Abstract
A predominant management practice to reduce wind erosion in the arid deserts of northwest China is the planting of shrubs. However, the carbon sequestration capacity of these sand-fixing plantations has not received much attention. In this study, the carbon sequestration capacity of six typical sand-fixing plantations (Haloxylon ammodendron (C. A. Mey.) Bunge, Caragana korshinskii Kom., Tamarix ramosissima Ledeb., Calligonum mongolicum Turcz., Artemisia desertorum Spreng. and Hedysarum scoparium Fisch. & C. A. Mey.) in the Shiyang River Basin were compared and analyzed. We evaluated how carbon sequestration may vary among different species, and examined if plantation age or management style (such as the additional construction of sand barriers, enclosure) positively or negatively influenced the carbon storage potential of these plantation ecosystems. Our results showed that all six plantations could store carbon, but plant species is the controlling factor driving carbon stock accumulation in plantations. The actual organic carbon stored beneath 25-year-old T. ramosissima, H. ammodendron, C. korshinskii, H. scoparium, C. mongolicum and A. desertorum plantations was 45.80, 31.80, 20.57, 20.2, 8.24 and1.76 Mg ha−1, respectively. Plantations using a clay–sand barrier had 1.3 times the carbon sequestration capacity of plantations that only used wheat straw and sand barriers. Similarly, enclosed plantations had 1.4 times the carbon storage capacity of unenclosed plantations. Plantation age greatly impacts carbon sequestration capacity. A 25-year-old H. ammodendron plantation has a carbon sequestration capacity three times greater than that of 3-year plantation. We conclude that while afforesting arid areas, H. ammodendron and T. ramosissima should be prioritized, and priority also should be given to using clay–sand barrier and enclosure.
1. Introduction
Global warming threatens global economic development [1,2,3]. Many countries have engaged in conservation measures to limit the global surface average temperature increase [4,5,6]. Afforestation is the planting of trees on land that was previously treeless, with the goal of increasing carbon storage and mitigating global warming [7,8,9]. It has been widely implemented in various regions, including drylands, to counteract desertification and increase carbon sequestration [7,10,11,12]. Drylands cover a significant portion of the global land area [13], and they play a crucial role in terrestrial carbon sequestration [14,15,16,17]. Additionally, afforestation in arid areas avoids land-use competition with agriculture and has great potential for carbon sequestration [16]. However, scientific afforestation planning needs to be conducted since human activities such as afforestation may cause the reduction in groundwater sources in arid and semi-arid areas [18].
To make scientific afforestation planning for increasing carbon storage in arid and semi-arid areas, three issues must be addressed. First, verifying the carbon sequestration potential of sand-fixing plantations. Numerous studies have focused on estimating the carbon sequestration potential of various afforestation strategies in arid areas [7,10,11,12,19,20,21,22,23]. Most of those studies have shown that sand-fixing plantations have a greater carbon sequestration potential than that of unplanted sand dunes [7,10,11,12]. However, the effect of land-use/cover conversion on carbon sinks is still under debate [21]. In addition to carbon sequestration, afforestation in arid areas has been evaluated for its ecological benefits and economic value [10,24,25]. Most studies have revealed positive benefits, such as better eco-services, resilience, health, and long-term productivity [7,22,26,27]. Nevertheless, some studies have highlighted negative impacts [28], particularly in regions with water scarcity, where large-scale afforestation can consume much water [29,30].
Second, determining the effect of factors on carbon sequestration in sand-fixing plantations. Previous studies have primarily focused on specific factors influencing carbon sequestration, such as plant species, plantation management, and soil depth [31]. However, the total carbon storage of plantations in dry areas is influenced by multiple factors [32], and a comprehensive understanding of these factors is needed to optimize plantation management for climate change mitigation. Planting fast-growing trees or shrubs can be beneficial for soil carbon sequestration [20], and designing and establishing complex forest systems in arid areas can enhance ecosystem services, resilience, health, and long-term productivity [33,34]. Additionally, the choice of plant species is crucial, especially in regions with extreme water scarcity [29,35]. Unfortunately, many of those studies have only assessed one shrub species [10,11,12].
Third, clarifying the distribution of carbon sequestration in various parts of plants, such as the proportion of carbon sequestration in aboveground and underground parts. Some studies have focused on the distribution of carbon in different plant compartments and in different depths of soil [7,19,20,32,33]. Those studies have found that the biomass of planted species is a powerful control on carbon sequestration and the effects of afforestation on soil carbon primarily appear in the upper 20 cm of the soil [36]. The effects of afforestation on soil carbon stock can vary depending on factors such as plant species [22], plantation management [17,32], irrigation [17,34,35], and the choice to include or exclude grazing species [28,37]. For example, afforestation on sand dunes has a high potential to sequester soil inorganic carbon (SIC) through pedogenic carbonate formation [19,38]. However, afforestation can also increase SIC in acidic soils and decrease SIC in alkaline soils, and the effects can vary among different tree species and soil depths [22]. The management practices of plantations, such as irrigation and grazing exclusion, have also gained attention, but their effects on soil carbon sequestration are still controversial.
Most of the above have investigated the carbon sequestration of afforestation in arid areas from one of the three perspectives above, which is of great benefit to our scientific afforestation. However, there is still a lack of comprehensive research on sand-fixing plantations from three perspectives: carbon sequestration capacity, influencing factors, and distribution of plant carbon sequestration.
Shiyang River Basin is located in a fragile ecological environment and a sensitive climate zone in China. It is one of the earliest desert grassland areas in China to have carried out desertification control and growing artificial sand-fixation vegetation. Its desertification and sand-fixation vegetation construction are representative of arid areas. We have investigated the impact of Haloxylon ammodendron (C. A. Mey.) Bunge on the carbon sequestration of desert ecosystems in our previous study [11]. However, under the carbon peak and carbon neutrality scenario, and against the background of reconstructing “New Three-North” [39], the characteristics and mechanism of carbon sequestration, and the scientific planning of artificial sand-fixing plantations in this region still require further research.
Our hypothesis is that the artificial sand-fixing plantations in the Shiyang River Basin have carbon sequestration capacity, and forest management measures have a significant impact on its carbon sequestration potential. Precisely, our focus was to understand the dynamics of carbon balance in sand-fixing plantations in the Shiyang River Basin of Gansu province, northwest China, under the influence of various factors, including shrub species, plantation management practices, plantation ages, and climates. The specific goals of this study are as follows:
- To estimate and analyze the carbon storage of six typical sand-fixing plantation ecosystems in comparison with zonal vegetation in the desert steppe.
- To evaluate how plantation management impacts carbon storage and sequestration capacity.
- To provide suggestions for optimizing plantation management in the study area.
By examining the carbon sequestration potential of different shrub species based on various influencing factors, this study aims to find a trade-off in the establishment of artificial plantations for climate change mitigation in arid areas. The findings of this study will contribute to the understanding of afforestation as a strategy to improve soil carbon sequestration and provide insights for optimizing plantation management practices in dry areas.
2. Materials and Methods
2.1. Study Area Description
This study was conducted in the Shiyang River Basin (36°29′~39°27′ N, 101°41′~104°16′ E, 1300–1810 m.a.s.l.) over a total area of 41,600 km2 in the east of the Hexi Corridor of northwest China (Figure 1). The study area has a temperate desert climate, having an annual precipitation of less than 150 mm and annual evaporation of 2000–2600 mm. The dryness of the area ranges from 15 to 25 on the Selianinov index [40], with more than 3000 h of annual sunshine per year. The average annual temperature is 7.8–10.0 °C.

Figure 1.
Study region and distribution of sand-fixing plantations.
The research area belongs to the temperate xeric, with four distinct seasons and a dry climate. The vegetation is mainly those growing in temperate desert, and the soil is desert soil. In the area, the native desert vegetation is composed of Nitraria tangutorum Bobrov, Caragana korshinskii Kom., Artemisia ordosica Krasch., Artemisia desertorum Spreng., Psammochloa villosa (Trin.) Bor, Stipa breviflora Griseb., Agriophyllum squarrosum (L.) Moq. Among them, N. tangutorum is predominant and considered the climax community. The artificial vegetation is mainly sand-fixing plantations, including H. ammodendron, C. korshinskii, Tamarix ramosissima Ledeb., Calligonum mongolicum Turcz., Hedysarum scoparium Fisch. & C. A. Mey. The surface runoff of the Shiyang River has declined continuously since the 1960s, with a reduction of 74.3% due to frequent over-exploitation of surface and groundwater resources. Since 2007, wells have been closed and farmland areas have been cut back as a part of the ‘Comprehensive Control of the Shiyang River Basin’ Project, the mean annual surface runoff of the lower Shiyang River was about 3 billion m3 after 2022.
2.2. Sampling Plot
Along the middle reaches of the Shiyang River Basin and its lower ones as well, artificial sand-fixing plantations of H. ammodendron, C. korshinskii, T. ramosissima, C. mongolicum, A. desertorum and H. scoparium were selected. Sample plots of sand-fixing plantations were constructed in different years. N. tangutorum shrub was selected as the control, which was the largest natural top-level vegetation community in the research area. Land afforested with H. ammodendron, C. mongolicum, A. desertorum, and H. scoparium was considered ‘mobile sand dunes’. Land afforested with C. korshinskii was considered ‘inter-dune land’. Land afforested with T. ramosissima was considered sand-covered salinized land.
In this study, we selected paired plots of 3-year and 25-year H. ammodendron and T. ramosissima shrubs to determine the influence of plantation age on ecosystem carbon storage. We chose these species as they are the most typical sand-fixing shrub planted in arid China. To study the influence of sand barriers on ecosystem carbon storage, we selected two types of sand barriers, clay–sand barrier and semi-mobile with a straw–sand barrier, respectively. Clay or wheat–straw–sand barriers are the most common plantation modes in the Shiyang River Basin. We selected 25-year H. ammodendron plantations with clay–sand and semi-mobile straw–sand barriers to compare the influence of this management style while keeping plant species and plantation age constant. Studies on management measures including grazing enclosed and unenclosed were conducted using H. scoparium. We established three sample plots and conducted vegetation and soil surveys from early August to middle September during the peak of the growing season. A fence with a height of 1.5 m was established to block and prohibit human activities such as grazing from one section of the plantation. Unenclosed areas were open to grazing species.
2.3. Vegetation Survey and Biomass Carbon Storage Measurement
Three 20 m × 20 m replicate sample plots of each artificial sand-fixing plantation and its control site were used to establish plant community composition and estimate shrub population demography through community and biomass surveys. A total of 45 sample plots were established, including 12 plots of H. ammodendron of different ages and with sand barriers, 6 plots of H. scoparium with or without enclosure, 6 plots of T. ramosissima of different ages, 9 plots of other C. korshinskii, D. mongolicum and A. desertorum sand-fixing plantations, and 12 control plots of stable natural N. tangutorum, mobile sand dune, inter-dune land and sand-covered salinized land. Within each of the plots, five small quadrats (1 m × 1 m) were set up at the corners and center of each plot to investigate herbaceous community composition and biomass. At each plantation plot, we investigated each shrub, with its crown length, width, height and ground diameter measured. And the number of plants (clusters) of each shrub was also recorded.
The above-ground biomass, below-ground biomass, and biomass of litter of each sand-fixing plantation were determined at harvest [41]. Litter included leaves and other organic detritus smaller than 5 cm, and so on. The ‘standard’ plants of H. ammodendron, C. korshinskii, T. ramosissima, C. mongolicum, A. desertorum, H. scoparium, and N. tangutorum were selected from each plot. The above-ground and below-ground biomass of each individual was measured. Biomass per unit area was calculated using the average density of the population. In each quadrat (1 m × 1 m) surveyed, all the above-ground green parts were scissored. All green biomass was homogenized as one sample, irrespective of species. Five small quadrats (1 m × 1 m) were homogenized into one group for collecting the litter in the sampling plots, and it was weighed and recorded as well. At the same time, 1 m pits were dug to extract the roots of plants in the herbaceous layer. The plant samples were taken to the laboratory. Then, they were dried in an oven at 80 °C for 72 h. Their dry weight was measured then, with the unit being g/m2.
By drying and weighing, water content (W) of plant samples was analyzed; carbon content of plant samples (C) was determined using the potassium dichromate oxidation method [42].
Carbon storage of above-ground biomass (BCS1):
BCS1 = ∑(Bi × (1 − Wi%) × Ci/Si)
i indicates different plants in the plots and small quadrat (herbaceous); Bi indicates above-ground biomass; Wi indicates water content of shoots; Ci indicates the organic carbon content of shoots; and Si indicates the plot area.
Carbon storage of below-ground biomass (BCS2):
BCS2 = ∑(Bj × (1 − Wj%) × Cj/Sj)
j indicates the different groups within the sampling plots; Bj indicates below-ground biomass at the depth of 1 m; Wj indicates water content of roots; Cj indicates the organic carbon content of roots, and Sj indicates the plot area.
Litter biomass carbon storage in plants (BCS3):
BCS3 = B × (1 − W%) × C/S
B is the fresh litter biomass; W is the water content of litter; C is the organic C content of litter; S is the plot area.
Biomass carbon (BC) storage of plantations (BCS):
BCS = BCS1 + BCS2 + BCS3
BC sink of plantations (ΔBCS):
ΔBCS = (BCS1 + BCS2 + BCS3)SP − (BCS1 + BCS2 + BCS3)CK
SP indicates the artificial sand-fixing plantations; CK indicates the control plots for afforesting sand-fixing plantations.
2.4. Soil Sampling and Organic Carbon Storage Measurement
In each plot, we dug three sampling ditches for the soil at seven depths (0–5, 5–10, 10–20, 20–40, 40–60, 60–80, and 80–100 cm), with three replicates. Currently, most studies use a depth of 1 m as the reference standard for calculating soil organic carbon [43]. Soil samples were put in sealed plastic bags and returned to the laboratory. Soils were rid of roots and debris by sieving (2 mm). Then, soil water content, bulk density, particle size, organic carbon, active organic carbon, inorganic carbon, total phosphorus, pH, and total nitrogen were determined. Other physical and chemical indicators were determined as well.
The organic carbon content of soil samples was determined using the potassium dichromate oxidation method [42].
Soil organic carbon (SOC) storage at 0–100 cm (SOCS):
where k indicates different soil layers; Ck indicates the soil organic content in each soil layer; dk indicates the bulk density of each soil layer; Dk indicates the thickness of each soil layer; and Sk indicates the plot area.
SOCS = ∑Ck × dk × Dk/Sk
Soil carbon sink of plantations (ΔSCS):
where SP indicates the artificial sand-fixing plantations; CK indicates the control plots for afforesting sand-fixing plantations.
ΔSOCS = (SOCS)SP − (SOCS)CK
Total organic carbon storage of plantations (TCS):
TCS = BCS + SOCS
Total organic carbon sink of plantations (ΔTCS):
where SP indicates the artificial sand-fixing plantations; CK indicates the control plot for afforesting sand-fixing plantations.
ΔTCS = (BCS + SOCS)SP − (BCS + SOCS)CK
The carbon sequestration rate (Mg C ha−1·yr−1) (SCSR) of the sand-fixing plantations was calculated by measuring the net carbon accumulation per unit divided by plantation age.
where SOCT is the SOC storage of the sand-fixing plantation after T years of planting; SOC0 is the SOC storage of the control plot; and T is the plantation age.
SCSR = (SOCT − SOC0)/T
2.5. Statistical Analysis
Matplotlib 3.5.3 was used for mapping. Statistical analyses were performed with SPSS 20.0 software (IBM, New York, NY, USA). One-way ANOVA with Tukey’s test on normalized data was used to assess significant differences among the different sand-fixing plantations. A T-test was used to compare the organic carbon storage of different sand-fixing plantations with or without enclosures or clay sand barriers, and that of the plantations of two different ages. Multiple comparisons were adopted for the comparison involving three variables (the comparison between two different sand-fixing plantations). We considered values of α ≤ 0.05 as significant and ≤0.01 as significant.
3. Results
3.1. The BC Storage and Sink in Different Sand-Fixing Plantations
The carbon stored in biomass (Biomass Carbon, or BC) in different sand-fixing plantations with 25-year age was analyzed (Table 1). The BC storage of T. ramosissima was significantly higher than that of C. korshinskii, H. ammodendron, C. mongolicum, A. desertorum and H. scoparium. The BC storage of C. korshinskii and H. ammodendron were significantly higher than that of C. mongolicum, A. desertorum and H. scoparium. A. desertorum had the lowest total biomass carbon storage in all sand-fixing plantations. Additionally, significant differences were present in the above-ground BC storage and the below-ground BC storage among the six sand-fixing plantations. The carbon storage in the litters of six sand-fixing plantations was very low, and there were no significant differences in carbon stored in litter among the six plantations.

Table 1.
The BC storage of the sand-fixing plantations and the control plots in the Shiyang River Basin (Mg C ha−1).
Compared to any of the unplanted controls, sand-fixing afforestation significantly increased the carbon storage of above-ground, below-ground and total biomass, with the exception of A. desertorum and H. scoparium plantations (Table 1).
The BC storage of stable natural N. tangutorum shrubs was different from that of sand-fixing plantations (Table 1). The above-ground and total BC storage of the stable natural N. tangutorum shrubs were significantly lower than those of planted T. ramosissima, H. ammodendron and C. korshinskii, but were significantly higher than those of C. mongolicum and A. desertorum. In addition, the below-ground BC storage of the stable natural N. tangutorum was significantly higher than that of other sand-fixing plantations with the exception of T. ramosissima, H. ammodendron and C. korshinskii.
The BC sink of different sand-fixing plantations with a 25-year age was different (Figure 2). The BC sink of T. ramosissima was significantly higher than that of other sand-fixing plantations. The BC sequestration rate of T. ramosissima, H. ammodendron, C. korshinskii, H. scoparium, C. mongolicum and A. desertorum reached 1.47, 0.35, 0.42, 0.06, 0.05 and 0.03 Mg C ha−1·yr−1, respectively.
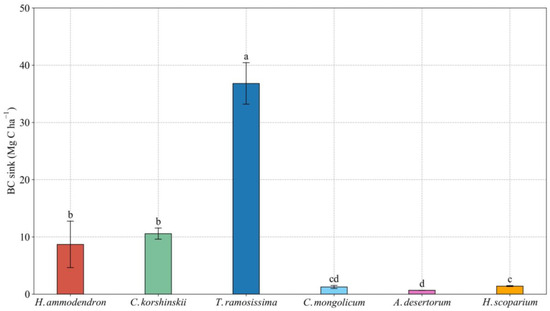
Figure 2.
BC sink of different sand-fixing plantations with a 25-year age. Values are means ± standard deviations (SD, n = 9). Different lowercase letters denote significant differences among different sand-fixing plantations at the 0.05 level (p < 0.05).
3.2. The SOC Storage and Sink in Different Sand-Fixing Plantations
In the Shiyang River Basin, the SOC storage in the T. ramosissima sand-fixing plantations was higher than that of other plantations with 25-year age, which was 9.4, 5.5, 3.0, 2.6, 1.5 times that of A. desertorum, C. mongolicum, H. scoparium, H. ammodendron and C. korshinskii, respectively (Figure 3). When compared to controls, significant difference was not present in SOC storage between the T. ramosissima plantation and the sand-covered salinized land control, and between C. korshinskii and inter-dune land control. Significant differences were present in SOC storage between the H. ammodendron, C. mongolicum and H. scoparium plantations and the mobile sand dune control. SOC storage in the T. ramosissima plantations with a 25-year age was similar to the stable native N. tangutorum shrub vegetation. SOC storage in the H. ammodendron, H. scoparium, C. mongolicum and A. desertorum plantation was significantly lower than that in the stable native N. tangutorum shrub vegetation.

Figure 3.
SOC storage of the different sand-fixing plantations at the layer of 0–100 cm. Values are means ± standard deviations (SD, n = 9). Different lowercase letters denote significant difference among different sand-fixing plantations at the 0.05 level (p < 0.05).
The SOC sink in the H. ammodendron plantation was significantly higher than that in the other sand-fixing plantations, which was 12.6, 2.9, 2.5, 2.3, and 1.2 times that of A. desertorum, C. mongolicum, T. ramosissima, C. korshinskii, and H. scoparium, respectively (Figure 4). The SOC sink in the A. desertorum plantation was significantly lower than that in the other sand-fixing plantations. Significant differences were not present in estimations of SOC sink capacity among C. korshinskii, T. ramosissima, and C. mongolicum plantations.
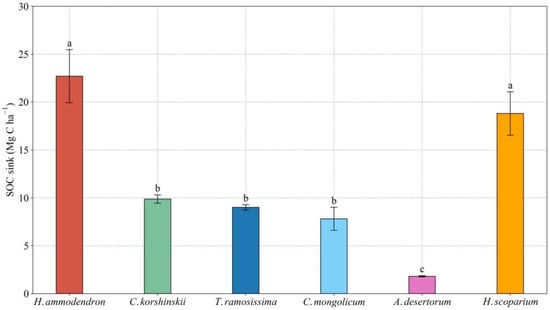
Figure 4.
SOC sink of different sand-fixing plantations. Values are means ± standard deviations (SD, n = 9). Different lowercase letters denote significant differences among different sand-fixing plantations at the 0.05 level (p < 0.05).
3.3. TC Storage and Sink in the Different Sand-Fixing Plantations
Total carbon (TC) storage in the T. ramosissima sand-fixing plantation was significantly higher than that in the C. korshinskii, H. ammodendron, C. mongolicum, A. desertorum and H. scoparium plantations, and significantly higher than that in the control plots. The TC storage in the A. desertorum plantation was lowest, only reaching 8.86 Mg C ha−1, which occupied 7.8%, 14.5%, 23.0%, 32.9%, 57.8% that of T. ramosissima, C. korshinskii, H. ammodendron, H. scoparium, and C. mongolicum plantation, respectively (Figure 5). Only TC storage in the T. ramosissima plantations was significantly higher than that in the native stable N. tangutorum vegetation. TC storage in the H. ammodendron, C. korshinkii, C. mongolicum, A. desertorum, and H. scoparium plantations was significantly lower than TC storage in the native stable N. tangutorum vegetation.

Figure 5.
TC storage of vegetation–soil system in the different sand-fixing plantations. Values are means ± standard deviations (SD, n = 9). Different lowercase letters denote significant differences among different sand-fixing plantations at the 0.05 level (p < 0.05).
The estimated TC sink in the T. ramosissima plantation reached 45.8 Mg C ha−1, which was significantly higher than that of the other sand-fixing plantations (Figure 6). Carbon sequestration rates in the A. desertorum plantation with a 25-year age only reached 0.07 Mg C ha−1·yr−1. The estimated TC sink in the A. desertorum plantation was the smallest of the plantation types, only being 3.8%, 5.5%, 8.6%, 8.7%, and 21.4% that of T. ramosissima, H. ammodendron, C. korshinskii, H. scoparium, and C. mongolicum plantation, respectively.
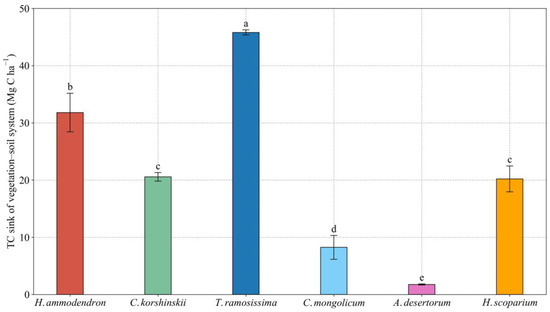
Figure 6.
TC sink of vegetation–soil system in the different sand-fixing plantations with 25-year age. Values are means ± standard deviations (SD, n = 9). Different lowercase letters denote significant differences among different sand-fixing plantations at the 0.05 level (p < 0.05).
The proportion of SOC storage to TC storage of the different sand-fixing plantations was much lower than that in any of the unplanted controls and stable natural N. tangutorum shrubs, excepted for A. desertorum and H. scoparium (Figure 7). The proportion of SOC storage to TC storage was different among different plant species (Figure 7). That in the H. ammodendron and T. ramosissima plantations were significantly lower than that in the other sand-fixing plantations, reaching 76.44% and 68.02%, respectively.

Figure 7.
The proportion of SOC storage to TC storage of the different sand-fixing plantations.
3.4. Effects of Management Measures on BC and SOC Storage in the Typical Sand-Fixing Plantations
Sand barriers need to be built on mobile sand dunes before H. ammodendron is planted. Sand barrier type significantly affected organic carbon storage in the sand-fixing plantations. BC storage in the H. ammodendron plantation with wheat–straw–sand barrier reached 12.3 Mg C ha−1, which was significantly higher than that of BC storage in a similar site with a clay sand barrier. SOC and TC storage in the H. ammodendron plantation with a wheat–straw–sand barrier reached 12.3 and 24.6 Mg C ha−1, respectively, each of which was significantly lower than that of a plantation constructed with a clay sand barrier (Figure 8).
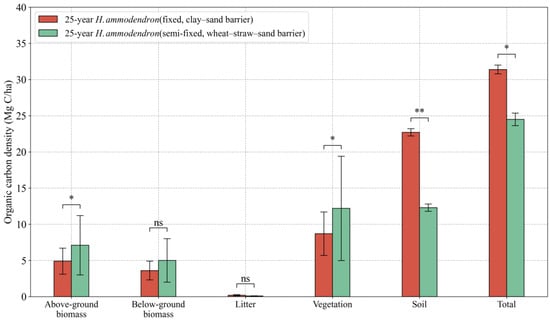
Figure 8.
Effects of plantation modes (clay–sand barrier or wheat–straw–sand barrier) on the allocation of each C sink in H. ammodendron ecosystem. Values are means ± standard deviations (SD, n = 9). Asterisks denote statistically significant differences * p < 0.05, ** p < 0.01.
H. scoparium plantations may have an enclosure set up around them to dissuade local herbivores and protect plants. In the Shiyang River Basin, the presence of an enclosure to reduce grazing significantly affected the organic carbon storage of H. scoparium plantations. BC storage in the H. scoparium plantation in semi-fixed sand dunes with non-enclosure reached 2.9 Mg C ha−1, which was significantly higher than that of plantations which were enclosed to prevent grazing. In fixed sand dunes, SOC and TC storage in the H. scoparium plantation protected from grazing by fencing were 18.8 and 20.2 Mg C ha−1, significantly higher than that of a site exposed to grazers (Figure 9).
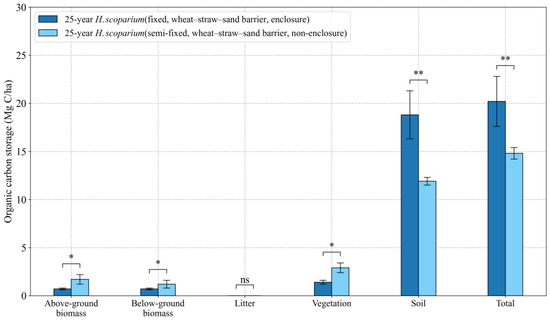
Figure 9.
Effects of enclosure on BC and SOC storage in H. scoparium plantation. Values are means ± standard deviations (SD, n = 9). An asterisk denotes statistically significant differences * p < 0.05, ** p < 0.01.
3.5. Effects of Plantation Age on BC and SOC Storage in Sand-Fixing Plantations
Considerably more organic carbon is stored in a 25-year H. ammodendron sand-fixing plantation than in a 3-year one (Figure 10). The organic carbon storage of above-ground biomass, below-ground biomass, vegetation, and soil were all much greater in the 25-year H. ammodendron sand-fixing than in the 3-year H. ammodendron sand-fixing plantation (Figure 10). However, we observed that the annual rate of carbon sequestration in above-ground biomass was lower in the 25-year plantation than in the 3-year plantation.
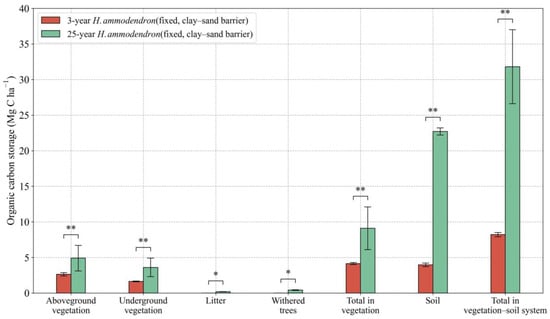
Figure 10.
Effects of plantation age on BC and SOC storage in the H. ammodendron ecosystem. Values are means ± standard deviations (SD, n = 9). * p < 0.05, ** p < 0.01.
The 3-year T. ramosissima plots observed in this study acted as carbon sources. For comparison, 25-year-old T. ramosissima plantations act as significant carbon sinks. Every carbon pool in the 25-year T. ramosissima sand-fixing plantations is much higher than that of the 3-year T. ramosissima sand-fixing plantations (Figure 11). Among them, the organic carbon storage of above-ground biomass, below-ground biomass, and total carbon storage in vegetation in the 25-year T. ramosissima plantations are 5.3, 6.3 and 6.4 times that of the 3-year T. ramosissima plantation, respectively (Figure 11).
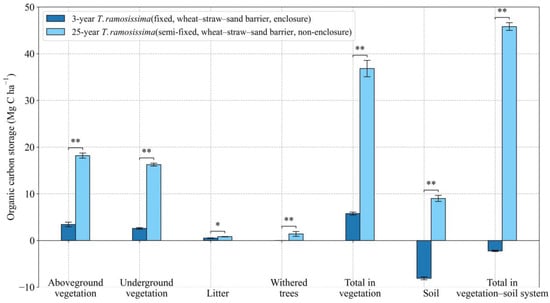
Figure 11.
Effects of plantation age on BC and SOC storage in T. ramosissima plantations. Values are means ± standard deviations (SD, n = 9). * p < 0.05, ** p < 0.01.
4. Discussion
4.1. The Comparisons of Carbon Storage Capacity with Other Sand-Fixing Plantations
The total organic carbon storage in the vegetation–soil system of six typical 25-year sand-fixing plantations ranged from 8.9 to 113.3 Mg C ha−1. The total organic carbon storage beneath T. ramosissima plantations was high (113.3 Mg C ha−1), near to observations in a similar Pinus sylvestris L. artificial forest within the Mu Us desert (98.86 Mg C ha−1) [44], and higher than that of both a Pinus halepensis Mill. plantation at the edge of the Negev desert (65 Mg C ha−1) [45], or a 10-year sand-fixing plantation mainly composed of Vitex negundo L. var. heterophylla and Hippohae rhamnoides (L.) A.Nelson. near Beijing (77.0 Mg C ha−1) [46].
The SOC storage of six typical 25-year sand-fixing plantations ranged from 8.2 to 77.1 Mg C ha−1, and in all settings, SOC was the main contributor to total carbon storage in sand-fixing plantations. However, even the maximum SOC storage of T. ramosissima was smaller than the average SOC storage of plantations in China (107.10 Mg C ha−1) [47] and major plantations in China (0–60 cm) with a value of 84.5 Mg C ha−1 [48]. Generally speaking, due to the dry climate and extremely poor hydrothermal conditions, the biomass carbon storage and soil carbon storage of sand-fixing plantations in arid desert are quite low [49]. However, the ecosystem carbon storage of T. ramosissima sand-fixing plantation (113.3 Mg C ha−1) exceeds that of the native N. tangutorum shrub ecosystem (92.1 Mg C ha−1), suggesting some plantation species may improve the ecosystem carbon storage potential of these desert ecosystems [50]. Thus, it can be concluded that the sand-fixing plantations in western China have certain carbon storage and have a great potential for carbon sequestration since drylands are representative of much of the region.
In this study, the T. ramosissima sand-fixing plantation has the highest vegetation carbon storage, SOC storage and total organic carbon storage of the vegetation–soil system, and the total ecosystem carbon sink increase rate of the six typical sand-fixing plantations. Thus, it can be concluded that T. ramosissima is the best choice for carbon sequestration in the arid region. We also observed that the vegetation carbon storage capacity of H. ammodendron and C. korshinskii is relatively higher than H. scoparium and A. desertorum. These differences are likely the consequence of physiological differences among the shrub species, T. ramosissima and H. ammodendron have tall trunks, dense branches, and deep and developed root systems. The large number of assimilating stems is a superior adaption to survive in arid regions [51]. It is unsurprising these two species express greater vegetation biomass in comparison with the short and sparse H. scoparium and A. desertorum. We observed that biomass carbon storage in H. ammodendron and T. ramosissima was roughly one-third of total carbon storage, suggesting that the ideal carbon sink candidate requires large above-ground biomass expression (Figure 7). This is consistent with former research which reported that biomass average carbon storage determines the total organic carbon storage of desert ecosystems [11].
It is worth noting, however, that our results are inconsistent with other research in the Karamay [52], which reported that H. ammodendron has the most carbon storage (0.21 Mg C ha−1) and T. ramosissima has the least carbon storage (0.01 Mg C ha−1). Despite the influence of vegetation, in desert ecosystems, most carbon is stored in the soil, and 67.6%–85.0% of the total carbon pool is SOC [53]. In this study, the T. ramosissima sand-fixing plantation has the highest vegetation carbon sink and total organic carbon sink of the vegetation–soil system of the six typical sand-fixing plantations. However, its estimated potential SOC sink is only ranked third among the six typical sand-fixing plantations, lower than that of either H. ammodendron or C. korshinskii sand-fixing plantations. Moreover, we observed that A. desertorum, C. mongolicum and H. scoparium sand-fixing plantation ecosystems have a lower proportion of vegetation organic carbon than that of T. ramosissima, C. korshinskii, and H. ammodendron, likely due the relatively shorter height, sparser branches and less developed root system [11,53]. The relatively high contribution of SOC stocks in these ecosystems may be because desert areas are characterized by low rainfall, high atmospheric evaporation, and a deep groundwater table. Most shrub sand-fixing plantations experience severe water shortages and have corresponding special water-conservation strategies [54,55] to exploit groundwater and water from deep soil layers [56]. This water stress may explain the success of T. ramosissima relative to its alternate species. The species is known for fast root growth and a greater hydraulic safety margin due to physiological adaptations that facilitate access to deep groundwater [57,58].
T. ramosissima has been shown to deliberately increase its water use efficiency first and decrease its growth demands until after water access has been satisfied [59]. The deep roots help T. ramosissima use groundwater reserves to resist drought during the dry period [60,61,62]. Unsurprisingly, this large root carbon allocation accounts for 27.53% of the total carbon storage of T. ramosissima [48]. We present this because fine root turnover to below-ground soil organic carbon is considered a major component of soil organic carbon, especially in the short term [63]. The fine root turnover of T. ramosissima happens 1.98 times every year [48], and this higher fine root turnover rate can partly explain the higher SOC, which has been attributed to facilitating the survival of plants under drought stress [64,65]. Taken together, these results suggest that T. ramosissima may demonstrate such improved carbon storage capacity due to its ability to adapt and weather drought–particularly given the intense water demand associated with the region.
In the present study, H. ammodendron showed a stronger soil carbon sequestration capacity than any other species. Afforestation efforts using H. ammodendron can increase the proportion of mesoaggregates in the soil and increase soil organic carbon [66]. In arid areas, H. ammodendron has been associated with a higher fraction of aggregate-associated soil organic carbon and aggregate-associated microbial biomass carbon compared to soils beneath T. ramosissima, Karelinia caspia (Pall.) Less., or Bassia dasyphylla (Fisch. & C. A. Mey.) Kuntze community [67]. However, we observed the soil carbon sequestration capacity of H. ammodendron was lower than that of C. korshinskii. As a nitrogen (N)-fixing leguminous plant, C. korshinskii may increase the net accumulation of microbial carbon in soil by reducing N limitation and/or increasing carbon inputs belowground, each thus increasing the relative abundance of microbes and MBC [68,69]. Furthermore, C. korshinskii is considered the optimal dominant species for the reconstruction in dryland regions given its high instantaneous water use efficiency and its organic carbon storage within the branches and leaves in arid regions [12,70,71]. The planting of C. korshinskii could enhance the carbon sequestration capacity through humus formation by altering the size as well as structure of the soil microbial community [72]. The SOC sink of C. korshinskii increased by 9.87 Mg C ha−1 compared to the inter-dune land in this study, which is higher than the value (1.40–3.19 Mg C ha−1) of the same plant reported in the semiarid Loess Hilly Region [73]. Under the “Grain for Green” Program, the total ecosystem carbon sink increase rate of the C. korshinskii plantations is 1.33 Mg ha−1 yr−1 from the young stage to the mature stage [74]. In this study, the total ecosystem carbon sink increase rate of the C. korshinskii plantations is 0.82 Mg ha−1 yr−1. The difference may be attributed to the large difference in soil moisture between study areas.
4.2. Main Factors on Carbon Storage and Sink of Sand-Fixing Plantations
There are many factors affecting the carbon storage and carbon sequestration of sand-fixing plantation ecosystems, including sand dune type, afforestation measures (sand barriers), presence/absence of grazing species, or time since planting (plantation age) [75]. The types of sand dunes atop which sand-fixing plantations were planted affect the carbon sequestration capacity of the eventual plantation ecosystem. In this study, we observed lower rates of carbon storage in fixed-sand land than semi-fixed, though more organic carbon was stored beneath semi-fixed sand lands than fixed sand lands. This is largely due to the formation of soil crusts in fixed lands, which inhibit evapotranspirative losses and consequently reduce the amount of water that returns to the soil every rain event. This water deficit likely harms deep-rooted shrubs like H. ammodendron and H. scoparium, reducing their above-ground biomass expression and consequently their potential to store organic carbon.
Moreover, afforestation measures (sand barriers) affect the carbon sequestration capacity of the sand-fixing plantation ecosystems. Clay content in the desert soil is quite low [76,77]. In the current study, we found that the use of a clay–sand barrier during planting may increase reserves of soil organic matter, facilitating the development of H. ammodendron sand-fixing plantations. The clay from interdune lowlands, with fine soil particles and rich organic substance, was used for such a barrier, which tended to form a crust on the sand surface, and was conducive to soil development. This could explain why the organic carbon storage of the total ecosystem and soil was much higher in the H. ammodendron sand-fixing plantations planted with clay–sand barrier. Moreover, the clay–sand barrier may accelerate the formation of abundant biological soil crusts [78], which in turn capture more fine soil particles and improve soil carbon sequestration capacity [79,80]. This is further verification that the clay content of the soil is closely related to the total organic carbon storage of the desert ecosystem in arid areas.
Management measures can largely affect the carbon sequestration capacity of the sand-fixing plantation ecosystems [81]. Improving the management of plantations to enhance the potential for a soil carbon sink is a promising option for enhancing the carbon sink function of terrestrial ecosystems to mitigate global climate change [82,83,84,85,86]. Enclosure (building fences to keep out opportunist grazers) is one management choice that can increase plant growth, species richness, and plant density, promoting the restoration of degraded sand-fixing shrub vegetation [87,88]. This is consistent with other observations [37]. Enclosed sand-fixing plantations are undisturbed by humans or animals, promoting the development of crust and eventually soil [37]. This can explain why there are higher levels of SOC beneath enclosed plantations. In addition, the contribution of soil organic carbon storage to overall carbon storage is around 68.0%–94.8% (Figure 7), hence the total carbon storage is still higher in the enclosed sand-fixing plantations (Figure 9). Of course, the BC storage in the unenclosed sand-fixing plantations is much higher than that of the enclosed sand-fixing ones, mainly due to the fact that when there is no fence, human interference damages the crust, promotes soil moisture infiltration and the growth of deep-rooted shrubs, thus increases the aboveground biomass carbon (Figure 9).
Plantation age is another factor that affects the carbon sequestration capacity of the sand-fixing plantations. Older plantations store more carbon than younger plantations, but their annual carbon storage rate saturates as growth saturates and the population reaches its climax size [89]. This result is consistent with most studies on plantations [90]. In this study, young plantations have lower carbon sinks, and may even be carbon sources, depending on plant species. As the plantation ages, the carbon sequestration storage increases which is consistent with other studies [7,11,91,92]. This pattern of saturating increase is reflected in the soil carbon as well, and Zhang et al. (2016) stated that the accumulation rates of SOC in an age sequence of H. ammodendron plantations is faster in the early stages (0–9 years) and becomes slower in the late stages (9–39 years) [93]. However, it is worth considering that Ma et al. (2021) reports the carbon sequestration capacity of the H. ammodendron plantations began to slowly decline after the 25-year mark and then rapidly decline after 50 years [11]. Similarly, Wang (2020) suggested that the planted sand-fixing vegetation of the Shiyang River Basin will begin to emit carbon around 80 years after planting [49]. This bell-shaped pattern of the carbon sink capacity is associated with the reduction in groundwater and soil water content as plants reach their climax state and begin to degrade. The carbon sequestration characteristics of T. ramosissima are different from those of other tree species, which may be related to its biological characteristics. T. ramosissima prefers saline-alkali and humid soil, and its afforestation site has saline soil covered by sand. Its deep soil has high organic carbon storage, and afforestation and land preparation cause carbon emissions, producing no carbon sink effect in the initial year, which is different from other species planted on mobile sand dunes.
These results are consistent with what we know from sustainable plantation management designed to reach carbon neutrality in plantations. Scientists often harvest and revegetate the land or adopt thinning measures designed to keep plantations at peak carbon storage capacity. Similarly, scientists have experimented to identify peak management intensities and stand ages for a variety of plantation species. For example, Li et al. (2022) reported that the carbon sequestration capacity of the 15-year C. korshinskii plantation reached its highest stocks of SOC and so it should be harvested and revegetated every 15 years to achieve the largest accumulations of soil carbon [58]. In the water-limited areas Loess Plateau, Deng et al. (2017) found that C. korshinskii plantation should be harvested and revegetated at 35-year intervals [74]. One potential management strategy to keep in mind might consider Chai et al. (2019) who reported a decrease (23.9%) of soil organic carbon (0–100 cm) when plantations aged from 10 to 30 years and asserted that SOC sequestration optimization must consider thinning management [94].
5. Conclusions
To obtain the ambitious Chinese goals of carbon neutrality, it is vital to reconstruct the “New Three-North”. Thus, it is urgent to fully grasp the characteristics and mechanisms of carbon sequestration and to know the scientific planning of artificial sand-fixing plantations in vast desert areas. In this study, we used the plot method to clarify carbon sequestration characteristics, such as capacity, main influencing factors, and the distribution of carbon sequestration of six typical sand-fixing plantations in arid desert areas in northwest China.
Ultimately, we evaluated the carbon sequestration capacity of major sand-fixing plantations and found the main influencing factors. Beyond that, it was found that afforestation using a clay–sand barrier is better than a wheat–straw–sand one in terms of carbon sequestration. Similarly, conservation and management do influence the carbon storage potential of those planted ecosystems, and we observed that enclosed plantations had improved carbon sequestration storage relative to the unenclosed. Moreover, plantation age also impacted carbon sequestration, and older plantations store more carbon than younger ones. At last, compared to control sites, we confirmed that soil carbon remains the priority when considering the carbon storage capacity of desert ecosystems.
Although great efforts were made to compare and analyze the carbon sequestration capacity of six typical sand-fixing plantations using the plot method, it is difficult to obtain accurate information regarding the variations in forest management and soil characteristics in desert ecosystems. Most of the vegetation data in this study were collected from sand-fixing plantations and hence site preparation and plot choices may increase the uncertainty about estimating the carbon sequestration of plantations. In addition, there may be a nonlinear relationship between plantation age and carbon storage potential, as older plantations begin to reduce soil water resources and degrade. Moreover, the changes in dynamic storage were not taken into account, which may increase the uncertainty as well. In the long run, more plant species need to be compared to better comprehend the carbon sequestration of sand-fixing plantations in desert areas. The findings are informative for sand-fixing planting and forest management in arid and semiarid regions, offering valuable insights into the scientific afforestation planning in reconstructing the “New Three-North”.
Author Contributions
Q.M.: conceptualization of the project. Q.M., L.W., F.C., D.Z. and H.J.: investigation. X.W.: writing—original draft preparation. X.W. and Q.M.: data analysis and visualization. X.W. and Q.M.: writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No.32160410), Forest Ecosystem Carbon Stock Baseline Survey and Carbon Sink Assessment Project in Gansu Province, Key Talent Projects in Gansu Province (2022 RCXM039), Central Finance Forestry Science and Technology Promotion Demonstration Project (2021 ZYTG002), the National Social Science Foundation of China (No.19 BMZ141), the Joint Research Program of the Alliance of Open Universities in the Yellow River Basin (2022) (HHLMKT202206), and the Research Project on Carbon Storage of Artificial Sand Fixation Forest in Gansu Province.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
Thanks should go to the anonymous reviewers for their valuable comments for improving this manuscript. We would like to thank Joseph Elliot at the University of Kansas for his assistance with English language and grammatical editing of the manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Diffenbaugh, N.S.; Burke, M. Global warming has increased global economic inequality. Proc. Natl. Acad. Sci. USA 2019, 116, 9808–9813. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.R.; Hao, T.X.; Zhu, J.X. Discussion on action strategies of China’s carbon peak and carbon neutrality. Bull. Chin. Acad. Sci. 2022, 37, 423–434. (In Chinese) [Google Scholar]
- Seth, P.; Sebastian, J. Plants and global warming: Challenges and strategies for a warming world. Plant Cell Rep. 2024, 43, 27. [Google Scholar] [CrossRef]
- Schleussner, C.F.; Lissner, T.K.; Fischer, E.M.; Wohland, J.; Perrette, M.; Golly, A.; Rogelj, J.; Childers, K.; Schewe, J.; Frieler, K.; et al. Differential climate impacts for policy-relevant limits to global warming: The case of 1.5 °C and 2 °C. Earth Syst. Dyn. 2016, 7, 327–351. [Google Scholar] [CrossRef]
- Rogelj, J.; den Elzen, M.; Höhne, N.; Fransen, T.; Fekete, H.; Winkler, H.; Schaeffer, R.; Sha, F.; Riahi, K.; Meinshausen, M. Paris Agreement climate proposals need a boost to keep warming well below 2 °C. Nature 2016, 534, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.b.; Zhou, S.; Jiang, K.J.; Bertram, C.; Harmsen, M.; Kriegler, E.; van Vuuren, D.P.; Wang, S.Y.; Fujimori, S.; Tavoni, M.; et al. Assessing China’s efforts to pursue the 1.5 °C warming limit. Science 2021, 372, 378–385. [Google Scholar] [CrossRef]
- Ma, Q.L.; Wang, Y.L.; Li, Y.K.; Sun, T.; Milne, E. Carbon storage in a wolfberry plantation chronosequence established on a secondary saline land in an arid irrigated area of Gansu Province, China. J. Arid Land 2018, 10, 202–216. [Google Scholar] [CrossRef]
- Diao, J.J.; Liu, J.X.; Zhu, Z.L.; Wei, X.Y.; Li, M.S. Active forest management accelerates carbon storage in plantation forests in Lishui, southern China. For. Ecosyst. 2022, 9, 33–46. [Google Scholar] [CrossRef]
- Ma, W.; Wang, Y. Optimizing China’s afforestation strategy: Biophysical impacts of afforestation with five locally adapted forest types. Forests 2024, 15, 182. [Google Scholar] [CrossRef]
- Loni, A.; Radnezhad, H.; Martynova-van kley, A.; Hassanvand, A.; Sadeghi, M.; Zaremanesh, H. The role of haloxylon plantations in improving carbon sequestration potential of sand dunes of iran. Appl. Ecol. Env. Res. 2018, 16, 321–333. [Google Scholar] [CrossRef]
- Ma, Q.L.; Wang, X.Y.; Chen, F.; Wei, L.Y.; Zhang, D.K.; Jin, H.J. Carbon sequestration of sand-fixing plantation of Haloxylon ammodendron in Shiyang River Basin: Storage, rate and potential. Glob. Ecol. Conserv. 2021, 28, e01607. [Google Scholar] [CrossRef]
- Yu, S.; Yang, J.; Norghauer, J.M.; Yang, J.; Yang, B.; Zhang, H.; Li, X. Soil carbon and nitrogen stocks and their influencing factors in different-aged stands of sand-fixing Caragana korshinskii in the Mu Us Desert of northwest China. Forests 2024, 15, 1018. [Google Scholar] [CrossRef]
- FAO. Land Resources Potential and Constraints at Region and Country Levels; World Soil Resources Reports 90; Land and Water Development Division, FAO: Rome, Italy, 2000. [Google Scholar]
- Ardö, J.; Olsson, L. Assessment of soil organic carbon in semi-arid Sudan using GIS and the CENTURY model. J. Arid Environ. 2003, 54, 633–651. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Mohammadi, F.; Fakhireh, A.; Bameri, A. Effects of Haloxylon spp. of different age classes on vegetation cover and soil properties on an arid desert steppe in Iran. Pedosphere 2019, 29, 619–631. [Google Scholar] [CrossRef]
- Suganuma, H.; Aikawa, S.; Sakai, Y.; Hamano, H.; Takahashi, N.; Tahara, K.; Kawarasaki, S.; Utsugi, H.; Egashira, Y.; Kawanishi, T.; et al. Estimation of CO2 sequestration potential by afforestation in the arid rangelands of western Australia based on long-term empirical data. Ecol. Eng. 2019, 133, 109–120. [Google Scholar] [CrossRef]
- El Kateb, H.; Zhang, H.F.; Abdallah, Z. Volume, biomass, carbon sequestration and potential of desert lands’ afforestation irrigated by wastewater on examples of three species. For. Ecol. Manag. 2022, 504, 119827. [Google Scholar] [CrossRef]
- Lu, Y.X.; Dai, L.L.; Yan, G.X.; Huo, Z.T.; Chen, W.; Lan, J.Z.; Zhang, C.M.; Xu, Q.Y.; Deng, S.M.; Chen, J. Effects of various land utilization types on groundwater at different temporal scales: A case study of Huocheng plain, Xinjiang, China. Front. Environ. Sci. 2023, 11, 1225916. [Google Scholar] [CrossRef]
- Gao, Y.; Tian, J.; Pang, Y.; Liu, J.B. Soil inorganic carbon sequestration following afforestation is probably induced by pedogenic carbonate formation in northwest China. Front. Plant Sci. 2017, 8, 1282. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.F.; Zhang, W.J.; Zhang, Z.Y.; Wang, L.; Ren, H.R.; Jiang, Y.; Zhang, X.S. Responses of soil C stock and soil C loss to land restoration in Ili River Valley, China. Catena 2018, 171, 469–474. [Google Scholar] [CrossRef]
- Yang, Y.H.; Chen, Y.N.; Li, Z.; Chen, Y.P. Land-use/cover conversion affects soil organic-carbon stocks: A case study along the main channel of the Tarim River, China. PLoS ONE 2018, 13, e0206903. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Chen, A.P. Contrasting responses of soil inorganic carbon to afforestation in acidic versus alkaline soils. Glob. Biogeochem. Cycles 2022, 36, e2021GB007038. [Google Scholar] [CrossRef]
- Li, P.; Li, H.J.; Si, B.C.; Zhou, T.; Zhang, C.H.; Li, M. Mapping planted forest age using LandTrendr algorithm and Landsat 5–8 on the Loess Plateau, China. Agric. For. Meteorol. 2024, 344, 109795. [Google Scholar] [CrossRef]
- Wang, X.Y.; Ma, Q.L.; Wang, Y.L. Carbon benefits evaluation of the artificial shelter forest in the Shiyanghe River Basin. J. Desert Res. 2020, 40, 197–205. (In Chinese) [Google Scholar]
- Yer Çelik, E.N.; Ayan, S.; Özel, H.B.; Turfan, N.; Mehmet Yer, B.; Abdaloğlu, G. Effects of melatonin applications on Anatolian black pine (Pinus nigra J. F. Arnold. subsp. pallasiana (Lamb.) Holmboe) afforestation performance in semi-arid areas. BioResources 2023, 18, 2551–2573. [Google Scholar] [CrossRef]
- Bauhus, J.; van der Meer, P.; Kanninen, M. Ecosystem Goods and Services from Plantation Forests; Routledge Press: London, UK, 2010. [Google Scholar]
- Tsaktsira, M.; Tsoulpha, P.; Economou, A.; Scaltsoyiannes, A. Mitigation of global climate change through genetic improvement of resin production from resinous pines: The case of pinus halepensis in greece. Sustainability 2023, 15, 8052. [Google Scholar] [CrossRef]
- Nosetto, M.D.; Jobbágy, E.G.; Paruelo, J.M. Carbon sequestration in semi-arid rangelands: Comparison of Pinus ponderosa plantations and grazing exclusion in NW Patagonia. J. Arid Environ. 2006, 67, 142–156. [Google Scholar] [CrossRef]
- Xu, Z.H.; Wei, H.J.; Dong, X.B.; Liu, Y.X.; Xue, H.L. Evaluating the ecological benefits of plantations in arid areas from the perspective of ecosystem service supply and demand-based on emergy analysis. Sci. Total Environ. 2020, 705, 135853. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Shao, M.A.; Zhu, Y.J.; Liu, Z.P. Impacts of land use and plant characteristics on dried soil layers in different climatic regions on the Loess Plateau of China. Agric. For. Meteorol. 2011, 151, 437–448. [Google Scholar] [CrossRef]
- Wang, C.; Liu, S.G.; Zhu, Y.; Smith, A.R.; Ning, Y.; Deng, D.M. Aboveground carbon sequestration of Cunninghamia lanceolata forests: Magnitude and drivers. For. Ecosyst. 2024, 11, 100165. [Google Scholar] [CrossRef]
- Singh, G.; Mishra, D.; Singh, K.; Shukla, S.; Choudhary, G.R. Geographical settings and tree diversity influenced soil carbon storage in different forest types in Rajasthan, India. Catena 2022, 209, 105856. [Google Scholar] [CrossRef]
- Bazgir, M.; Omidipour, R.; Heydari, M.; Zainali, N.; Hamidi, M.; Dey, D.C. Prioritizing woody species for the rehabilitation of arid lands in western Iran based on soil properties and carbon sequestration. J. Arid Land 2020, 12, 640–652. [Google Scholar] [CrossRef]
- Liu, C.A.; Siddique, K.H.M.; Hua, S.; Rao, X. The trade-off in the establishment of artificial plantations by evaluating soil properties at the margins of oases. Catena 2017, 157, 363–371. [Google Scholar] [CrossRef]
- Hbirkou, C.; Martius, C.; Khamzina, A.; Lamers, J.P.A.; Welp, G.; Amelung, W. Reducing topsoil salinity and raising carbon stocks through afforestation in Khorezm, Uzbekistan. J. Arid Environ. 2011, 75, 146–155. [Google Scholar] [CrossRef]
- Li, Y.Q.; Brandle, J.; Awada, T.; Chen, Y.P.; Han, J.J.; Zhang, F.X.; Lou, Y.Q. Accumulation of carbon and nitrogen in the plant–soil system after afforestation of active sand dunes in China’s Horqin Sandy Land. Agric. Ecosyst. Environ. 2013, 177, 75–84. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, X.Y.; Wang, S.K.; Zhang, F.X.; Lian, J.; Huang, W.D.; Mao, W. Carbon accumulation in the bulk soil and different soil fractions during the rehabilitation of desertified grassland in horqin sandy land (Northern China). Pol. J. Ecol. 2015, 63, 88–101. [Google Scholar] [CrossRef]
- Liu, J.B.; Wu, P.X.; Zhao, Z.; Gao, Y. Afforestation on cropland promotes pedogenic inorganic carbon accumulation in deep soil layers on the Chinese loess plateau. Plant Soil 2022, 478, 597–612. [Google Scholar] [CrossRef]
- Lu, Q.; Xiao, C.L.; Bao, Y.S.; Cui, M.C.; Cao, X.M.; Que, X.E.; Yang, L.; Cui, G.P. Implementation path and strategic planning of winning the battle of “Three-North” and reconstructing “New Three-North”. Bull. Chin. Acad. Sci. 2023, 38, 956–965. [Google Scholar]
- Wu, D.X.; Wei, W.S.; Zhang, S.M. Protocol for Standard Biological Observation and Measurement in Terrestrial Ecosystem; China Environmental Sciences Press: Beijing, China, 2007; pp. 46–75. (In Chinese) [Google Scholar]
- Selyaninov, G.T. About climate agricultural estimation. Proc. Agric. Meteorol. 1928, 20, 165–177. [Google Scholar]
- Soil Science Society of China. Soil Agrochemical Analysis Method; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Wang, S.Q.; Liu, J.Y.; Yu, G.R. Error analysis of estimating terrestrial soil organic carbon storage in China. Chin. J. Appl. Ecol. 2003, 14, 797–802. (In Chinese) [Google Scholar]
- Xu, S.; Liao, C.Y.; Duo, J.J.; Fu, G.J. Study of carbon storage and spatial distribution of Pinus sylvestris artificial forest under different slope aspects in Mu Us Desert. Res. Soil Water Conserv. 2015, 22, 14–18. (In Chinese) [Google Scholar]
- Grünzweig, J.M.; Lin, T.L.; Rotenberg, E.; Schwartz, A.; Yakir, D. Carbon sequestration in arid-land forest. Glob. Chang. Biol. 2003, 9, 791–799. [Google Scholar] [CrossRef]
- Liu, X.P.; Zhang, W.J.; Cao, J.S.; Yang, B.; Cai, Y.J. Carbon sequestration of plantation in Beijing-Tianjin sand source areas. J. Mt. Sci. 2018, 15, 2148–2158. [Google Scholar] [CrossRef]
- Liu, S.R.; Wang, H.; Luan, J.W. A review of research progress and future prospective of forest soil carbon stock and soil carbon process in China. Acta Ecol. Sin. 2011, 31, 5437–5448. (In Chinese) [Google Scholar]
- Wang, D.P.; Wang, W.B.; Zheng, L.; Luo, X.H.; Zou, B.X.; Zhang, Y.F.; Wu, X.P. Compared of soil organic carbon of different plantations in China. Ecol. Environ. Sci. 2014, 23, 698–704. (In Chinese) [Google Scholar]
- Wang, X.Y. A Study on the Process, Rate and Benefits of Carbon Sequestration of Planted Sand-Fixing Vegetation in Shiyang River Basin; Lanzhou University: Lanzhou, China, 2020. (In Chinese) [Google Scholar]
- Wang, X.Y.; Ma, Q.L.; Jin, H.J.; Fan, B.L.; Wang, D.B.; Lin, H.L. Change in characteristics of soil carbon and nitrogen during the succession of Nitraria tangutorum in an arid desert area. Sustainability 2019, 11, 1146. [Google Scholar] [CrossRef]
- Zhou, Z.J.; Su, P.X.; Wu, X.K.; Zhang, H.N.; Shi, R.; Yang, J.P. Leaf and canopy photosynthesis of four desert plants: Considering different photosynthetic organs. Photosynth. Res. 2022, 151, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.S.; Meikere, A.; Zhang, Y.T.; Chang, S.L.; Yi, H.F.; Zhao, F.S.; Zhang, W.Y. Research on carbon sinks of Poplar plantations in Karamay. Arid Zone Res. 2015, 32, 388–394. (In Chinese) [Google Scholar]
- Yang, H.T.; Li, X.R.; Wang, Z.R.; Jia, R.L.; Liu, L.C.; Chen, Y.L.; Wei, Y.P.; Gao, Y.H.; Li, G. Carbon sequestration capacity of shifting sand dune after establishing new vegetation in the Tengger Desert, northern China. Sci. Total Environ. 2014, 478, 1–11. [Google Scholar] [CrossRef]
- Brouillette, L.C.; Mason, C.M.; Shirk, R.Y.; Donovan, L.A. Adaptive differentiation of traits related to resource use in a desert annual along a resource gradient. New Phytol. 2014, 201, 1316–1327. [Google Scholar] [CrossRef]
- Wu, Y.S.; Liao, Z.L.; Yu, X.Q.; Yin, Q.; Wang, H.; Gao, L. Stable isotopic analysis of water utilization characteristics of four xerophytic shrubs in the Hobq Desert, Northern China. Front. Plant Sci. 2023, 14, 1103789. [Google Scholar]
- Li, E.G.; Tong, Y.Q.; Huang, Y.M.; Li, X.Y.; Wang, P.; Chen, H.Y.; Yang, C.Y. Responses of two desert riparian species to fluctuating groundwater depths in hyperarid areas of northwest China. Ecohydrology 2019, 12, e2078. [Google Scholar] [CrossRef]
- Xia, J.B.; Lang, Y.; Zhao, Q.K.; Liu, P.; Su, L. Photosynthetic characteristics of Tamarix chinensis under different groundwater depths in freshwater habitats. Sci. Total Environ. 2020, 761, 143221. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.J.; Sheng, Y.; He, X.R. Whole-plant water use and hydraulics of Populus euphratica and Tamarix ramosissima seedlings in adaption to groundwater variation. Water 2022, 14, 1869. [Google Scholar] [CrossRef]
- Imin, B.; Dai, Y.; Shi, Q.D.; Guo, Y.C.; Li, H.; Nijat, M. Responses of two dominant desert plant species to the changes in groundwater depth in hinterland natural oasis, Tarim Basin. Ecol. Evol. 2021, 11, 9460–9471. [Google Scholar] [CrossRef]
- Yu, T.F.; Feng, Q.; Si, J.H.; Xi, H.Y.; Li, Z.X.; Chen, A.F. Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China’s extremely arid region. Plant Soil 2013, 372, 297–308. [Google Scholar] [CrossRef]
- Gou, S.; Miller, G. A groundwater-soil-plant-atmosphere continuum approach for modeling water stress, uptake, and hydraulic redistribution in phreatophytic vegetation. Ecohydrology 2014, 7, 1029–1041. [Google Scholar] [CrossRef]
- Dong, Z.W.; Li, S.Y.; Zhao, Y.; Lei, J.Q.; Wang, Y.D.; Li, C.J. Stable oxygen-hydrogen isotopes reveal water use strategies of Tamarix taklamakanensis in the Taklimakan Desert, China. J. Arid Land 2020, 12, 115–129. [Google Scholar] [CrossRef]
- Lai, Z.R.; Zhang, Y.Q.; Liu, J.B.; Wu, B.; Qin, S.G.; Fa, K.Y. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. Forest Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Fogel, R. Root turnover and productivity of coniferous forests. Plant Soil 1983, 71, 75–85. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Van Rees, K.C.J. The growth and function of pine roots. Ecol. Bull. 1994, 43, 76–91. [Google Scholar]
- Zhong, Z.K.; Han, X.H.; Xu, Y.D.; Zhang, W.; Fu, S.Y.; Liu, W.C.; Ren, C.J.; Yang, G.H.; Ren, G.X. Effects of land use change on organic carbon dynamics associated with soil aggregate fractions on the Loess Plateau, China. Land Degrad. Dev. 2019, 30, 1070–1082. [Google Scholar] [CrossRef]
- Yang, H.C.; Wang, J.Y.; Zhang, F.H. Soil aggregation and aggregate-associated carbon under four typical halophyte communities in an arid area. Environ. Sci. Pollut. Res. 2016, 23, 23920–23929. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Jia, L.; Zhang, Y.M.; Mou, X.M.; Li, X.G. Leguminous Caragana korshinskii evidently enhances microbial necromass carbon accumulation in dryland soils. Catena 2022, 215, 106342. [Google Scholar] [CrossRef]
- Lu, Q.; Ma, H.B.; Zhou, Y.; Calvelo-Pereira, R.; Shen, Y. Caragana korshinskii Kom. Plantation reduced soil aggregate stability and aggregate-associated organic carbon on desert steppe. PeerJ 2022, 10, e12507. [Google Scholar] [CrossRef]
- Gong, C.M.; Bai, J.; Wang, J.H.; Zhou, Y.L.; Kang, T.; Wang, J.J.; Hu, C.X.; Guo, H.B.; Chen, P.L.; Xie, P.; et al. Carbon storage patterns of Caragana korshinskii in areas of reduced environmental moisture on the Loess Plateau, China. Sci. Rep. 2016, 6, 28883. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.F.; Liao, Z.Q.; Yao, G.Q.; Tian, X.Q.; Bi, M.H.; Da Silva, J.A.T.; Gao, T.P.; Fang, X.W. Divergent stem hydraulic strategies of Caragana korshinskii resprouts following a disturbance. Tree Physiol. 2022, 42, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Cheng, M.; Huang, Y.M.; An, S.S.; Darboux, F. Changes in soil microbial community and its effect on carbon sequestration following afforestation on the Loess Plateau, China. Int. J. Environ. Res. Public Health 2017, 14, 948. [Google Scholar] [CrossRef]
- Xu, H.W.; Qu, Q.; Wang, M.G.; Li, P.; Li, Y.Z.; Xue, S.; Liu, G.B. Soil organic carbon sequestration and its stability after vegetation restoration in the Loess Hilly Region, China. Land Degrad. Dev. 2020, 31, 568–580. [Google Scholar] [CrossRef]
- Deng, L.; Han, Q.S.; Zhang, C.; Tang, Z.S.; Shangguan, Z.P. Above-ground and below-ground ecosystem biomass accumulation and carbon sequestration with Caragana korshinskii kom plantation development. Land Degrad. Dev. 2017, 28, 906–917. [Google Scholar] [CrossRef]
- Piao, S.L.; He, Y.; Wang, X.H.; Chen, F.H. Estimation of China’s terrestrial ecosystem carbon sink: Methods, progress and prospects. Sci. China Earth Sci. 2022, 65, 641–651. [Google Scholar] [CrossRef]
- Lowery, B.; Swan, J.B.; Schumacher, T.E.; Jones, A.J. Physical properties of selected soils by erosion class. J. Soil Water Conserv. 1995, 50, 306–311. [Google Scholar]
- Saxton, K.E.; Rawls, W.J. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef]
- Niu, J.P.; Yang, K.; Tang, Z.J.; Wang, Y.T. Relationships between soil crust development and soil properties in the desert region of North China. Sustainability 2017, 9, 725. [Google Scholar] [CrossRef]
- Zhao, H.L.; Guo, Y.R.; Zhou, R.L.; Drake, S. The effects of plantation development on biological soil crust and topsoil properties in a desert in northern China. Geoderma 2011, 160, 367–372. [Google Scholar] [CrossRef]
- Fan, B.L.; Zhang, A.P.; Yang, Y.; Ma, Q.L.; Li, X.M.; Zhao, C.M. Long-term effects of xerophytic shrub Haloxylon ammodendron plantations on soil properties and vegetation dynamics in northwest China. PLoS ONE 2016, 11, e0168000. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Li, H. Improving carbon sequestration capacity of forest vegetation in China: Afforestation or forest management? Forests 2023, 14, 1077. [Google Scholar] [CrossRef]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Law, B.E.; Berner, L.T.; Mildrexler, D.J.; Bloemers, R.O.; Ripple, W.J. Strategic reserves in Oregon’s forests for biodiversity, water, and carbon to mitigate and adapt to climate change. Front. For. Glob. Chang. 2022, 5, 1028401. [Google Scholar] [CrossRef]
- Hu, H.F.; Wang, S.P.; Guo, Z.D.; Xu, B.; Fang, J.Y. The stage⁃classified matrix models project a significant increase in biomass carbon stocks in China’s forests between 2005 and 2050. Sci. Rep. 2015, 5, 11203. [Google Scholar] [CrossRef] [PubMed]
- Le Houérou, H.N. Restoration and rehabilitation of arid and semiarid Mediterranean ecosystems in North Africa and West Asia: A review. Arid Soil Res. Rehab. 2000, 14, 3–14. [Google Scholar] [CrossRef]
- Byambadorj, S.O.; Park, B.B.; Lkhagvasuren, S.; Tsedensodnom, E.; Byambasuren, O.; Khajid, A.; Chiatante, D.; Nyam-Osor, B. Influence of irrigation on biomass partitioning in above- and belowground organs of trees planted in desert sites of Mongolia. Forests 2024, 15, 46. [Google Scholar] [CrossRef]
- Liu, J.K.; Zhang, K.B. Spatial pattern and population structure of Artemisia ordosica shrub in a desert grassland under enclosure, northwest China. Int. J. Environ. Res. Public Health 2018, 15, 946. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Y.; Lin, N.F.; Cao, B.S.; Ye, X.; Pang, B.; Du, W.; Dou, H.S.; Zou, C.X.; Xu, C.; Xu, D.L.; et al. Assessing the effectiveness of Ecological Conservation Red Line for mitigating anthropogenic habitat degradation in river corridors. Ecol. Indic. 2023, 154, 110742. [Google Scholar] [CrossRef]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.G.; Khanna, P.K. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Xu, B.; Guo, Z.D.; Piao, S.L.; Fang, J.Y. Biomass carbon stocks in China’s forests between 2000 and 2050: A prediction based on forest biomass-age relationships. Sci. China Life Sci. 2010, 53, 776–783. [Google Scholar] [CrossRef]
- Shang, W.; Li, Y.Q.; Zhao, X.Y.; Zhang, T.H.; Ma, Q.L.; Tang, J.N.; Feng, J.; Su, N. Effects of Caragana microphylla plantations on organic carbon sequestration in total and labile soil organic carbon fractions in the Horqin Sandy Land, northern China. J. Arid Land 2017, 9, 688–700. [Google Scholar] [CrossRef][Green Version]
- Gyenge, J.; Gatica, G.; Sandoval, M.; Lupi, A.M.; Gaute, M.; Fernández, M.E.; Peri, P.L. Change of soil carbon storage in monoculture tree plantations across wide environmental gradients: Argentina as a case study. For. Ecol. Manag. 2024, 552, 121565. [Google Scholar] [CrossRef]
- Zhang, K.; Su, Y.Z.; Wang, T.; Liu, T.N. Soil stoichiometry characteristics of Haloxylon ammodendron with different plantation age in the desert-oasis ecotone, north China. Acta Ecol. Sin. 2016, 36, 3235–3243. (In Chinese) [Google Scholar]
- Chai, Q.L.; Ma, Z.Y.; An, Q.Q.; Wu, G.L.; Chang, X.F.; Zheng, J.Y.; Wang, G.J. Does Caragana korshinskii plantation increase soil carbon continuously in a water-limited landscape on the Loess Plateau, China? Land Degrad. Dev. 2019, 30, 1691–1698. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).