
Quantitative Evaluation of the Applicability of Classical Forest Ecosystem Carbon Cycle Models in China: A Case Study of the Biome-BGC Model


Abstract
1. Introduction
2. Materials and Methods
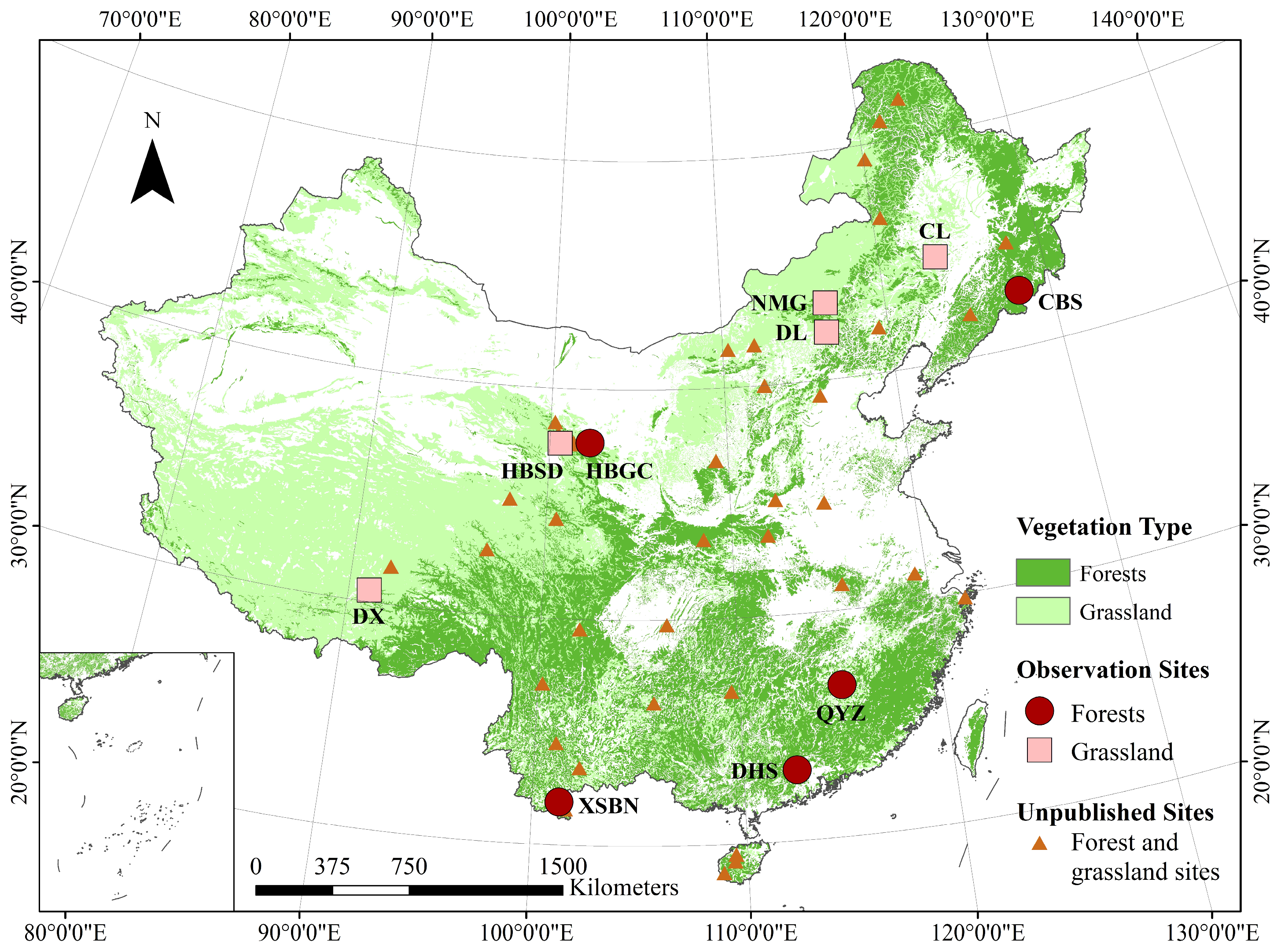
2.1. Study Area
2.2. Data Sources
2.2.1. Driving Data for the Biome-BGC Model
2.2.2. Eddy Covariance Data
2.3. Biome-BGC Model
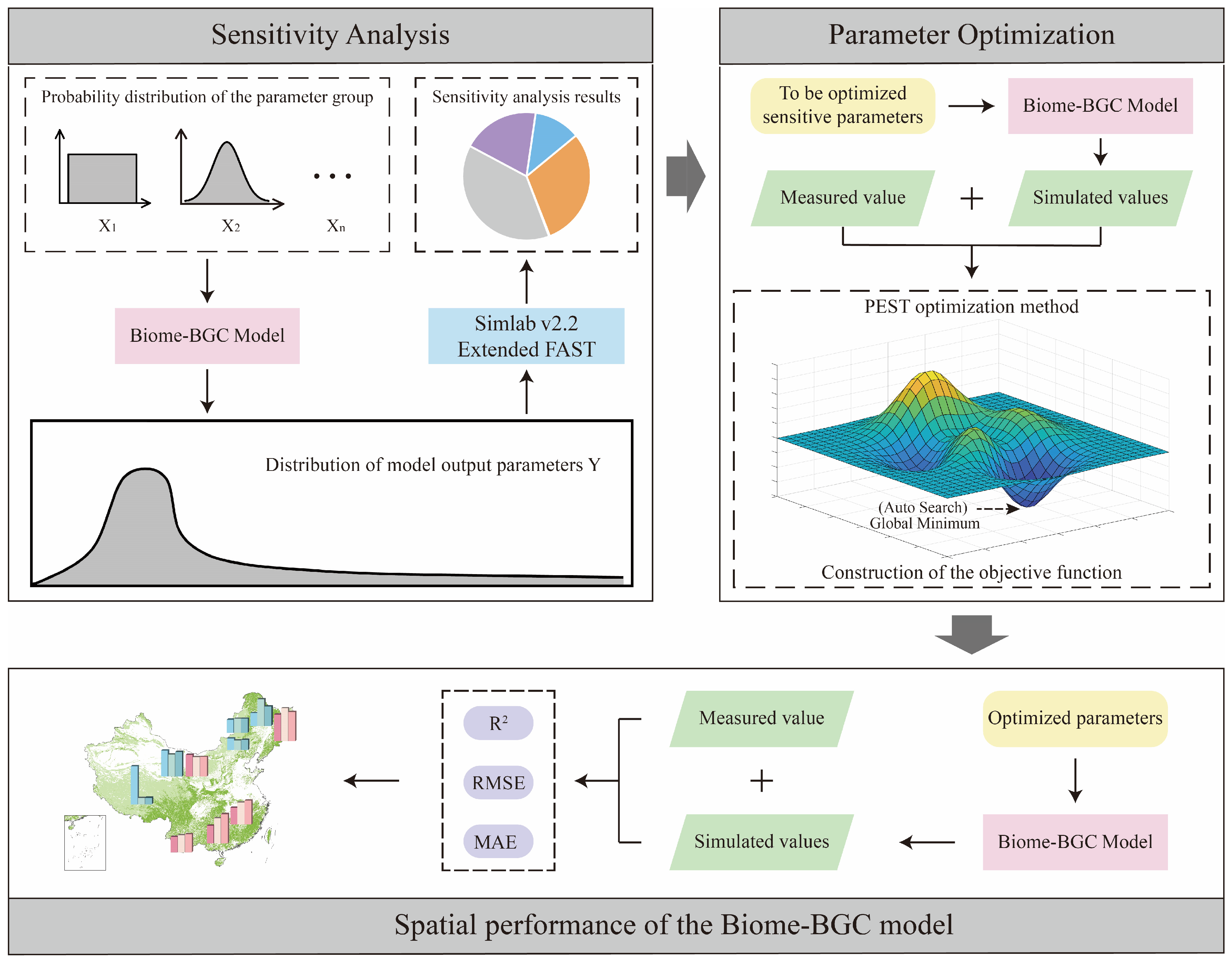
2.4. Sensitivity Analysis
2.5. Parameter Optimization
2.6. Evaluation of Simulation Accuracy
2.7. Technical Route of This Study
3. Results
3.1. Sensitivity Analysis of Ecophysiological Parameters
3.2. Calibration of Optimal Values for Sensitive Ecophysiological Parameters
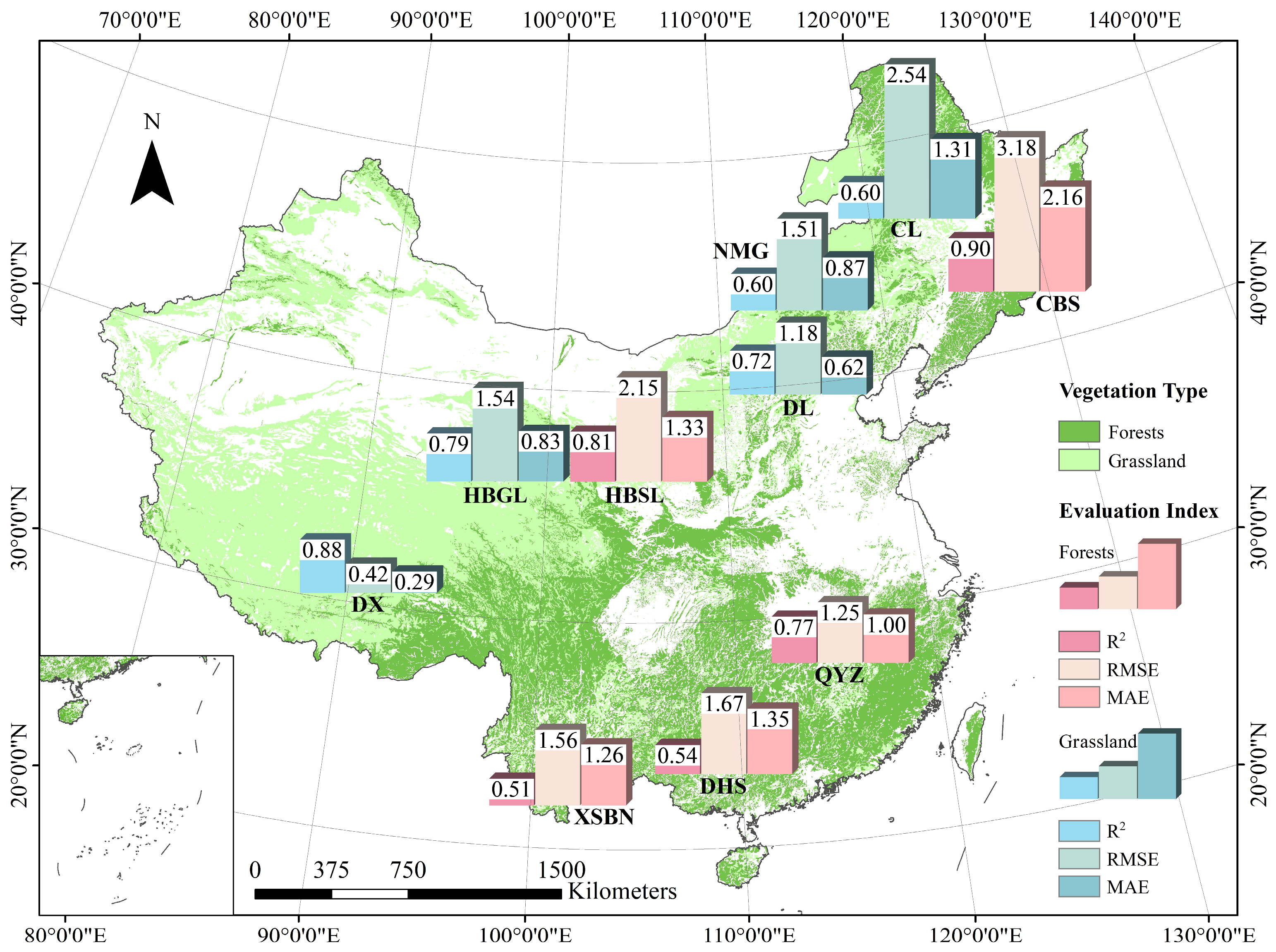
3.3. Spatial Pattern of Biome-BGC Model Applicability in China
4. Discussion
4.1. Uncertainty of Parameter Sensitivity
4.2. The Latitude Pattern of Model Applicability and Its Reasons
4.3. Suggestions for Using the Original Biome-BGC Model in China and Future Research Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, J.; Yu, G.; Liu, L.; Hu, S.; Chapin, F.S. Climate Change, Human Impacts, and Carbon Sequestration in China. Proc. Natl. Acad. Sci. USA 2018, 115, 4015–4020. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Si, G.; Yu, Q.; Huang, H.; Huang, Y.; Liu, W.; Guo, H. Study on the Relationship between Topological Characteristics of Vegetation Ecospatial Network and Carbon Sequestration Capacity in the Yellow River Basin, China. Remote Sens. 2021, 13, 4926. [Google Scholar] [CrossRef]
- Fang, J.; Ke, J.; Tang, Z.; Chen, A. Implications and estimations of four terrestrial productivity parameters. Acta Phytoecol. Sin. 2001, 25, 414–419. [Google Scholar]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of National Ecological Restoration Projects on Carbon Sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef]
- Li, X.; Ma, H.; Ran, Y.; Wang, X.; Zhu, G.; Liu, F.; He, H.; Zhang, Z.; Huang, C. Terrestrial Carbon Cycle Model-Data Fusion: Progress and Challenges. Sci. China Earth Sci. 2021, 64, 1645–1657. [Google Scholar] [CrossRef]
- Fang, M.; Cheng, C.; He, N.; Si, G.; Sun, O.J. Implication of Community-Level Ecophysiological Parameterization to Modelling Ecosystem Productivity: A Case Study across Nine Contrasting Forest Sites in Eastern China. J. For. Res. 2024, 35, 7. [Google Scholar] [CrossRef]
- Su, H.; Sang, W.; Wang, Y.; Ma, K. Simulating Picea Schrenkiana Forest Productivity under Climatic Changes and Atmospheric CO2 Increase in Tianshan Mountains, Xinjiang Autonomous Region, China. For. Ecol. Manag. 2007, 246, 273–284. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, X.; Ouyang, S.; Xu, K.; Hawkins, B.A.; Sun, O.J. A Test of BIOME-BGC with Dendrochronology for Forests along the Altitudinal Gradient of Mt. Changbai in Northeast China. J. Plant Ecol. 2017, 10, 415–425. [Google Scholar] [CrossRef]
- Luo, Z.; Sun, O.J.; Wang, E.; Ren, H.; Xu, H. Modeling Productivity in Mangrove Forests as Impacted by Effective Soil Water Availability and Its Sensitivity to Climate Change Using Biome-BGC. Ecosystems 2010, 13, 949–965. [Google Scholar] [CrossRef]
- Wen, Y.; Han, H.; Cheng, X.; Li, Z. Forest water use efficiency in Qianyanzhou based on Biome-BGC model, Jiangxi Province of eastern China. J. Beijing For. Univ. 2019, 41, 69–77. [Google Scholar] [CrossRef]
- Li, Y.Z.; Zhang, T.L.; Liu, Q.Y.; Li, Y. Temporal and spatial heterogeneity analysis of optimal value of sensitive parameters in ecological process model: The BIOME-BGC model as an example. Ying Yong Sheng Tai Xue Bao 2018, 29, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Zhang, L.; Yan, M.; Tian, X.; Zheng, X. Sensitivity Analysis of Biome-BGCMuSo for Gross and Net Primary Productivity of Typical Forests in China. For. Ecosyst. 2022, 9, 100011. [Google Scholar] [CrossRef]
- Sun, J.; Mao, F.; Du, H.; Li, X.; Xu, C.; Zheng, Z.; Teng, X.; Ye, F.; Yang, N.; Huang, Z. Improving the Simulation Accuracy of the Net Ecosystem Productivity of Subtropical Forests in China: Sensitivity Analysis and Parameter Calibration Based on the BIOME-BGC Model. Forests 2024, 15, 552. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, R.; Peng, C.; Zhou, G.; Wang, C.; Zhu, Q.; Yang, Y. Integrating a Model with Remote Sensing Observations by a Data Assimilation Approach to Improve the Model Simulation Accuracy of Carbon Flux and Evapotranspiration at Two Flux Sites. Sci. China Earth Sci. 2016, 59, 337–348. [Google Scholar] [CrossRef]
- Yan, M.; Tian, X.; Li, Z.; Chen, E.; Wang, X.; Han, Z.; Sun, H. Simulation of Forest Carbon Fluxes Using Model Incorporation and Data Assimilation. Remote Sens. 2016, 8, 567. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, T.; Du, M.; Gao, H.; Zhang, Q.; Sun, R. A Better Carbon-Water Flux Simulation in Multiple Vegetation Types by Data Assimilation. For. Ecosyst. 2022, 9, 100013. [Google Scholar] [CrossRef]
- Yan, M.; Li, Z.; Tian, X.; Zhang, L.; Zhou, Y. Improved Simulation of Carbon and Water Fluxes by Assimilating Multi-Layer Soil Temperature and Moisture into Process-Based Biogeochemical Model. For. Ecosyst. 2019, 6, 12. [Google Scholar] [CrossRef]
- Running, S.W.; Gower, S.T. FOREST-BGC, A General Model of Forest Ecosystem Processes for Regional Applications. II. Dynamic Carbon Allocation and Nitrogen Budgets. Tree Physiol. 1991, 9, 147–160. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Liu, C.; Piao, S.; Sack, L.; Xu, L.; Luo, Y.; He, J.; Han, X.; Zhou, G.; Zhou, X.; et al. Ecosystem Traits Linking Functional Traits to Macroecology. Trends Ecol. Evol. 2019, 34, 200–210. [Google Scholar] [CrossRef]
- Earth’s CO2 Home Page. Available online: https://www.co2.earth/ (accessed on 13 November 2022).
- Resource and Environmental Science Data Platform. Available online: https://www.resdc.cn/ (accessed on 1 July 2024).
- Thornton, P.E.; Running, S. User’s Guide for BIOME-BGC; Version 4.1.1; Numerical Terradynamic Simulation Group, School of Forestry, University of Montana: Missoula, MT, USA, 2000. [Google Scholar]
- University of Montana. MT-CLIM. Available online: https://www.umt.edu/numerical-terradynamic-simulation-group/project/mt-clim.php/ (accessed on 23 August 2024).
- China Meteorological Data Network. Available online: https://data.cma.cn/ (accessed on 13 November 2022).
- Zhu, J.; Zhang, F.; Li, H.; He, H.; Li, Y.; Yang, Y.; Zhang, G.; Wang, C.; Luo, F. Seasonal and Interannual Variations of CO2 Fluxes Over 10 Years in an Alpine Wetland on the Qinghai-Tibetan Plateau. JGR Biogeosci. 2020, 125, e2020JG006011. [Google Scholar] [CrossRef]
- Zhu, X.-J.; Yu, G.-R.; Chen, Z.; Zhang, W.-K.; Han, L.; Wang, Q.-F.; Chen, S.-P.; Liu, S.-M.; Wang, H.-M.; Yan, J.-H.; et al. Mapping Chinese Annual Gross Primary Productivity with Eddy Covariance Measurements and Machine Learning. Sci. Total Environ. 2023, 857, 159390. [Google Scholar] [CrossRef]
- ChinaFLUX. Available online: http://www.chinaflux.org/ (accessed on 23 August 2024).
- FLUXNET. Available online: https://fluxnet.org/ (accessed on 13 November 2022).
- Srinet, R.; Nandy, S.; Patel, N.R.; Padalia, H.; Watham, T.; Singh, S.K.; Chauhan, P. Simulation of Forest Carbon Fluxes by Integrating Remote Sensing Data into Biome-BGC Model. Ecol. Model. 2023, 475, 110185. [Google Scholar] [CrossRef]
- Ichii, K.; Hashimoto, H.; White, M.A.; Potter, C.; Hutyra, L.R.; Huete, A.R.; Myneni, R.B.; Nemani, R.R. Constraining Rooting Depths in Tropical Rainforests Using Satellite Data and Ecosystem Modeling for Accurate Simulation of Gross Primary Production Seasonality. Glob. Chang. Biol. 2007, 13, 67–77. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Zeng, W.; Wang, J.; Meng, J. Development of Crown Ratio and Height to Crown Base Models for Masson Pine in Southern China. Forests 2020, 11, 1216. [Google Scholar] [CrossRef]
- Peng, H.; Jia, Y.; Zhan, C.; Xu, W. Topographic Controls on Ecosystem Evapotranspiration and Net Primary Productivity under Climate Warming in the Taihang Mountains, China. J. Hydrol. 2020, 581, 124394. [Google Scholar] [CrossRef]
- Running, S.; Hunt, E. Generalization of a Forest Ecosystem Process Model for Other Biomes, BIOME-BCG, and an Application for Global-Scale Models. In Scaling Physiological Processes: Leaf to Globe: A Volume in Physiological Ecology; Academic Press: Cambridge, MA, USA, 1993; pp. 141–158. [Google Scholar] [CrossRef]
- Thornton, P.E. Regional Ecosystem Simulation: Combining Surface-and Satellite-Based Observations to Study Linkages between Terrestrial Energy and Mass Budgets. Ph.D. Thesis, University of Montana, Missoula, MT, USA, 1998. [Google Scholar]
- Beck, M.B. Water Quality Modeling: A Review of the Analysis of Uncertainty. Water Resour. Res. 1987, 23, 1393–1442. [Google Scholar] [CrossRef]
- Cukier, R.I.; Fortuin, C.M.; Shuler, K.E.; Petschek, A.G.; Schaibly, J.H. Study of the Sensitivity of Coupled Reaction Systems to Uncertainties in Rate Coefficients. I Theory. J. Chem. Phys. 1973, 59, 3873–3878. [Google Scholar] [CrossRef]
- Sobol’, I.M. On Sensitivity Estimation for Nonlinear Mathematical Models. Mat. Model. 1990, 2, 112–118. [Google Scholar]
- Saltelli, A.; Tarantola, S.; Chan, K.P.-S. A Quantitative Model-Independent Method for Global Sensitivity Analysis of Model Output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- Saltelli, A. Sensitivity Analysis for Importance Assessment. Risk Anal. 2002, 22, 579–590. [Google Scholar] [CrossRef]
- Zagayevskiy, Y.; Deutsch, C.V. A Methodology for Sensitivity Analysis Based on Regression: Applications to Handle Uncertainty in Natural Resources Characterization. Nat. Resour. Res. 2015, 24, 239–274. [Google Scholar] [CrossRef]
- Doherty, J. PEST Model-Independent Parameter Estimation User Manual. Watermark Numer. Comput. 2004, 3338, 3349. [Google Scholar]
- Bahremand, A.; De Smedt, F. Predictive Analysis and Simulation Uncertainty of a Distributed Hydrological Model. Water Resour. Manag. 2010, 24, 2869–2880. [Google Scholar] [CrossRef]
- Mei, X.; Li, D.; Wang, Q.; Tian, J.; Tian, Z.; Liu, D. Spatial-temporal Analysis of Forest Carbon Flux of in Xiaoxing’anling Based on Biome—BGC Model. Geomat. Spat. Inf. Technol. 2021, 44, 7–10. [Google Scholar]
- Zhang, W.; Wu, B.; Ren, Y.; Yang, G. Regionally Compatible Individual Tree Growth Model under the Combined Influence of Environment and Competition. Plants 2023, 12, 2697. [Google Scholar] [CrossRef]
- Iskra, I.; Droste, R. Application of Non-Linear Automatic Optimization Techniques for Calibration of HSPF. Water Environ. Res. 2007, 79, 647–659. [Google Scholar] [CrossRef]
- Skahill, B.E.; Baggett, J.S.; Frankenstein, S.; Downer, C.W. More Efficient PEST Compatible Model Independent Model Calibration. Environ. Model. Softw. 2009, 24, 517–529. [Google Scholar] [CrossRef]
- Goegebeur, M.; Pauwels, V.R.N. Improvement of the PEST Parameter Estimation Algorithm through Extended Kalman Filtering. J. Hydrol. 2007, 337, 436–451. [Google Scholar] [CrossRef]
- Rafique, R.; Kumar, S.; Luo, Y.; Kiely, G.; Asrar, G. An Algorithmic Calibration Approach to Identify Globally Optimal Parameters for Constraining the DayCent Model. Ecol. Model. 2015, 297, 196–200. [Google Scholar] [CrossRef]
- Nolan, B.T.; Malone, R.W.; Doherty, J.E.; Barbash, J.E.; Ma, L.; Shaner, D.L. Data Worth and Prediction Uncertainty for Pesticide Transport and Fate Models in Nebraska and Maryland, United States. Pest. Manag. Sci. 2015, 71, 972–985. [Google Scholar] [CrossRef]
- Watson, T.A.; Doherty, J.E.; Christensen, S. Parameter and Predictive Outcomes of Model Simplification. Water Resour. Res. 2013, 49, 3952–3977. [Google Scholar] [CrossRef]
- PEST. Home. Available online: https://pesthomepage.org/ (accessed on 13 November 2022).
- Hodson, T.O. Root-Mean-Square Error (RMSE) or Mean Absolute Error (MAE): When to Use Them or Not. Geosci. Model Dev. 2022, 15, 5481–5487. [Google Scholar] [CrossRef]
- Di Bucchianico, A. Coefficient of Determination (2). In Encyclopedia of Statistics in Quality and Reliability; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; ISBN 978-0-470-06157-2. [Google Scholar]
- Li, X.; Sun, J. Testing parameter sensitivities and uncertainty analysis of Biome-BGC model in simulating carbon and water fluxes in broadleaved-Korean pine forests. Chin. J. Plant Ecol. 2018, 42, 1131–1144. [Google Scholar] [CrossRef]
- Zhou, C.; Hao, Z.; He, H.; Zhou, D. Sensitivity of parameters in net primary productivity model of broadleaf-Korean pine mixed. Ying Yong Sheng Tai Xue Bao 2008, 19, 929–935. [Google Scholar]
- Houborg, R.; Cescatti, A.; Migliavacca, M. Constraining Model Simulations of GPP Using Satellite Retrieved Leaf Chlorophyll. In Proceedings of the 2012 IEEE International Geoscience and Remote Sensing Symposium, Munich, Germany, 22–27 July 2012; pp. 6455–6458. [Google Scholar]
- Raj, R.; Hamm, N.A.S.; van der Tol, C.; Stein, A. Variance-Based Sensitivity Analysis of BIOME-BGC for Gross and Net Primary Production. Ecol. Model. 2014, 292, 26–36. [Google Scholar] [CrossRef]
- He, L.; Wang, H.; Lei, X. Parameter Sensitivity of Simulating Net Primary Productivity of Larix Olgensis Forest Based on BIOME-BGC Model. J. Appl. Ecol. 2016, 27, 412–420. [Google Scholar] [CrossRef]
- You, Y.; Wang, S.; Ma, Y.; Wang, X.; Liu, W. Improved Modeling of Gross Primary Productivity of Alpine Grasslands on the Tibetan Plateau Using the Biome-BGC Model. Remote Sens. 2019, 11, 1287. [Google Scholar] [CrossRef]
- Schmid, S.; Zierl, B.; Bugmann, H. Analyzing the Carbon Dynamics of Central European Forests: Comparison of Biome-BGC Simulations with Measurements. Reg. Environ. Change 2006, 6, 167–180. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, R.; Hu, B.; Feng, L.; Zhang, R. Simulation of Water and Carbon Fluxes in Harvard Forest Area by Using Improved BiomeBGC Model. Chin. J. Ecol. 2011, 30, 2099–2106. [Google Scholar] [CrossRef]
- Cheng, K.; Hu, Z.; Li, S.; Guo, Q.; Hao, Y.; Yuan, W. Improvement of Predicting Ecosystem Productivity by Modifying Carbon–Water–Nitrogen Coupling Processes in a Temperate Grassland. J. Plant Ecol. 2021, 14, 10–21. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Latitude (°, N) | Longitude (°, E) | Elevation (m) | Plant Functional Type | Annual Average Temperature (°C) | Precipitation (mm) | Provinces |
|---|---|---|---|---|---|---|---|
| CBS | 42.40 | 128.10 | 738 | DBF | 3.6 | 713 | Jilin |
| HBSL | 37.61 | 101.32 | 3190 | SHRUB | −1.2 | 535 | Qinghai |
| QYZ | 26.74 | 115.06 | 101 | ENF | 17.9 | 1542 | Jiangxi |
| DHS | 23.17 | 112.53 | 362 | EBF | 20.9 | 1956 | Guangdong |
| XSBN | 21.93 | 101.27 | 750 | EBF | 21.8 | 1493 | Yunnan |
| CL | 44.59 | 123.51 | 141 | C3grass | 4.9 | 470 | Jilin |
| NMG | 43.33 | 116.40 | 1200 | C3grass | 0.9 | 338 | Inner Mongolia |
| DL | 42.05 | 116.28 | 1317 | C3grass | 2.0 | 318 | Inner Mongolia |
| HBGL | 37.61 | 101.32 | 3202 | C3grass | −1.1 | 490 | Qinghai |
| DX | 30.50 | 91.07 | 4332 | C3grass | 1.3 | 450 | Tibet |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, M.; Liu, W.; Zhang, J.; Ma, J.; Liang, Z.; Yu, Q. Quantitative Evaluation of the Applicability of Classical Forest Ecosystem Carbon Cycle Models in China: A Case Study of the Biome-BGC Model. Forests 2024, 15, 1609. https://doi.org/10.3390/f15091609
Fang M, Liu W, Zhang J, Ma J, Liang Z, Yu Q. Quantitative Evaluation of the Applicability of Classical Forest Ecosystem Carbon Cycle Models in China: A Case Study of the Biome-BGC Model. Forests. 2024; 15(9):1609. https://doi.org/10.3390/f15091609
Chicago/Turabian StyleFang, Minzhe, Wei Liu, Jieyu Zhang, Jun Ma, Zhisheng Liang, and Qiang Yu. 2024. "Quantitative Evaluation of the Applicability of Classical Forest Ecosystem Carbon Cycle Models in China: A Case Study of the Biome-BGC Model" Forests 15, no. 9: 1609. https://doi.org/10.3390/f15091609
APA StyleFang, M., Liu, W., Zhang, J., Ma, J., Liang, Z., & Yu, Q. (2024). Quantitative Evaluation of the Applicability of Classical Forest Ecosystem Carbon Cycle Models in China: A Case Study of the Biome-BGC Model. Forests, 15(9), 1609. https://doi.org/10.3390/f15091609