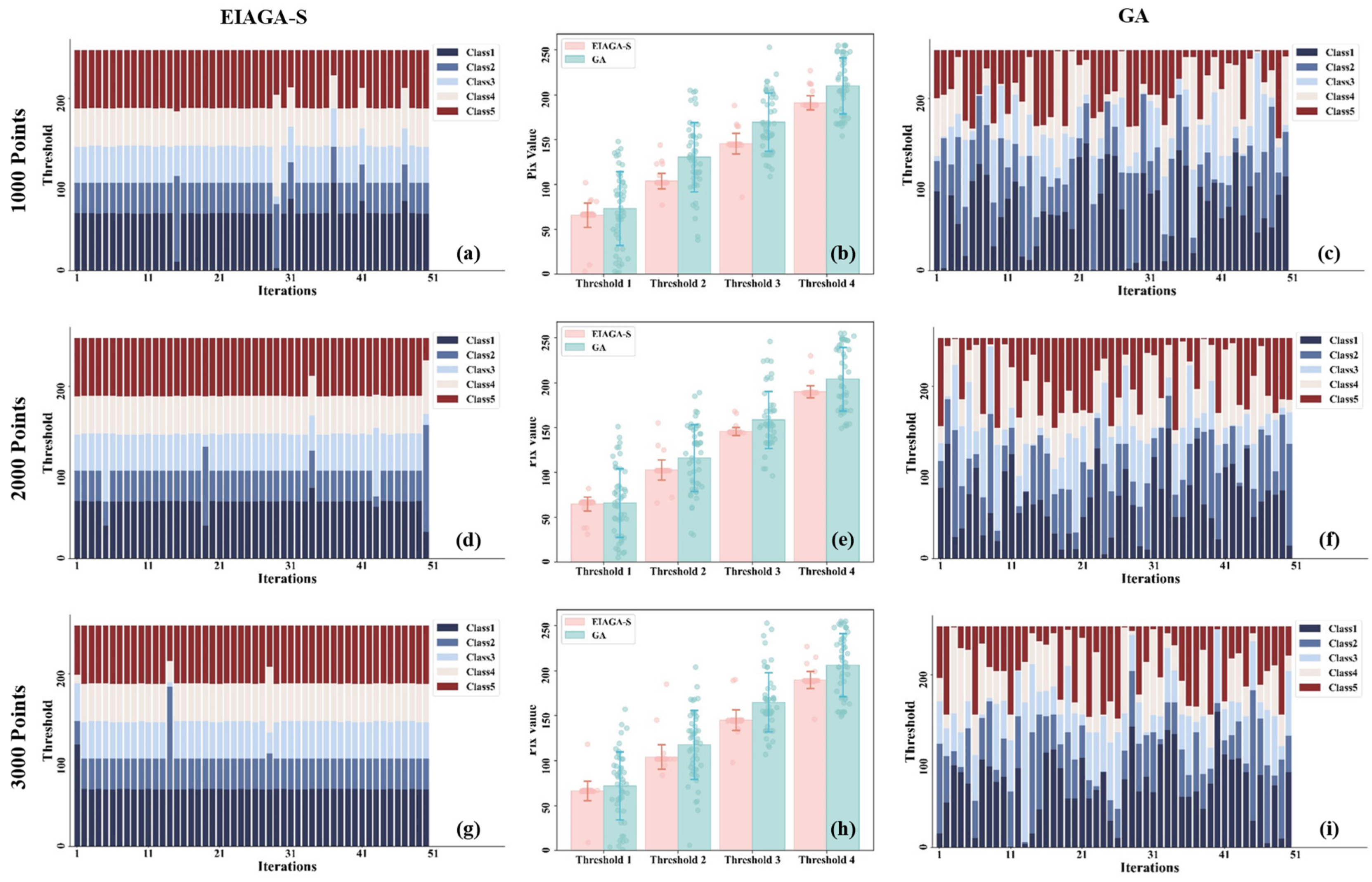
5.1. Analysis of Model Convergence and Stability for EIAGA-S
5.2. Comparison of Different Vegetation Index Results
5.3. Analysis of MEVI’s Best Empirical Parameters and Results
5.4. Advantages and Challenges of EIAGA-S
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rijal, S.S.; Pham, T.D.; Noer’Aulia, S.; Putera, M.I.; Saintilan, N. Mapping Mangrove Above-Ground Carbon Using Multi-Source Remote Sensing Data and Machine Learning Approach in Loh Buaya, Komodo National Park, Indonesia. Forests 2023, 14, 94. [Google Scholar] [CrossRef]
- Schürholz, D.; Castellanos-Galindo, G.A.; Casella, E.; Mejía-Rentería, J.C.; Chennu, A. Seeing the forest for the trees: Mapping cover and counting trees from aerial images of a mangrove forest using artificial intelligence. Remote Sens. 2023, 15, 3334. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, G.; de Souza Filho, C.R. Tracking canopy gaps in mangroves remotely using deep learning. Remote Sens. Ecol. Conserv. 2022, 8, 890–903. [Google Scholar] [CrossRef]
- Lu, C.; Li, L.; Wang, Z.; Su, Y.; Su, Y.; Huang, Y.; Jia, M.; Mao, D. The national nature reserves in China: Are they effective in conserving mangroves? Ecol. Indic. 2022, 142, 109265. [Google Scholar] [CrossRef]
- de Souza Moreno, G.M.; de Carvalho Júnior, O.A.; de Carvalho, O.L.F.; Andrade, T.C. Deep semantic segmentation of mangroves in Brazil combining spatial, temporal, and polarization data from Sentinel-1 time series. Ocean Coast. Manag. 2023, 231, 106381. [Google Scholar] [CrossRef]
- Xu, C.; Wang, J.; Sang, Y.; Li, K.; Liu, J.; Yang, G. An Effective Deep Learning Model for Monitoring Mangroves: A Case Study of the Indus Delta. Remote Sens. 2023, 15, 2220. [Google Scholar] [CrossRef]
- Wu, S.; Zeng, W.; Chen, H. A sub-pixel image registration algorithm based on SURF and M-estimator sample consensus. Pattern Recognit. Lett. 2020, 140, 261–266. [Google Scholar] [CrossRef]
- Thakur, S.; Mondal, I.; Ghosh, P.; Das, P.; De, T. A review of the application of multispectral remote sensing in the study of mangrove ecosystems with special emphasis on image processing techniques. Spat. Inf. Res. 2020, 28, 39–51. [Google Scholar] [CrossRef]
- Fu, C.; Song, X.; Xie, Y.; Wang, C.; Luo, J.; Fang, Y.; Cao, B.; Qiu, Z. Research on the spatiotemporal evolution of mangrove forests in the Hainan Island from 1991 to 2021 based on SVM and Res-UNet Algorithms. Remote Sens. 2022, 14, 5554. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Mao, D.; Ren, C.; Song, K.; Zhao, C.; Wang, C.; Xiao, X.; Wang, Y. Mapping global distribution of mangrove forests at 10-m resolution. Sci. Bull. 2023, 12, 1306–1316. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Tan, Z.; Liu, X.; Li, M. Swin-UperNet: A Semantic Segmentation Model for Mangroves and Spartina alterniflora Loisel Based on UperNet. Electronics 2023, 12, 1111. [Google Scholar] [CrossRef]
- Gao, E.; Zhou, G. Spatio-Temporal Changes of Mangrove-Covered Tidal Flats over 35 Years Using Satellite Remote Sensing Imageries: A Case Study of Beibu Gulf, China. Remote Sens. 2023, 15, 1928. [Google Scholar] [CrossRef]
- Dong, H.; Gao, Y.; Chen, R.; Wei, L. MangroveSeg: Deep-Supervision-Guided Feature Aggregation Network for Mangrove Detection and Segmentation in Satellite Images. Forests 2024, 15, 127. [Google Scholar] [CrossRef]
- Zhang, Z.; Ahmed, M.R.; Zhang, Q.; Li, Y.; Li, Y. Monitoring of 35-Year Mangrove Wetland Change Dynamics and Agents in the Sundarbans Using Temporal Consistency Checking. Remote Sens. 2023, 15, 625. [Google Scholar] [CrossRef]
- Wu, S.; Chen, H. Smart city oriented remote sensing image fusion methods based on convolution sampling and spatial transformation. Comput. Commun. 2020, 157, 444–450. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, S.; Huang, Y. Transformer-Based Model with Dynamic Attention Pyramid Head for Semantic Segmentation of VHR Remote Sensing Imagery. Entropy 2022, 24, 1619. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, Y.; Wang, Y.; Chen, J.; Zang, T.; Chen, H. Convolution Feature Inference-Based Semantic Understanding Method for Remote Sensing Images of Mangrove Forests. Electronics 2023, 12, 881. [Google Scholar] [CrossRef]
- Tang, R.; Pu, F.; Yang, R.; Xu, Z.; Xu, X. Multi-domain fusion graph network for semi-supervised PolSAR image classification. Remote Sens. 2022, 15, 160. [Google Scholar] [CrossRef]
- Li, X.; Pu, F.; Yang, R.; Gui, R.; Xu, X. AMN: Attention metric network for one-shot remote sensing image scene classification. Remote Sens. 2020, 12, 4046. [Google Scholar] [CrossRef]
- Robin, S.L.; Marchand, C.; Mathian, M.; Baudin, F.; Alfaro, A.C. Distribution and bioaccumulation of trace metals in urban semi-arid mangrove ecosystems. Front. Environ. Sci. 2022, 10, 2202. [Google Scholar] [CrossRef]
- Long, J.; Shelhamer, E.; Darrell, T. Fully convolutional networks for semantic segmentation. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Boston, MA, USA, 7–12 June 2015; pp. 3431–3440. [Google Scholar]
- Li, J.; Pu, F.; Chen, H.; Xu, X.; Yu, Y. Crop Segmentation of Unmanned Aerial Vehicle Imagery Using Edge Enhancement Network. IEEE Geosci. Remote Sens. Lett. 2024. [Google Scholar] [CrossRef]
- Tong, Q.; Wu, J.; Zhu, Z.; Zhang, M.; Xing, H. STIRUnet: SwinTransformer and inverted residual convolution embedding in unet for Sea–Land segmentation. J. Environ. Manag. 2024, 357, 120773. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Tan, X.; Guo, B.; Zhu, K.; Liao, P.; Wang, T.; Wang, Q.; Zhang, X. SDFCNv2: An improved FCN framework for remote sensing images semantic segmentation. Remote Sens. 2021, 13, 4902. [Google Scholar] [CrossRef]
- Guo, M.; Yu, Z.; Xu, Y.; Huang, Y.; Li, C. ME-Net: A Deep Convolutional Neural Network for Extracting Mangrove Using Sentinel-2A Data. Remote Sens. 2021, 13, 1292. [Google Scholar] [CrossRef]
- Fu, B.; He, X.; Yao, H.; Liang, Y.; Deng, T.; He, H.; Fan, D.; Lan, G.; He, W. Comparison of RFE-DL and stacking ensemble learning algorithms for classifying mangrove species on UAV multispectral images. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102890. [Google Scholar] [CrossRef]
- Sun, Z.; Jiang, W.; Ling, Z.; Zhong, S.; Zhang, Z.; Song, J.; Xiao, Z. Using Multisource High-Resolution Remote Sensing Data (2 m) with a Habitat–Tide–Semantic Segmentation Approach for Mangrove Mapping. Remote Sens. 2023, 15, 5271. [Google Scholar] [CrossRef]
- Wu, S.; Bai, Y.; Chen, H. Change detection methods based on low-rank sparse representation for multi-temporal remote sensing imagery. Clust. Comput. 2019, 22, 9951–9966. [Google Scholar] [CrossRef]
- Chu, B.; Gao, F.; Chai, Y.; Liu, Y.; Yao, C.; Chen, J.; Wang, S.; Li, F.; Zhang, C. Large-area full-coverage remote sensing image collection filtering algorithm for individual demands. Sustainability 2021, 13, 13475. [Google Scholar] [CrossRef]
- Zhang, R.; Jia, M.; Wang, Z.; Zhou, Y.; Mao, D.; Ren, C.; Zhao, C.; Liu, X. Tracking annual dynamics of mangrove forests in mangrove National Nature Reserves of China based on time series Sentinel-2 imagery during 2016–2020. Int. J. Appl. Earth Obs. Geoinf. 2022, 112, 102918. [Google Scholar] [CrossRef]
- Copenhaver, K.L. Combining Tabular and Satellite-Based Datasets to Better Understand Cropland Change. Land 2022, 11, 714. [Google Scholar] [CrossRef]
- Li, H.; Hu, B.; Li, Q.; Jing, L. CNN-based individual tree species classification using high-resolution satellite imagery and airborne LiDAR data. Forests 2021, 12, 1697. [Google Scholar] [CrossRef]
- Fu, L.; Chen, J.; Wang, Z.; Zang, T.; Chen, H.; Wu, S.; Zhao, Y. MSFANet: Multi-scale fusion attention network for mangrove remote sensing lmage segmentation using pattern recognition. J. Cloud Comput. 2024, 13, 27. [Google Scholar] [CrossRef]
- Hashim, F.A.; Houssein, E.H.; Hussain, K.; Mabrouk, M.S.; Al-Atabany, W. Honey Badger Algorithm: New metaheuristic algorithm for solving optimization problems. Math. Comput. Simul. 2022, 192, 84–110. [Google Scholar] [CrossRef]
- Lian, J.; Hui, G. Human evolutionary optimization algorithm. Expert Syst. Appl. 2024, 241, 122638. [Google Scholar] [CrossRef]
- Jayathunga, S.; Pearse, G.D.; Watt, M.S. Unsupervised Methodology for Large-Scale Tree Seedling Mapping in Diverse Forestry Settings Using UAV-Based RGB Imagery. Remote Sens. 2023, 15, 5276. [Google Scholar] [CrossRef]
- Li, X.; Zheng, H.; Han, C.; Wang, H.; Dong, K.; Jing, Y.; Zheng, W. Cloud detection of superview-1 remote sensing images based on genetic reinforcement learning. Remote Sens. 2020, 12, 3190. [Google Scholar] [CrossRef]
- Shen, Y.; Wei, Y.; Zhang, H.; Rui, X.; Li, B.; Wang, J. Unsupervised Change Detection in HR Remote Sensing Imagery Based on Local Histogram Similarity and Progressive Otsu. Remote Sens. 2024, 16, 1357. [Google Scholar] [CrossRef]
- Dutta, K.; Talukdar, D.; Bora, S.S. Segmentation of unhealthy leaves in cruciferous crops for early disease detection using vegetative indices and Otsu thresholding of aerial images. Measurement 2022, 189, 110478. [Google Scholar] [CrossRef]
- Ma, G.; Yue, X. An improved whale optimization algorithm based on multilevel threshold image segmentation using the Otsu method. Eng. Appl. Artif. Intell. 2022, 113, 104960. [Google Scholar] [CrossRef]
- Amirteimoori, A.; Mahdavi, I.; Solimanpur, M.; Ali, S.S.; Tirkolaee, E.B. A parallel hybrid PSO-GA algorithm for the flexible flow-shop scheduling with transportation. Comput. Ind. Eng. 2022, 173, 108672. [Google Scholar] [CrossRef]
- Tebbal, I.; Hamida, A.F. Effects of Crossover Operators on Genetic Algorithms for the Extraction of Solar Cell Parameters from Noisy Data. Eng. Technol. Appl. Sci. Res. 2023, 13, 10630–10637. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Wang, C.; Mao, D.; Zhang, Y. A new vegetation index to detect periodically submerged mangrove forest using single-tide Sentinel-2 imagery. Remote Sens. 2019, 11, 2043. [Google Scholar] [CrossRef]
- Tian, Y.; Jia, M.; Wang, Z.; Mao, D.; Du, B.; Wang, C. Monitoring invasion process of Spartina alterniflora by seasonal Sentinel-2 imagery and an object-based random forest classification. Remote Sens. 2020, 12, 1383. [Google Scholar] [CrossRef]
- Díaz, B.M.; Blackburn, G.A. Remote sensing of mangrove biophysical properties: Evidence from a laboratory simulation of the possible effects of background variation on spectral vegetation indices. Int. J. Remote Sens. 2003, 24, 53–73. [Google Scholar] [CrossRef]
- de Jong, S.M.; Shen, Y.; de Vries, J.; Bijnaar, G.; van Maanen, B.; Augustinus, P.; Verweij, P. Mapping mangrove dynamics and colonization patterns at the Suriname coast using historic satellite data and the LandTrendr algorithm. Int. J. Appl. Earth Obs. Geoinf. 2021, 97, 102293. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, L. How to automate timely large-scale mangrove mapping with remote sensing. Remote Sens. Environ. 2021, 264, 112584. [Google Scholar] [CrossRef]
- Zhang, X.; Treitz, P.M.; Chen, D.; Quan, C.; Shi, L.; Li, X. Mapping mangrove forests using multi-tidal remotely-sensed data and a decision-tree-based procedure. Int. J. Appl. Earth Obs. Geoinf. 2017, 62, 201–214. [Google Scholar] [CrossRef]
- Chandrasekar, K.; Sesha Sai, M.; Roy, P.; Dwevedi, R. Land Surface Water Index (LSWI) response to rainfall and NDVI using the MODIS Vegetation Index product. Int. J. Remote Sens. 2010, 31, 3987–4005. [Google Scholar] [CrossRef]
- Xiang, K.; Yuan, W.; Wang, L.; Deng, Y. An LSWI-based method for mapping irrigated areas in China using moderate-resolution satellite data. Remote Sens. 2020, 12, 4181. [Google Scholar] [CrossRef]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, M.; Zhang, H.; Liu, Y. Mapping mangrove using a red-edge mangrove index (REMI) based on Sentinel-2 multispectral images. IEEE Trans. Geosci. Remote Sens. 2023. [Google Scholar] [CrossRef]
- Herrmann, I.; Karnieli, A.; Bonfil, D.; Cohen, Y.; Alchanatis, V. SWIR-based spectral indices for assessing nitrogen content in potato fields. Int. J. Remote Sens. 2010, 31, 5127–5143. [Google Scholar] [CrossRef]
- Schuster, C.; Förster, M.; Kleinschmit, B. Testing the red edge channel for improving land-use classifications based on high-resolution multi-spectral satellite data. Int. J. Remote Sens. 2012, 33, 5583–5599. [Google Scholar] [CrossRef]
- Kumar, T.; Mandal, A.; Dutta, D.; Nagaraja, R.; Dadhwal, V.K. Discrimination and classification of mangrove forests using EO-1 Hyperion data: A case study of Indian Sundarbans. Geocarto Int. 2019, 34, 415–442. [Google Scholar] [CrossRef]
- van Beers, F.; Lindström, A.; Okafor, E.; Wiering, M. Deep neural networks with intersection over union loss for binary image segmentation. In Proceedings of the 8th International Conference on Pattern Recognition Applications and Methods, Prague, Czech Republic, 19–21 February 2019; pp. 438–445. [Google Scholar]
- Maung, W.S.; Tsuyuki, S.; Guo, Z. Improving land use and land cover information of Wunbaik mangrove area in Myanmar using U-Net model with multisource remote sensing datasets. Remote Sens. 2023, 16, 76. [Google Scholar] [CrossRef]
- Shulei, W.; Fengru, Z.; Huandong, C.; Yang, Z. Semantic understanding based on multi-feature kernel sparse representation and decision rules for mangrove growth. Inf. Process. Manag. 2022, 59, 102813. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algorithm | Classes | ||||
|---|---|---|---|---|---|
| Mangrove | Ocean | Bare Soil | Shrub | Residential Area | |
| GA | 0.313 | 0.385 | 0.104 | 0.262 | 0.302 |
| K-means | 0.875 | 0.403 | 0.135 | 0.448 | 0.207 |
| SVM | 0.567 | 0.580 | 0.013 | 0.318 | 0.030 |
| U-Net | 0.857 | 0.815 | 0.272 | 0.369 | 0.102 |
| EIAGA-S | 0.920 | 0.930 | 0.450 | 0.616 | 0.902 |
| Algorithm | Classes | ||||
|---|---|---|---|---|---|
| Mangrove | Ocean | Bare Soil | Shrub | Residential Area | |
| GA | 0.476 | 0.556 | 0.188 | 0.415 | 0.464 |
| K-means | 0.933 | 0.574 | 0.237 | 0.618 | 0.344 |
| SVM | 0.723 | 0.734 | 0.026 | 0.482 | 0.059 |
| U-Net | 0.923 | 0.898 | 0.428 | 0.539 | 0.185 |
| EIAGA-S | 0.958 | 0.964 | 0.621 | 0.762 | 0.948 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wu, S.; Zhang, X.; Luo, H.; Chen, H.; Song, C. EIAGA-S: Rapid Mapping of Mangroves Using Geospatial Data without Ground Truth Samples. Forests 2024, 15, 1512. https://doi.org/10.3390/f15091512
Zhao Y, Wu S, Zhang X, Luo H, Chen H, Song C. EIAGA-S: Rapid Mapping of Mangroves Using Geospatial Data without Ground Truth Samples. Forests. 2024; 15(9):1512. https://doi.org/10.3390/f15091512
Chicago/Turabian StyleZhao, Yuchen, Shulei Wu, Xianyao Zhang, Hui Luo, Huandong Chen, and Chunhui Song. 2024. "EIAGA-S: Rapid Mapping of Mangroves Using Geospatial Data without Ground Truth Samples" Forests 15, no. 9: 1512. https://doi.org/10.3390/f15091512
APA StyleZhao, Y., Wu, S., Zhang, X., Luo, H., Chen, H., & Song, C. (2024). EIAGA-S: Rapid Mapping of Mangroves Using Geospatial Data without Ground Truth Samples. Forests, 15(9), 1512. https://doi.org/10.3390/f15091512