
Spatiotemporal Dynamics of Forest Vegetation in Northern China and Their Responses to Climate Change



Abstract
1. Introduction
2. Data and Methods
2.1. Study Area
2.2. Datasets
2.2.1. MODIS NDVI Dataset
2.2.2. Land Cover Dataset
2.2.3. Meteorological Data
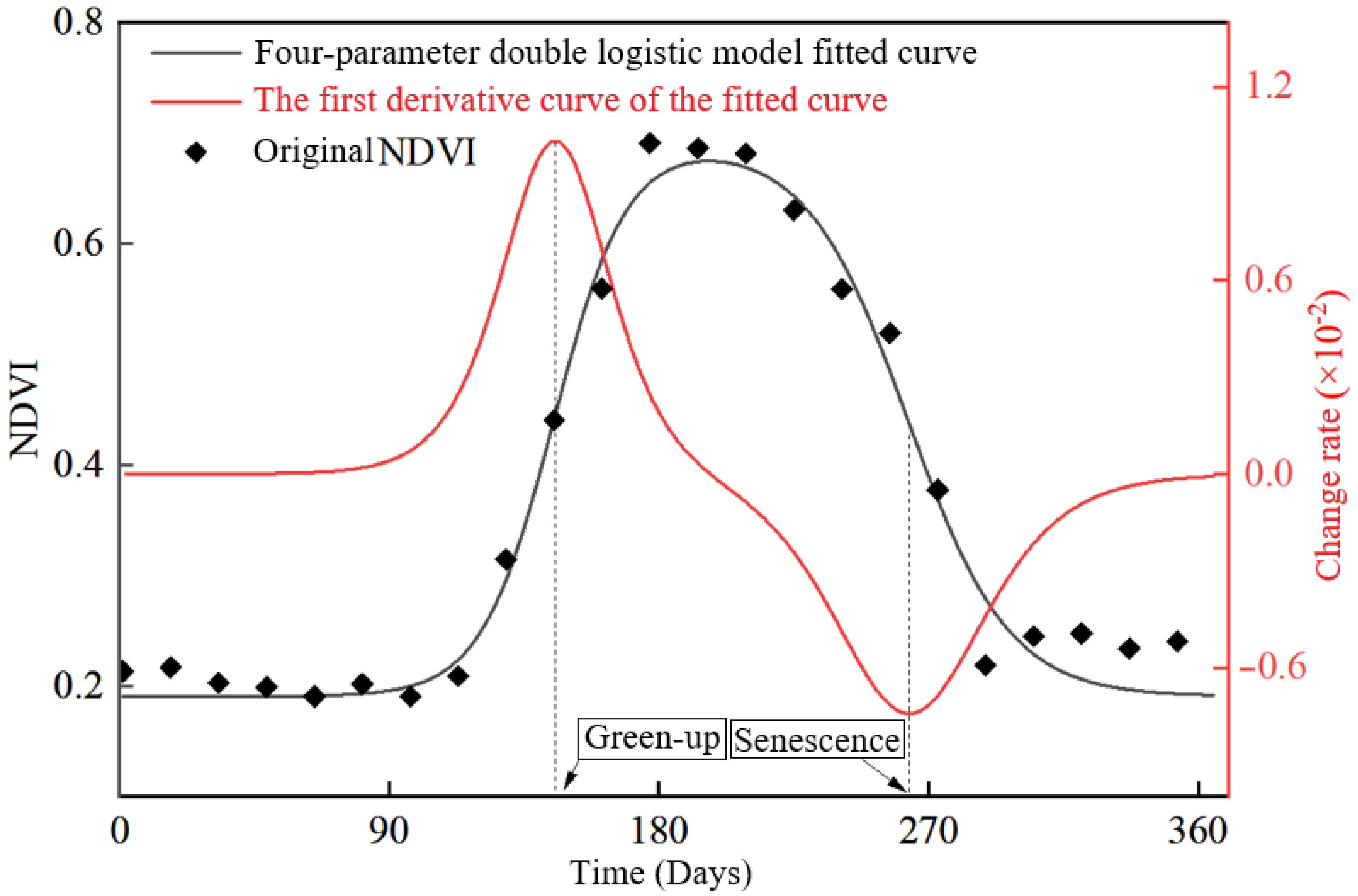
2.3. Forest Vegetation Phenological Feature Extraction
2.4. Statistical Analysis
2.4.1. Linear Regression Model
2.4.2. Partial Correlation Analysis
3. Results
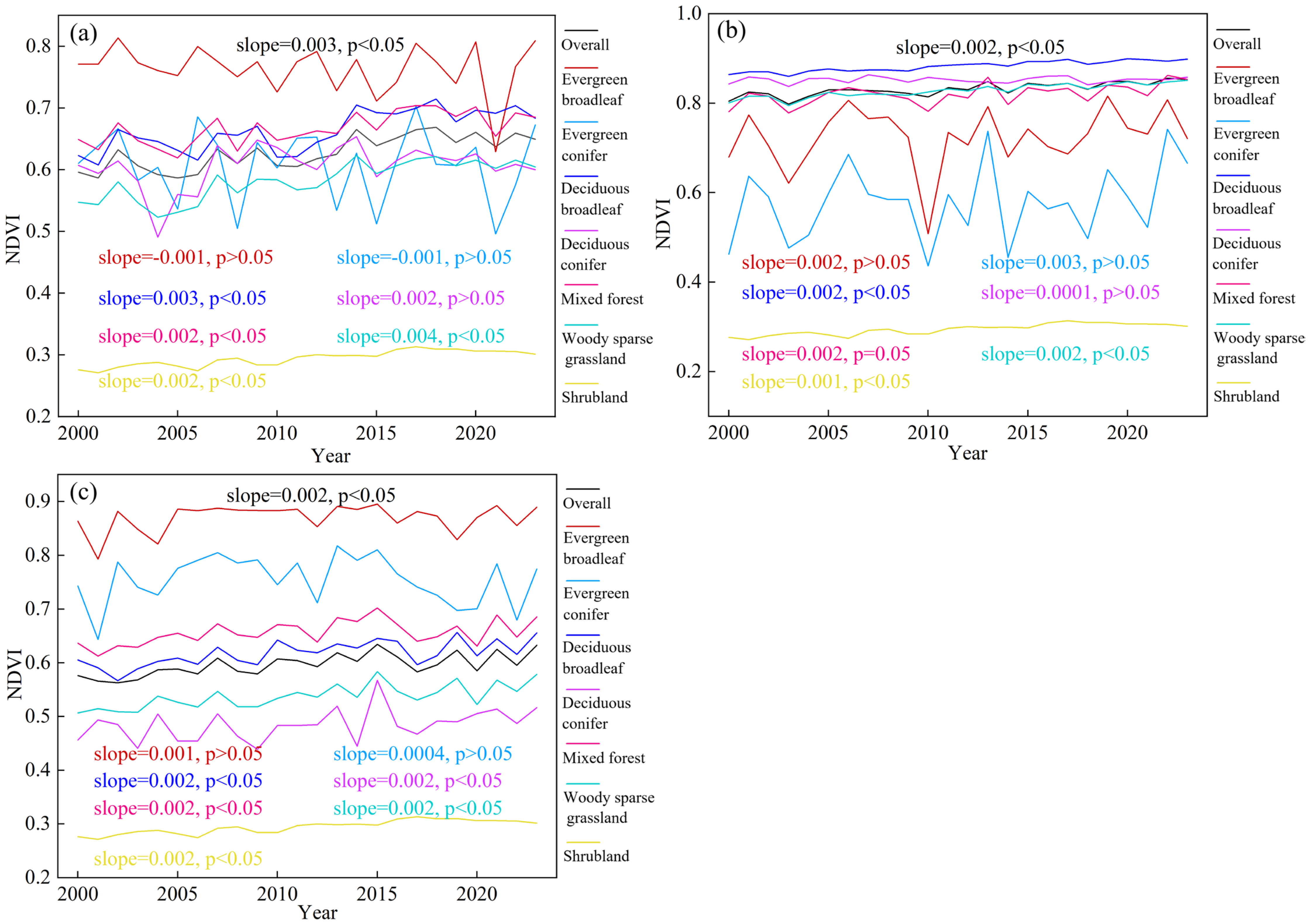
3.1. Spatiotemporal Dynamics in Forest Vegetation
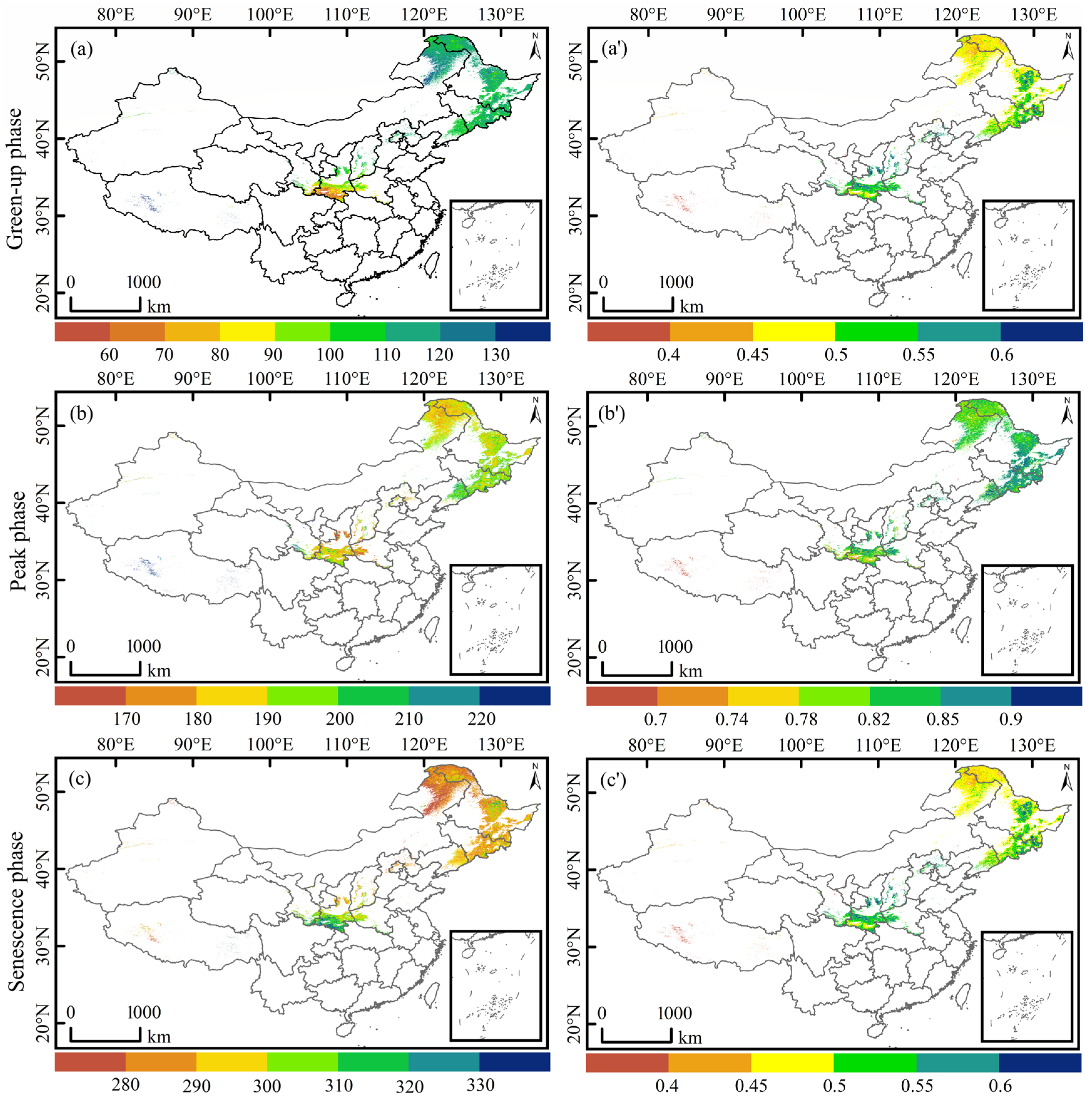
3.2. Spatiotemporal Dynamics in Forest Vegetation Phenology
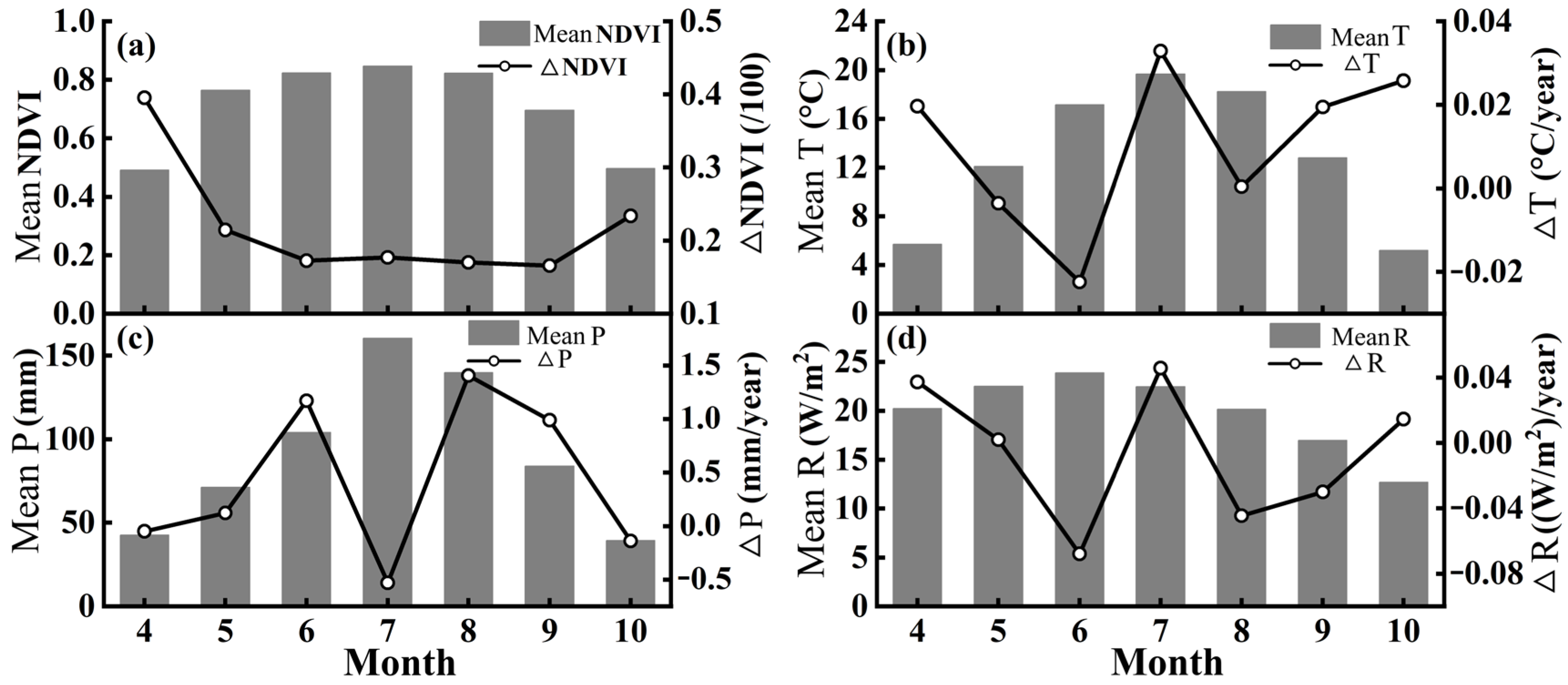
3.3. Response to Climate Change
4. Discussion
4.1. Forest Vegetation Dynamics Under Climate Change
4.2. Forest Vegetation Phenological Dynamics Under Climate Change
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhai, J.J.; Wang, L.; Liu, Y.; Wang, C.Y.; Mao, X.G. Assessing the effects of China’s Three-North Shelter Forest Program over 40 years. Sci. Total Environ. 2023, 857, 159354. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Li, S.; He, H.S.; Liang, Y.; Xu, W.R.; Wu, M.M.; Wu, Z.W.; Huang, C.; Chen, F.S. Effects of forest management practices on carbon dynamics of China’s boreal forests under changing climates. J. Environ. Manag. 2023, 335, 117497. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Trancoso, R.; Ma, Q.; Ciais, P.; Gouve, L.P.; Yue, C.F.; Assis, J.; Blanco, J.A. Carbon density in boreal forests responds non-linearly to temperature: An example from the Greater Khingan Mountains, northeast China. Agric. For. Meteorol. 2023, 338, 109519. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, Q.A.; Zhang, J.; Liu, J.; Zhu, C.F.; Qu, L.S. Vegetation dynamics alter the hydrological interconnections between upper and mid-lower reaches of the Yellow River Basin, China. Ecol. Indic. 2023, 148, 110083. [Google Scholar] [CrossRef]
- Zhao, G.L.; Tian, S.M.; Liang, S.; Jing, Y.C.; Chen, R.X.; Wang, W.W.; Han, B. Dynamic evolution trend and driving mechanisms of water conservation in the Yellow River Basin, China. Sci. Rep. 2024, 14, 26304. [Google Scholar] [CrossRef]
- Wei, X.X.; Liu, R.G.; Liu, Y.; He, J.Y.; Chen, J.L.; Qi, L.; Zhou, Y.L.; Qin, Y.W.; Wu, C.Y.; Dong, J.W.; et al. Forest Areas in China Are Recovering Since the 21st Century. Geophys. Res. Lett. 2024, 51, e2024GL110312. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Ellison, D.; Futter, M.N.; Bishop, K. On the forest cover-water yield debate: From demand- to supply-side thinking. Glob. Change Biol. 2012, 18, 806–820. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Piao, S.L.; Liu, Q.; Chen, A.P.; Janssens, I.A.; Fu, Y.S.; Dai, J.H.; Liu, L.L.; Lian, X.; Shen, M.G.; Zhu, X.L. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Visser, M.E.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. B-Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Mbow, H.-O.P.; Reisinger, A.; Canadell, J.; O’Brien, P. Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems (SR2); IPCC: Ginevra, Switzerland, 2017. [Google Scholar]
- Avitabile, V.; Camia, A. An assessment of forest biomass maps in Europe using harmonized national statistics and inventory plots. Forest Ecol. Manag. 2018, 409, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Change 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Tucker, C.J.; Townshend, J.R.G.; Goff, T.E. African Land-Cover Classification Using Satellite Data. Science 1985, 227, 369–375. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the Phenology of Global Vegetation Using Meteorological Satellite Data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Hansen, M.; Potapov, P.; Margono, B.; Stehman, S.; Turubanova, S.; Tyukavina, A. Response to comment on “High-resolution global maps of 21st-century forest cover change”. Science 2014, 344, 981. [Google Scholar] [CrossRef]
- Vermote, E.F.; El Saleous, N.Z.; Justice, C.O. Atmospheric correction of MODIS data in the visible to middle infrared: First results. Remote Sens. Environ. 2002, 83, 97–111. [Google Scholar] [CrossRef]
- Reiche, J.; Verbesselt, J.; Hoekman, D.; Herold, M. Fusing Landsat and SAR time series to detect deforestation in the tropics. Remote Sens. Environ. 2015, 156, 276–293. [Google Scholar] [CrossRef]
- Aziz, G.; Minallah, N.; Saeed, A.; Frnda, J.; Khan, W. Remote sensing based forest cover classification using machine learning. Sci. Rep. 2024, 14, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Defries, R.S.; Townshend, J.R.G. Ndvi-Derived Land-Cover Classifications at a Global-Scale. Int. J. Remote Sens. 1994, 15, 3567–3586. [Google Scholar] [CrossRef]
- Hartoyo, A.P.P.; Sunkar, A.; Ramadani, R.; Faluthi, S.; Hidayati, S. Normalized difference vegetation index (NDVI) analysis for vegetation cover in Leuser Ecosystem Area, Sumatra, Indonesia. Biodivers. J. Biol. Divers. 2021, 22, 1160–1171. [Google Scholar] [CrossRef]
- Jia, K.; Liang, S.L.; Zhang, L.; Wei, X.Q.; Yao, Y.J.; Xie, X.H. Forest cover classification using Landsat ETM plus data and time series MODIS NDVI data. Int. J. Appl. Earth Obs. 2014, 33, 32–38. [Google Scholar] [CrossRef]
- Ma, N.; Cao, S.S.; Bai, T.; Yang, Z.H.; Cai, Z.Z.; Sun, W. Assessment of Vegetation Dynamics in Xinjiang Using NDVI Data and Machine Learning Models from 2000 to 2023. Sustainability 2025, 17, 306. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Pan, Z.K.; Huang, J.F.; Zhou, Q.B.; Wang, L.M.; Cheng, Y.X.; Zhang, H.K.; Blackburn, G.A.; Yan, J.; Liu, J.H. Mapping crop phenology using NDVI time-series derived from HJ-1 A/B data. Int. J. Appl. Earth Obs. 2015, 34, 188–197. [Google Scholar] [CrossRef]
- Verhegghen, A.; Bontemps, S.; Defourny, P. A global NDVI and EVI reference data set for land-surface phenology using 13 years of daily SPOT-VEGETATION observations. Int. J. Remote Sens. 2014, 35, 2440–2471. [Google Scholar] [CrossRef]
- Wu, C.Y.; Peng, D.L.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; Lafleur, P.M.; Peichl, M.; Gonsamo, A.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Zheng, W.R.; Liu, Y.Q.; Yang, X.G.; Fan, W.Y. Spatiotemporal Variations of Forest Vegetation Phenology and Its Response to Climate Change in Northeast China. Remote Sens. 2022, 14, 2909. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- White, K.; Pontius, J.; Schaberg, P. Remote sensing of spring phenology in northeastern forests: A comparison of methods, field metrics and sources of uncertainty. Remote Sens. Environ. 2014, 148, 97–107. [Google Scholar] [CrossRef]
- Liu, R.G.; Shang, R.; Liu, Y.; Lu, X.L. Global evaluation of gap-filling approaches for seasonal NDVI with considering vegetation growth trajectory, protection of key point, noise resistance and curve stability. Remote Sens. Environ. 2017, 189, 164–179. [Google Scholar] [CrossRef]
- He, X.R.; Zhang, L.Q.; Lu, Y.H.; Chai, L.H. Spatiotemporal Variations of Vegetation and Its Response to Climate Change and Human Activities in Arid Areas—A Case Study of the Shule River Basin, Northwestern China. Forests 2024, 15, 1147. [Google Scholar] [CrossRef]
- Zhang, D.J.; Ge, W.Y.; Zhang, Y. Evaluating the vegetation restoration sustainability of ecological projects: A case study of Wuqi County in China. J. Clean. Prod. 2020, 264, 121751. [Google Scholar] [CrossRef]
- Huo, H.; Sun, C.P. Spatiotemporal variation and influencing factors of vegetation dynamics based on Geodetector: A case study of the northwestern Yunnan Plateau, China. Ecol. Indic. 2021, 130, 108005. [Google Scholar] [CrossRef]
- Alemayehu, B.; Suarez-Minguez, J.; Rosette, J. Plantation forests driven spatiotemporal vegetation trends and its interplay with climate variables in the Northwestern Highlands of Ethiopia. Int. J. Appl. Earth Obs. 2025, 137, 104411. [Google Scholar] [CrossRef]
- Estoque, R.C.; Dasgupta, R.; Winkler, K.; Avitabile, V.; Johnson, B.A.; Myint, S.W.; Gao, Y.; Ooba, M.; Murayama, Y.; Lasco, R.D. Spatiotemporal pattern of global forest change over the past 60 years and the forest transition theory. Environ. Res. Lett. 2022, 17, 084022. [Google Scholar] [CrossRef]
- Fu, W.X.; Tian, L.; Tao, Y.; Li, M.Y.; Guo, H.D. Spatiotemporal changes in the boreal forest in Siberia over the period 1985-2015 against the background of climate change. Earth Syst. Dynam. 2023, 14, 223–239. [Google Scholar] [CrossRef]
- Wahelo, T.T.; Mengistu, D.A.; Merawi, T.M. Spatiotemporal trends and drivers of forest cover change in Metekel Zone forest areas, Northwest Ethiopia. Environ. Monit. Assess. 2024, 196, 1170. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.H.; Wang, X.Z.; Li, J.W.; Yang, F.; Shangguan, Z.P.; Deng, L. The impact of vegetation reconstruction on soil erosion in the Loess plateau. J. Environ. Manag. 2024, 363, 121382. [Google Scholar] [CrossRef]
- Qiu, B.W.; Chen, G.; Tang, Z.H.; Lu, D.F.; Wang, Z.Z.; Chen, C.C. Assessing the Three-North Shelter Forest Program in China by a novel framework for characterizing vegetation changes. ISPRS J. Photogramm. 2017, 133, 75–88. [Google Scholar] [CrossRef]
- Jiang, F.G.; Deng, M.L.; Long, Y.; Sun, H. Spatial Pattern and Dynamic Change of Vegetation Greenness From 2001 to 2020 in Tibet, China. Front. Plant Sci. 2022, 13, 892625. [Google Scholar] [CrossRef]
- Li, P.L.; Zhu, D.; Wang, Y.L.; Liu, D. Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric. For. Meteorol. 2020, 295, 108190. [Google Scholar] [CrossRef]
- Dang, C.Y.; Shao, Z.F.; Huang, X.; Zhuang, Q.W.; Cheng, G.; Qian, J.X. Climate warming-induced phenology changes dominate vegetation productivity in Northern Hemisphere ecosystems. Ecol. Indic. 2023, 151, 110326. [Google Scholar] [CrossRef]
- Pulliainen, J.; Aurela, M.; Laurila, T.; Aalto, T.; Takala, M.; Salminen, M.; Kulmala, M.; Barr, A.; Heimann, M.; Lindroth, A.; et al. Early snowmelt significantly enhances boreal springtime carbon uptake. Proc. Natl. Acad. Sci. USA 2017, 114, 11081–11086. [Google Scholar] [CrossRef]
- Gu, H.S.; Qiao, Y.X.; Xi, Z.X.; Rossi, S.; Smith, N.G.; Liu, J.Q.; Chen, L. Warming-induced increase in carbon uptake is linked to earlier spring phenology in temperate and boreal forests. Nat. Commun. 2022, 13, 3698. [Google Scholar] [CrossRef]
- Zhang, T.; Shan, B.; Xu, M.; Zhao, G.; Zheng, Z.; Tang, Y.; Chen, N.; Zhu, J.; Cong, N.; Niu, B.; et al. Soil moisture alters the responses of alpine ecosystem productivity to environmental factors, especially VPD, on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2024, 947, 174518. [Google Scholar] [CrossRef]
- Jia, Q.; Gao, X.T.; Jiang, Z.L.; Li, H.X.; Guo, J.P.; Lu, X.Y.; Li, F.Y. Sensitivity of temperate vegetation to precipitation is higher in steppes than in deserts and forests. Ecol. Indic. 2024, 166, 112317. [Google Scholar] [CrossRef]
- Xiao, X.M.; Boles, S.; Liu, J.Y.; Zhuang, D.F.; Liu, M.L. Characterization of forest types in Northeastern China, using multi-temporal SPOT-4 VEGETATION sensor data. Remote Sens. Environ. 2002, 82, 335–348. [Google Scholar] [CrossRef]
- Chen, J.H.; Wang, S.Q.; Shi, H.; Chen, B.; Wang, J.B.; Zheng, C.; Zhu, K. Radiation and temperature dominate the spatiotemporal variability in resilience of subtropical evergreen forests in China. Front. Glob. Change 2023, 6, 1166481. [Google Scholar] [CrossRef]
- Griebel, A.; Bennett, L.T.; Arndt, S.K. Evergreen and ever growing—Stem and canopy growth dynamics of a temperate eucalypt forest. Forest Ecol. Manag. 2017, 389, 417–426. [Google Scholar] [CrossRef]
- Zhao, J.J.; Wang, Y.Y.; Zhang, Z.X.; Zhang, H.Y.; Guo, X.Y.; Yu, S.; Du, W.L.; Huang, F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sens. 2016, 8, 400. [Google Scholar] [CrossRef]
- Tao, J.; Zhang, Y.J.; Zhu, J.T.; Jiang, Y.B.; Zhang, X.Z.; Zhang, T.; Xi, Y. Elevation-dependent temperature change in the Qinghai-Xizang Plateau grassland during the past decade. Theor. Appl. Climatol. 2014, 117, 61–71. [Google Scholar] [CrossRef]
- Ols, C.; Kålås, I.H.; Drobyshev, I.; Söderström, L.; Hofgaard, A. Spatiotemporal variation in the relationship between boreal forest productivity proxies and climate data. Dendrochronologia 2019, 58, 125648. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types | Climate Factors | r | ||||||
|---|---|---|---|---|---|---|---|---|
| April | May | June | July | August | September | October | ||
| Evergreen broadleaf | T | n.s | n.s | n.s | n.s | n.s | n.s | 0.47 * |
| P | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| R | n.s | n.s | n.s | n.s | n.s | 0.48 * | 0.43 * | |
| Evergreen needleleaf | T | n.s | n.s | n.s | n.s | n.s | n.s | n.s |
| P | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| R | n.s | n.s | n.s | n.s | n.s | 0.60 ** | n.s | |
| Mixed forest | T | 0.50 * | n.s | n.s | n.s | n.s | n.s | n.s |
| P | n.s | n.s | n.s | n.s | n.s | 0.45 * | n.s | |
| R | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| Shrubland | T | 0.67 ** | n.s | n.s | n.s | 0.43 * | 0.57 ** | n.s |
| P | n.s | n.s | n.s | 0.44 * | n.s | n.s | n.s | |
| R | n.s | n.s | n.s | −0.53 * | n.s | n.s | n.s | |
| Deciduous broadleaf | T | 0.75 ** | n.s | n.s | 0.55 ** | n.s | n.s | n.s |
| P | 0.49 * | n.s | n.s | n.s | n.s | n.s | n.s | |
| R | 0.49 * | n.s | n.s | n.s | n.s | n.s | n.s | |
| Deciduous needleleaf | T | n.s | 0.58 ** | n.s | 0.44 * | 0.44 * | n.s | 0.44 * |
| P | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| R | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| Woody sparse grassland | T | 0.53 * | n.s | n.s | 0.50 * | n.s | 0.39 * | n.s |
| P | 0.62 ** | n.s | n.s | n.s | n.s | n.s | n.s | |
| R | 0.66 ** | n.s | n.s | 0.45 * | n.s | n.s | n.s | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, E.; Feng, Z.; Chen, P.; Wang, L. Spatiotemporal Dynamics of Forest Vegetation in Northern China and Their Responses to Climate Change. Forests 2025, 16, 671. https://doi.org/10.3390/f16040671
Ma E, Feng Z, Chen P, Wang L. Spatiotemporal Dynamics of Forest Vegetation in Northern China and Their Responses to Climate Change. Forests. 2025; 16(4):671. https://doi.org/10.3390/f16040671
Chicago/Turabian StyleMa, Erlun, Zhongke Feng, Panpan Chen, and Liang Wang. 2025. "Spatiotemporal Dynamics of Forest Vegetation in Northern China and Their Responses to Climate Change" Forests 16, no. 4: 671. https://doi.org/10.3390/f16040671
APA StyleMa, E., Feng, Z., Chen, P., & Wang, L. (2025). Spatiotemporal Dynamics of Forest Vegetation in Northern China and Their Responses to Climate Change. Forests, 16(4), 671. https://doi.org/10.3390/f16040671