Prediction of Potential Habitat of Monochamus alternatus Based on Shared Socioeconomic Pathway Scenarios


Abstract
:1. Introduction
2. Materials and Methods
2.1. Constructing Data to Build SDMs
2.1.1. Occurrence Points
2.1.2. Ecoclimatic Indices (EIs)
2.1.3. Terrain Variables
2.1.4. Forest Theme Map (FTM)
2.2. Species Distribution Models (SDMs)
2.2.1. MaxEnt
2.2.2. Ensemble
2.3. Validating Accuracy of SDMs
2.3.1. Validating Accuracy of MaxEnt
2.3.2. Validating Accuracy of Ensemble
3. Results
3.1. MaxEnt Prediction Results
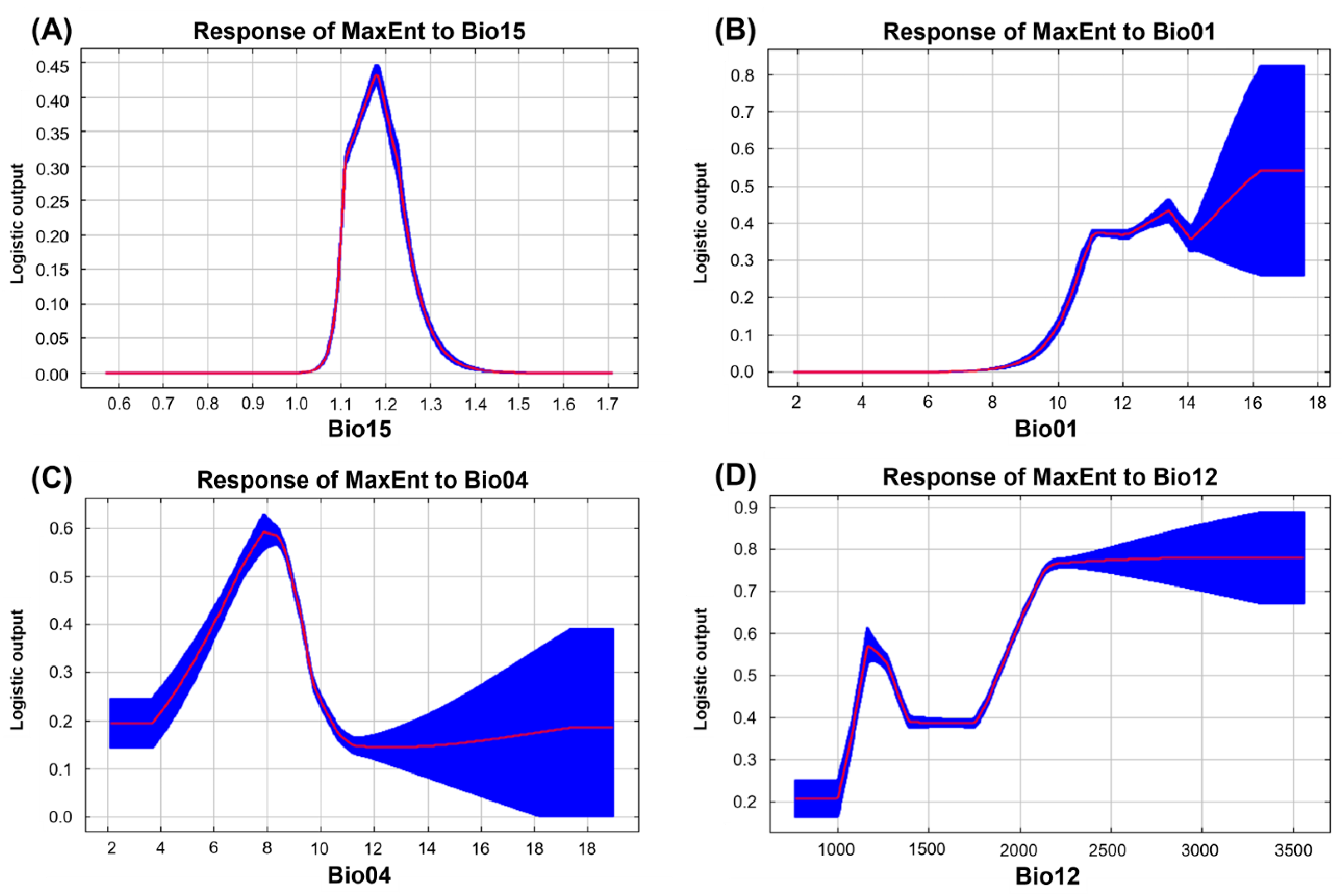
3.1.1. Evaluating Variables
3.1.2. Jackknife Validation
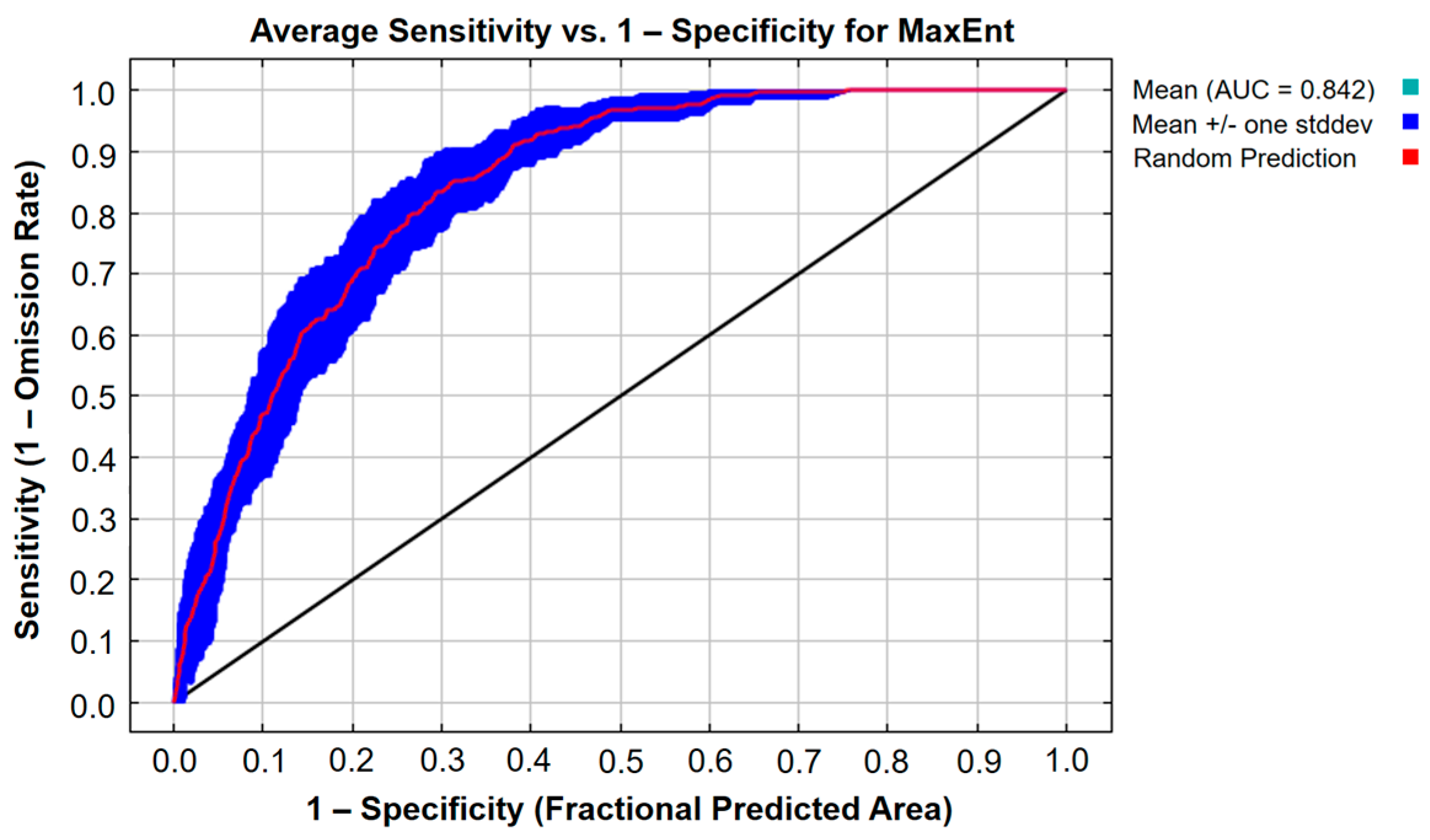
3.1.3. Evaluating Accuracy
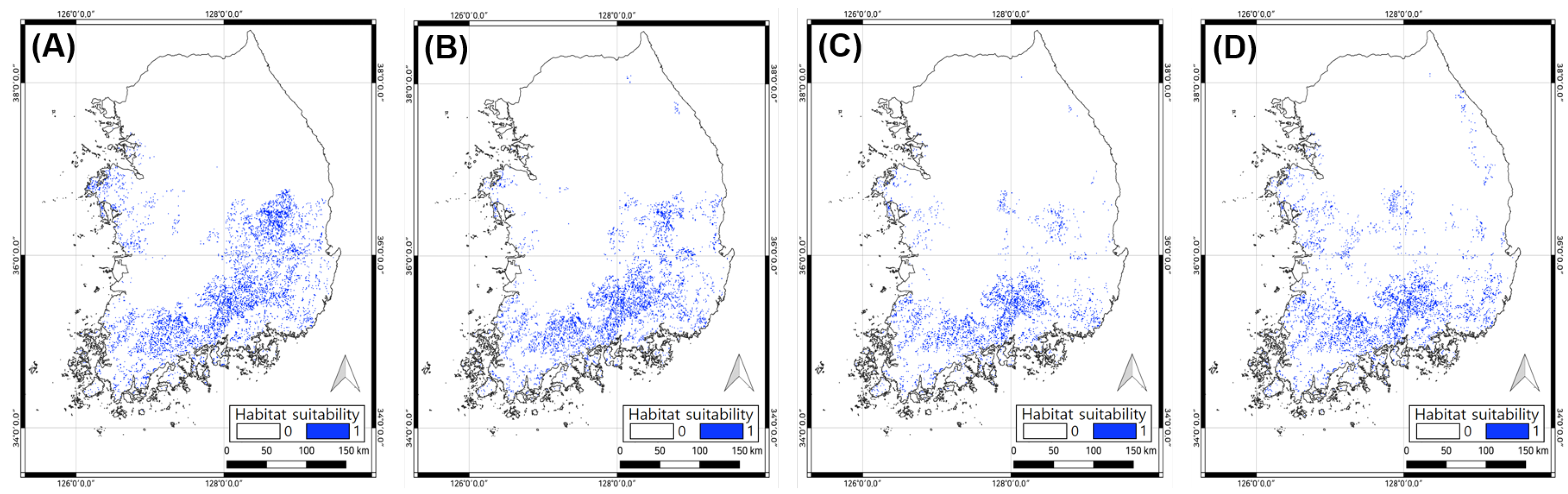
3.1.4. Potential Habitat Prediction
3.2. Ensemble Prediction Results
3.2.1. Evaluating Variables
3.2.2. Evaluating Accuracy
3.2.3. Potential Habitat Prediction
3.3. Comparing MaxEnt and Ensemble Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Separation | Description | Categorization |
|---|---|---|
| Bio01 | Average annual temperature | °C |
| Bio02 | Average diurnal range | °C |
| Bio03 | Isothermal | % |
| Bio04 | Temperature seasonality (standard deviation) | °C |
| Bio04a | Temperature Seasonality (CV) | % |
| Bio05 | Highest temperature in the warmest month | °C |
| Bio06 | Minimum temperature in the coldest month | °C |
| Bio07 | Annual temperature range | °C |
| Bio08 | Average temperature in the wettest quarter | °C |
| Bio09 | Average temperature in the driest quarter | °C |
| Bio10 | Average temperature in the warmest quarter | °C |
| Bio11 | Average temperature in the coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation in the wettest month | mm |
| Bio14 | Precipitation in the driest months | mm |
| Bio15 | Precipitation seasonality | % |
| Bio16 | Wettest quarter precipitation | mm |
| Bio17 | Dryest quarter precipitation | mm |
| Bio18 | Warmest quarter precipitation | mm |
| Bio19 | Coldest quarter precipitation | mm |
Appendix B
| Bio01 | Bio02 | Bio03 | Bio04 | Bio04a | Bio05 | Bio06 | Bio07 | Bio08 | Bio09 | Bio10 | Bio11 | Bio12 | Bio13 | Bio14 | Bio15 | Bio16 | Bio17 | Bio18 | Bio19 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.000 | −0.414 | −0.132 | −0.733 | −0.769 | 0.900 | 0.961 | −0.785 | 0.825 | 0.938 | 0.888 | 0.967 | −0.219 | −0.264 | −0.010 | −0.316 | −0.340 | 0.088 | −0.421 | 0.164 |
| 1.000 | 0.920 | 0.720 | 0.710 | −0.067 | −0.526 | 0.687 | −0.035 | −0.535 | −0.061 | −0.534 | 0.166 | 0.251 | −0.388 | 0.401 | 0.295 | −0.388 | 0.303 | −0.372 | |
| 1.000 | 0.408 | 0.393 | 0.129 | −0.207 | 0.364 | 0.129 | −0.216 | 0.124 | −0.219 | 0.116 | 0.145 | −0.384 | 0.203 | 0.179 | −0.316 | 0.158 | −0.262 | ||
| 1.000 | 0.999 | −0.377 | −0.884 | 0.995 | −0.253 | −0.902 | −0.345 | −0.880 | 0.117 | 0.291 | −0.304 | 0.626 | 0.337 | −0.451 | 0.416 | −0.523 | |||
| 1.000 | −0.426 | −0.908 | 0.998 | −0.304 | −0.922 | −0.395 | −0.904 | 0.129 | 0.296 | −0.284 | 0.613 | 0.345 | −0.429 | 0.425 | −0.502 | ||||
| 1.000 | −0.764 | −0.452 | 0.954 | 0.723 | 0.997 | 0.771 | −0.208 | −0.167 | −0.197 | −0.056 | −0.243 | −0.139 | −0.317 | −0.073 | |||||
| 1.000 | −0.921 | 0.657 | 0.990 | 0.741 | 0.999 | −0.188 | −0.292 | 0.117 | −0.472 | −0.361 | 0.248 | −0.454 | 0.329 | ||||||
| 1.000 | −0.331 | −0.933 | −0.422 | −0.915 | 0.135 | 0.303 | −0.282 | 0.618 | 0.353 | −0.427 | 0.436 | −0.500 | |||||||
| 1.000 | 0.598 | 0.966 | 0.668 | −0.286 | −0.199 | −0.252 | 0.045 | −0.261 | −0.258 | −0.276 | −0.240 | ||||||||
| 1.000 | 0.697 | 0.989 | −0.158 | −0.279 | 0.181 | −0.523 | −0.350 | 0.325 | −0.447 | 0.413 | |||||||||
| 1.000 | 0.749 | −0.208 | −0.161 | −0.200 | −0.030 | −0.234 | −0.157 | −0.299 | −0.100 | ||||||||||
| 1.000 | −0.191 | −0.290 | 0.108 | −0.456 | −0.359 | 0.234 | −0.448 | 0.315 | |||||||||||
| 1.000 | 0.901 | 0.350 | 0.256 | 0.944 | 0.406 | 0.876 | 0.451 | ||||||||||||
| 1.000 | 0.112 | 0.578 | 0.973 | 0.123 | 0.913 | 0.153 | |||||||||||||
| 1.000 | −0.553 | 0.129 | 0.951 | 0.132 | 0.861 | ||||||||||||||
| 1.000 | 0.542 | −0.646 | 0.569 | −0.645 | |||||||||||||||
| 1.000 | 0.145 | 0.964 | 0.171 | ||||||||||||||||
| 1.000 | 0.100 | 0.970 | |||||||||||||||||
| 1.000 | 0.092 | ||||||||||||||||||
| 1.000 |
| Variable | Contribution (%) | Importance (%) |
|---|---|---|
| Bio01 | 22.7 | 17.5 |
| Bio02 | 4.9 | 7.9 |
| Bio04 | 19.2 | 17.6 |
| Bio12 | 14.4 | 8.4 |
| Bio14 | 2.4 | 5.7 |
| Bio15 | 24.2 | 34.5 |
| Aspect | 3.3 | 3.4 |
| Slope | 0.9 | 0.8 |
| DEM | 4.3 | 3.3 |
| FTM | 3.8 | 0.8 |
| Variable | Importance (%) |
|---|---|
| Bio01 | 0.101 |
| Bio02 | 0.240 |
| Bio04 | 0.196 |
| Bio12 | 0.079 |
| Bio14 | 0.038 |
| Bio15 | 0.202 |
| Aspect | 0.046 |
| Slope | 0.018 |
| DEM | 0.029 |
| FTM | 0.005 |
| MaxEnt | Ensemble | |||
|---|---|---|---|---|
| Separation | Year | Area (km2) | Year | Area (km2) |
| Baseline Period | 1981 to 2010 | 4807 | 1981 to 2010 | 5477 |
| SSP2-4.5 | 2011 to 2040 | 4926 | 2011 to 2040 | 4862 |
| 2041 to 2070 | 5942 | 2041 to 2070 | 3811 | |
| 2071 to 2100 | 7600 | 2071 to 2100 | 3177 | |
| SSP5-8.5 | 2011 to 2040 | 4262 | 2011 to 2040 | 4318 |
| 2041 to 2070 | 7278 | 2041 to 2070 | 3136 | |
| 2071 to 2100 | 11,345 | 2071 to 2100 | 4034 | |
References
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Linit, M.J. Nematode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227. [Google Scholar] [PubMed]
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.I.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle, and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. [Google Scholar]
- Kang, D.I.; Son, S.C.; Yu, S.M.; Lee, B.Y.; Park, Y.S. 2022 Statistical Yearbook of Forestry. Available online: https://kfss.forest.go.kr/stat/ptl/fyb/frstyYrBookList.do?curMenu=9854 (accessed on 10 December 2022).
- Lu, Q.; Wang, W.; Liang, J. Potential suitability assessment of Bursaphelenchus xylophilus in China. For. Res. Chin. Acad. For. 2005, 18, 460. [Google Scholar]
- Lee, M.G.; Cho, H.B.; Youm, S.K.; Kim, S.W. Detection of pine wilt disease using time series UAV imagery and deep learning semantic segmentation. Forests 2023, 14, 1576. [Google Scholar] [CrossRef]
- Futai, K. Pine wilt in Japan: From first incidence to the present. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 5–12. [Google Scholar]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Matsui, T. Pine wilt disease. J. For. Res. 2008, 13, 3–11. [Google Scholar]
- Kim, J.B.; Kim, D.Y.; Park, N.C. Development of aerial precision monitoring techniques for pine wilt disease using GIS and GPS. J. Korean Soc. Geospat. Inf. Sci. 2010, 13, 28–34. [Google Scholar]
- Kim, M.J.; Bang, H.S.; Lee, J.W. Monitoring techniques for the leading edge of pine wilt disease using unmanned aerial vehicles: Focused on Sejong City. J. Korean For. Soc. 2017, 106, 100–109. [Google Scholar]
- Jung, C.S.; Jeong, Y.J.; Moon, I.S.; Han, H.R.; Ko, S.H.; Choi, K.S.; Kim, K.H.; Lim, J.H.; Yang, H.M.; Shin, H.C.; et al. Research on the Ecological Characteristics of Pine Wilt Disease; Korea Forest Research Institute: Seoul, Republic of Korea, 2009. [Google Scholar]
- Wilson, J.W.; Sexton, J.O.; Jobe, R.T.; Haddad, N.M. The relative contribution of terrain, land cover, and vegetation structure indices to species distribution models. Biol. Conserv. 2013, 164, 170–176. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Mynsberge, A.R. Evaluating ensemble forecasts of plant species distributions under climate change. Ecol. Model. 2013, 266, 126–130. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Thorn, J.S.; Nijman, V.; Smith, D.; Nekaris, K.A.I. Ecological niche modelling as a technique for assessing threats and setting conservation priorities for Asian slow lorises (Primates: Nycticebus). Divers. Distrib. 2009, 15, 289–298. [Google Scholar] [CrossRef]
- Venette, R. Pest Risk Modeling and Mapping for Invasive Alien Species; CABI: Wallingford, UK, 2015; pp. 1–17. [Google Scholar]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Berger, A.; Della Pietra, S.A.; Della Pietra, V.J. A maximum entropy approach to natural language processing. Comput. Linguist. 1996, 22, 39–71. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Kwon, H.S. Application of ensemble models for uncertainty assessment of species distribution models. J. Korean Assoc. Geogr. Inf. Stud. 2014, 22, 47–52. [Google Scholar]
- Jung, H.I.; Choi, Y.Y.; Ryu, J.E.; Jeon, S.W. Accuracy evaluation of predicted distribution of Pinus koraiensis potential habitats using species distribution models-Verification of ensemble methodology. J. Korean Soc. Clim. Change Res. 2020, 11, 37–51. [Google Scholar]
- Araújo, M.B.; Whittaker, R.J.; Ladle, R.J.; Erhard, M. Reducing uncertainty in projections of extinction risk from climate change. Glob. Ecol. Biogeogr. 2005, 14, 529–538. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD-a platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Meller, L.; Cabeza, M.; Pironon, S.; Barbet-Massin, M.; Maiorano, L.; Georges, D.; Thuiller, W. Ensemble distribution models in conservation prioritization: From consensus predictions to consensus reserve networks. Divers. Distrib. 2014, 20, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Choi, W.I.; Nam, Y.; Park, Y.S. Predicting potential occurrence of pine wilt disease based on environmental factors in South Korea using machine learning algorithms. Ecol. Inform. 2021, 64, 101378. [Google Scholar] [CrossRef]
- Choi, H.J. Predicting the Geographical Distribution of Pine Wilt Disease and Oak Wilt Disease under Climate Change Scenarios. Master’s Thesis, Gyeongsang National University, Graduate School, Jinju-si, Republic of Korea, 2019. [Google Scholar]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takenaka Takano, K.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef]
- Kim, J.W.; Jeong, H.C.; Park, Y.H. Predicting the potential distribution of Monochamus alternatus under climate change. Korean J. Appl. Entomol. 2016, 55, 501–511. [Google Scholar] [CrossRef]
- Arguez, A.; Vose, R.S. The definition of the standard WMO climate normal: The key to deriving alternative climate normals. Bull. Am. Meteorol. Soc. 2011, 92, 699–704. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Korea Forestry Promotion Institute. 2023. Available online: https://namudr.kofpi.or.kr (accessed on 12 December 2022).
- Jung, Y.Y.; Kim, S.W. A Study on the UAV-based vegetable index comparison for detection of pine wilt disease trees. J. Cadastre Land InformatiX 2020, 50, 201–214. [Google Scholar]
- Yackulic, C.B.; Chandler, R.; Zipkin, E.F.; Royle, J.A.; Nichols, J.D.; Campbell Grant, E.H.; Veran, S. Presence-only modelling using MAXENT: When can we trust the inferences? Methods Ecol. Evol. 2013, 4, 236–243. [Google Scholar] [CrossRef]
- Xu, T.; Hutchinson, M. ANUCLIM Version 6.1 User Guide; The Australian National University, Fenner School of Environment and Society: Acton, Australia, 2011. [Google Scholar]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Shim, G.M.; Kim, Y.S.; Heo, J.N.; Kang, M.G.; Jo, S.R.; Choi, W.J. A Technical Guide for Producing Detailed Climate Change Scenario Data for Agricultural Applications. Available online: https://www.korea.kr/archive/expDocView.do?docId=39829 (accessed on 13 December 2022).
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modeling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Lautenbach, S. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Negrini, M.; Fidelis, E.G.; Picanço, M.C.; Ramos, R.S. Mapping of the Steneotarsonemus spinki invasion risk in suitable areas for rice (Oryza sativa) cultivation using MaxEnt. Exp. Appl. Acarol. 2020, 80, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Jo, N.H.; Kim, E.S.; Lee, B.R.; Lim, J.H. Prediction of potential distribution of pine trees using the MaxEnt model and analysis of the relationship with environmental variables. Proc. Korean Soc. Agric. For. Meteorol. 2019, 274, 47–56. [Google Scholar]
- Kim, H.M.; Kim, C.-Y.; Cho, J.-P.; Heo, J.-N.; Song, W.K. Prediction of Acer pictum subsp. mono Distribution using Bioclimatic Predictor Based on SSP Scenario Detailed Data. Ecol. Resilient Infrastruct. 2022, 9, 163–173. [Google Scholar]
- Kiyohara, T.; Suzuki, K.; Hashimoto, H. Population dynamics of Bursaphelenchus lignicolus at the early stage after inoculation. Trans. Annu. Meet. Jpn. For. Soc. 1975, 86, 299–300. [Google Scholar]
- Son, M.H.; Lee, W.K.; Lee, S.H.; Jo, H.G.; Lee, J.H. Natural dispersion analysis of pine wilt disease damage by spatial statistical methods. J. Korean For. Soc. 2006, 95, 240–249. [Google Scholar]
- Park, W.H.; Go, D.W.; Kwon, T.S.; Nam, Y.W.; Kwon, Y.D. Analysis of Pinewood Nematode Damage Expansion in Gyeonggi Province Based on Monitoring Data from 2008 to 2015. J. Korean For. Soc. 2018, 107, 486–496. [Google Scholar]
- CGIAR-CSI.CGIAR Consortium for Spatial Information. 2023. Available online: https://srtm.csi.cgiar.org/ (accessed on 21 November 2022).
- Korea Forest Service. National Forest Information System. 2023. Available online: https://www.forest.go.kr/newkfsweb/html/HtmlPage.do?pg=/fgis/UI_KFS_5002_020100.html&mn=KFS_02_04_03_04_01&orgId=fgis (accessed on 10 November 2022).
- Jeon, H.M.; Moon, I.S.; Heo, H.S.; Lee, S.G.; Park, N.C. Breaking News of Forest Science; Korea Forest Research Institute: Seoul, Republic of Korea, 2009. [Google Scholar]
- Kim, D.S.; Kwon, T.H.; Kim, J.M.; Lee, S.G.; Park, Y.B. Oviposition of Dendroctonus valens on Pinus densiflora and Larix leptolepis infested and uninfested logs. In Proceedings of the 2016 Annual Meeting and International Symposium of the Korean Society of Applied Entomology, Jeju-si, Republic of Korea, 25–26 April 2016; p. 71. [Google Scholar]
- Korea Forest Service. 2023. Available online: https://map.forest.go.kr/forest/ (accessed on 5 November 2022).
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, L.; Zhang, J.; He, L.; Sun, R. Maxent Modeling for Predicting the Potential Geographical Distribution of Castanopsis carlesii under Various Climate Change Scenarios in China. Forests 2023, 14, 1397. [Google Scholar] [CrossRef]
- Grendár, J.M.; Grendár, M. Maximum entropy: Clearing up mysteries. Entropy 2001, 3, 58–63. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff Alberta, AB, Canada, 4–8 July 2004; p. 83. [Google Scholar]
- Chen, S.; You, C.; Zhang, Z.; Xu, Z. Predicting the Potential Distribution of Quercus oxyphylla in China under Climate Change Scenarios. Forests 2024, 15, 1033. [Google Scholar] [CrossRef]
- Townsend Peterson, A.; Papeş, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kim, N.S.; Lee, H.C.; Cha, J.Y. Study on the change of plant seasons and spatial distribution characteristics using MODIS satellite images. J. Korean Soc. Environ. Restor. Technol. 2013, 16, 59–69. [Google Scholar] [CrossRef]
- Song, W.K. Habitat analysis of Hyla suweonensis in the breeding season using species distribution modeling. J. Korean Soc. Environ. Restor. Technol. 2015, 18, 71–82. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 18, e3093. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, M.H.; Choi, S.G.; Kim, M.K.; Uh, J.W.; Yeop, S.J.; Bang, J.H.; Lee, Y.H. Predicting suitable habitats for Peromyscus maniculatus under SSPs scenarios. Ecol. Resilient Infrastruct. 2021, 8, 154–163. [Google Scholar]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Gigović, L.; Pourghasemi, H.R.; Drobnjak, S.; Bai, S. Testing a new ensemble model based on SVM and random forest in forest fire susceptibility assessment and its mapping in Serbia’s Tara National Park. Forests 2019, 10, 408. [Google Scholar] [CrossRef]
- Latif, Q.S.; Saab, V.A.; Dudley, J.G.; Hollenbeck, J.P. Ensemble modeling to predict habitat suitability for a large-scale disturbance specialist. Ecol. Evol. 2013, 3, 4348–4364. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Zimmermann, N.E. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Tasche, D. Validation of internal rating systems and PD estimates. arXiv 2006, arXiv:physics/0606071. [Google Scholar]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.W.; Park, Y.R.; Choi, Y.S. Comparison study of biological species distribution models according to types of location data. J. Korean Spat. Inf. Soc. 2008, 16, 59–64. [Google Scholar]
- Quenouille, M.H. Approximate tests of correlation in time-series 3. In Mathematical Proceedings of the Cambridge Philosophical Society; Cambridge University Press: Cambridge, UK, 1949; Volume 45, pp. 483–484. [Google Scholar] [CrossRef]
- Wu, C.F.J. Jackknife, bootstrap and other resampling methods in regression analysis. Ann. Stat. 1986, 14, 1261–1295. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, Kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modeling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Riahi, K.; Ebi, K.L.; Hallegatte, S.; Carter, T.R.; Marthur, R.; van Vuuren, D.P. A new scenario framework for climate change research: The concept of shared socioeconomic pathways. Clim. Change 2014, 122, 387–400. [Google Scholar] [CrossRef]
- Yonhap News. Chungnam Province Launches Campaign to Prevent Pine Wilt Disease in Taean. 2021. Available online: https://www.yna.co.kr/view/AKR20210307012800063 (accessed on 12 May 2024).
- Asia Economy. Forest Service starts emergency recovery of ‘Uljin Geumgang Pine Forest’ damaged by heavy snowfall. 2024. Available online: https://view.asiae.co.kr/article/2024032207542853226 (accessed on 24 May 2024).
- GlobalE. [Global E-Photo] Premium Forest <2> Gangneung Daegwallyeong Pine Forest… 100-Year-Old Geumgang Pine Colony. 2021. Available online: https://www.globale.co.kr/news/articleView.html?idxno=13813 (accessed on 24 May 2024).
- Thuiller, W.; Miguel, B.; Araújo, M.B.; Richard, G.; Pearson, R.W.; Lluis, B.; Sandra, L. Biodiversity conservation: Uncertainty in predictions of extinction risk. Nature 2004, 427, 145–148. [Google Scholar]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.B.; Lluls, M.; Colin, M.; Lera, S.; Pedro, D.; Terence, P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H. Do they? How do they? WHY do they differ? On finding reasons for differing performances of species distribution models. Ecography 2009, 32, 66–77. [Google Scholar] [CrossRef]
- Miller, J.A. Virtual species distribution models: Using simulated data to evaluate aspects of model performance. Prog. Phys. Geogr. 2014, 38, 117–128. [Google Scholar] [CrossRef]
- Ahn, Y.J.; Lee, D.G.; Kim, H.G.; Park, C.; Kim, J.Y.; Kim, J.W. Predicting the distribution of suitable habitats for future Korean pine trees considering uncertainty. Korean Environ. Res. Technol. 2015, 3, 51–64. [Google Scholar]
- Lee, S.J.; Shin, D.B.; Lee, A.R.; Song, J.S.; Lim, C.H.; Oh, S.H. Analysis of the characteristics of dead pine trees and prediction of risk areas in the Wangpicheon Ecological Landscape Conservation Area: Focusing on the comparison between ensemble models and the MaxEnt model. Proc. Korean Soc. Environ. Ecol. 2022, 2, 49. [Google Scholar]
- Korea Forestry Promotion Institute. Advanced Information on Mountain Soil in North Korea and Selection of Suitable Afforestation Species; Korea Forestry Promotion Institute: Seoul, Republic of Korea, 2016; p. 78. [Google Scholar]
- Yonhap News Agency. Full-scale South-North Forest Cooperation through Joint Pine Wilt Disease Control and Modernization of 10 Nurseries. 2018. Available online: https://www.yna.co.kr/view/AKR20181022171200063 (accessed on 25 May 2024).
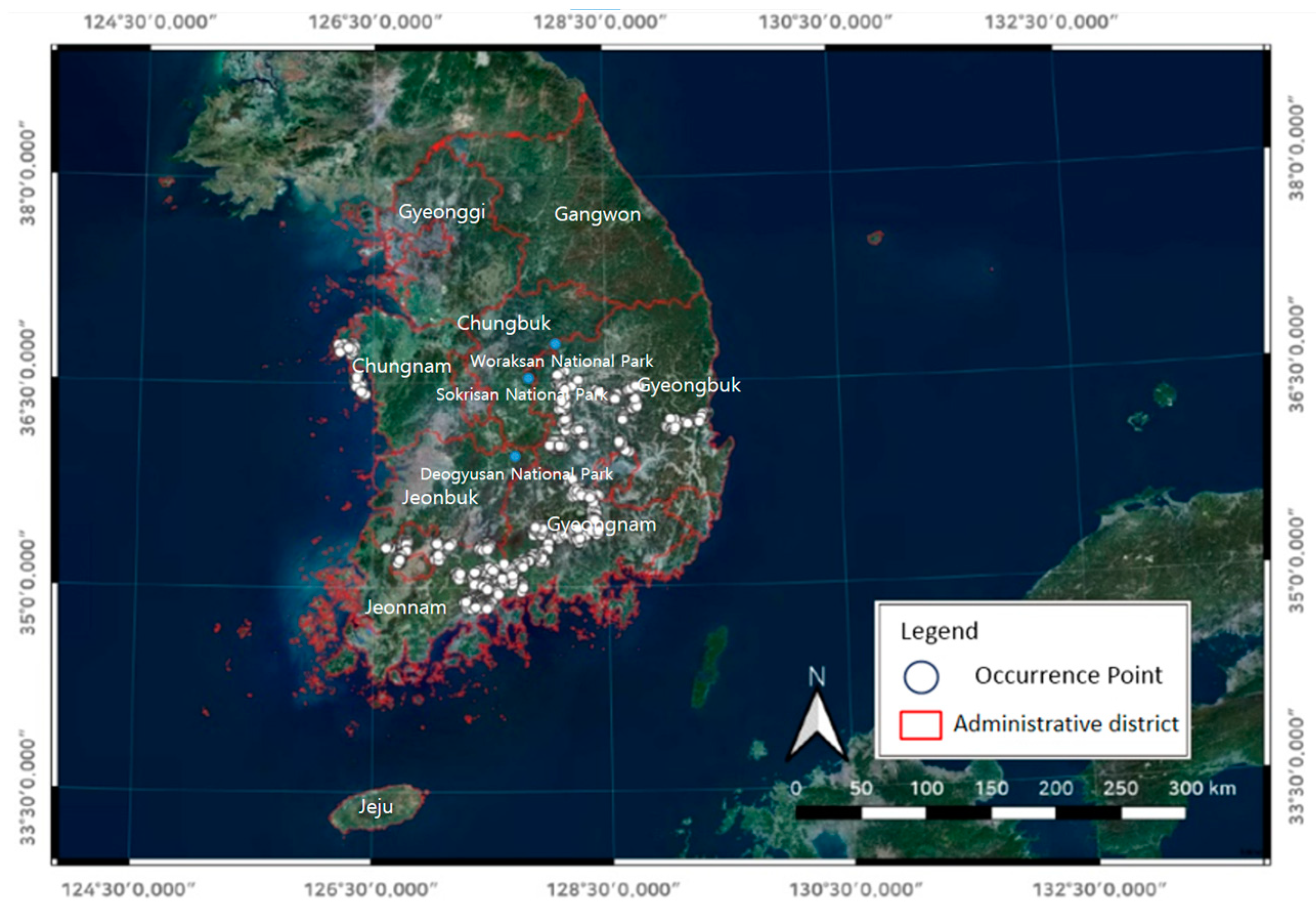







| Order | Region | Occurrence Points |
|---|---|---|
| 1 | Gyeongnam | 3902 |
| 2 | Gyeongbuk | 5555 |
| 3 | Gwangju | 221 |
| 4 | Daegu | 429 |
| 5 | Jeonnam | 9282 |
| 6 | Chungnam | 1125 |
| Total | 20,514 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, B.-J.; Lee, M.-G.; Kim, S.-W. Prediction of Potential Habitat of Monochamus alternatus Based on Shared Socioeconomic Pathway Scenarios. Forests 2024, 15, 1563. https://doi.org/10.3390/f15091563
Jung B-J, Lee M-G, Kim S-W. Prediction of Potential Habitat of Monochamus alternatus Based on Shared Socioeconomic Pathway Scenarios. Forests. 2024; 15(9):1563. https://doi.org/10.3390/f15091563
Chicago/Turabian StyleJung, Byeong-Jun, Min-Gyu Lee, and Sang-Wook Kim. 2024. "Prediction of Potential Habitat of Monochamus alternatus Based on Shared Socioeconomic Pathway Scenarios" Forests 15, no. 9: 1563. https://doi.org/10.3390/f15091563