1. Introduction
Trembling aspen (
Populus tremuloides Michx.) is one of the few North American tree species that has a transcontinental distribution and is found over a wide range of site conditions [
1]. In boreal mixedwood stands, aspen usually establishes immediately after a severe fire event. Over time, successional changes shift stand composition from aspen dominated to mixed species and, finally, to stands dominated by conifers [
2,
3,
4,
5,
6]. Different trajectories of canopy species replacement are possible during stand development [
7,
8], depending on the on-site climatic and edaphic conditions, the disturbance regime, regeneration potential, biotic interactions and the life histories of the species that are present [
3,
9].
Boreal forest ecosystems are largely controlled by natural disturbance events, especially fire [
10]. The fire regime, which is influenced by regional climate [
11], can significantly affect the processes of species regeneration [
6] and the long-term development of forest stands [
12]. Forest regeneration that is present in the understory can indicate the potential successional pathway that forest stand composition could take during its evolution [
13]. In the case of aspen, its composition could be maintained as pure- and hardwood-dominated stands or a transition may occur that leads to conifer-dominated stands, depending on the local environmental conditions (climate, disturbance regime and soil). Studies indicate that large and severe fires play an important role in creating stands, with little regeneration attributable to late-successional species [
14,
15]. Indeed, the recurrence of large and severe fires can regularly restart forest succession, favoring the establishment of shade-intolerant species, thereby limiting the establishment and development of late-successional species. These latter species require the presence of residual seed trees preserved in unburnt areas in order to successfully reinvade a burn site [
3,
5,
14,
16,
17,
18]; they also generally take longer to recolonize a site following fire [
19].
In the boreal forest, the nature and distribution of tree regeneration that is present at a given location is the result of climatic and physiographic conditions [
20] and the fire regime associated with this region [
21]. Ongoing climate change is expected to lead to changes in the frequency and severity of fires [
22,
23,
24], both at local and regional scales. This could change the current nature of the successional dynamics of boreal forest ecosystems [
25] and the relative importance of different temporal patterns across regions [
26]. Knowledge of the regeneration dynamics of species along a wide gradient of environmental conditions would be essential for a better understanding of vegetation response in a constantly changing environment. Furthermore, an analysis of the distribution of vegetation that is dominated by the same canopy species at different locations across a wide and continuous forest ecosystem could help resolve the potential contrasts in the understory composition between regions, by identifying the different factors that control the variation that is observed within a region [
27].
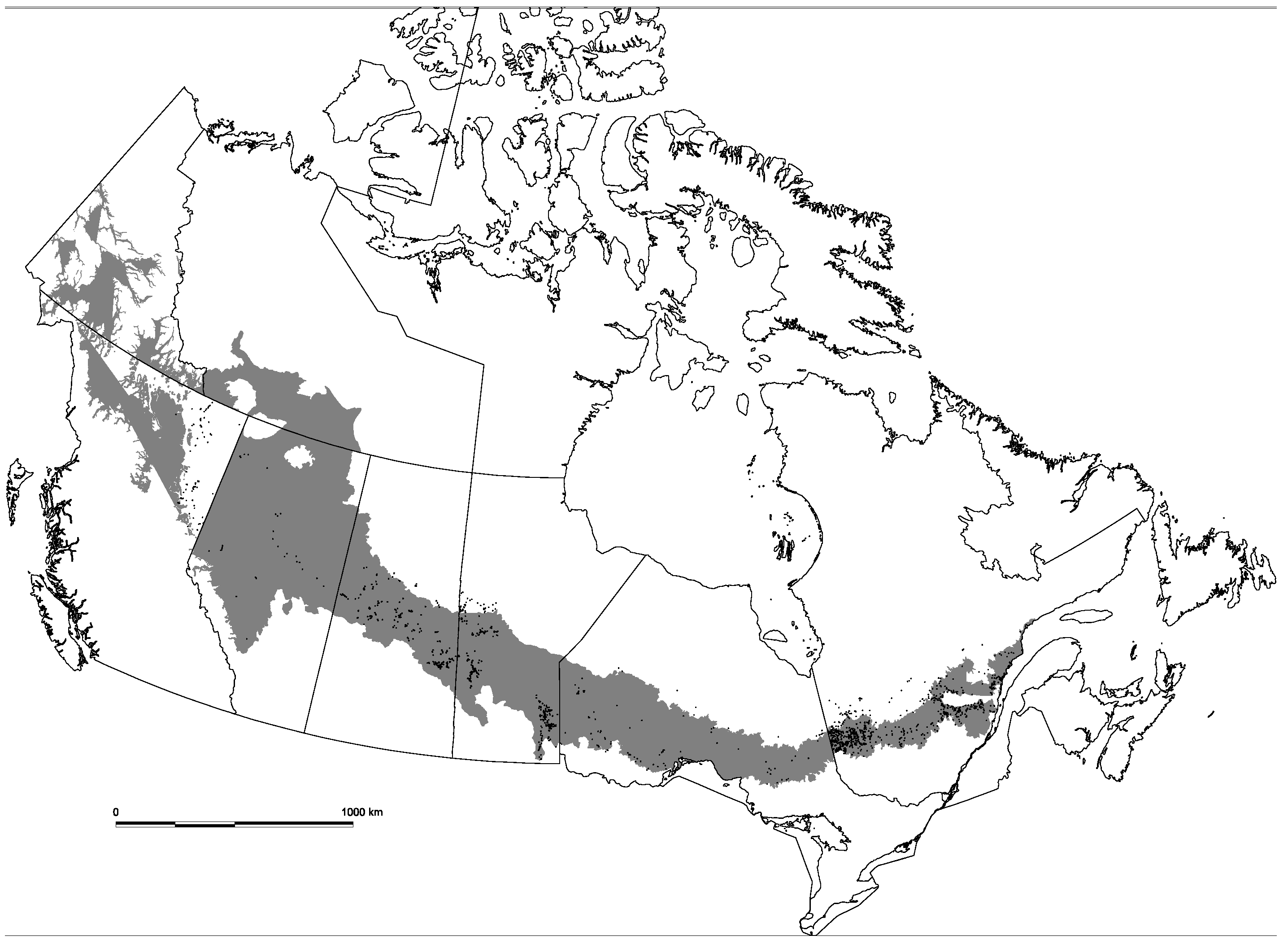
This study analyses the abundance and occurrence of recruitment of three shade-tolerant conifer species in aspen-dominated stands that are distributed along a broad gradient of climate and fire regimes across Canada. The pan-Canadian scale of this study represents a unique situation in North America. Various studies on the analysis of forest regeneration in the boreal forest are generally restricted to local or regional scales (e.g., [
28,
29,
30,
31,
32]).
The abundance and occurrence of coniferous regeneration should reflect the influence of environmental factors on the distribution of coniferous regeneration within aspen stands. Three conifer species are examined: white spruce or Ws (
Picea glauca (Moench) Voss); black spruce or Bs (
Picea mariana (Mill.) BSP); and balsam fir or Bf (
Abies balsamea (L.) Mill.). These late-successional species are commonly found in association with aspen in boreal mixedwood stands [
6,
14,
20,
33]. We only target aspen stands along the study gradient, thus controlling for the effect of forest cover. This approach allows us to observe how variability in environmental conditions affected the recruitment of shade-tolerant conifers in this stand type.
Longitudinally from east to west, the precipitation regime exhibits the most prominent change across the North American boreal forest, subsequently affecting fire regime [
33]. Large regional variation in climatic factors and the risk of fire exist among and within the Canadian provinces [
10,
34]. The contrast in precipitation and temperature and their effects on the fire regime within eastern and western Canada are likely to have an effect on the dynamics of conifer regeneration between these two regions.
Depending upon these regional differences, we hypothesize that coniferous regeneration will be more abundant in aspen stands located in the eastern boreal forest due to higher precipitation and, by extension, the longer fire cycle that dominates this part of the transect; conversely, aspen stands in the west, which experience lower precipitation and more frequent and large fires, should exhibit lower coniferous regeneration [
35,
36].
4. Discussion
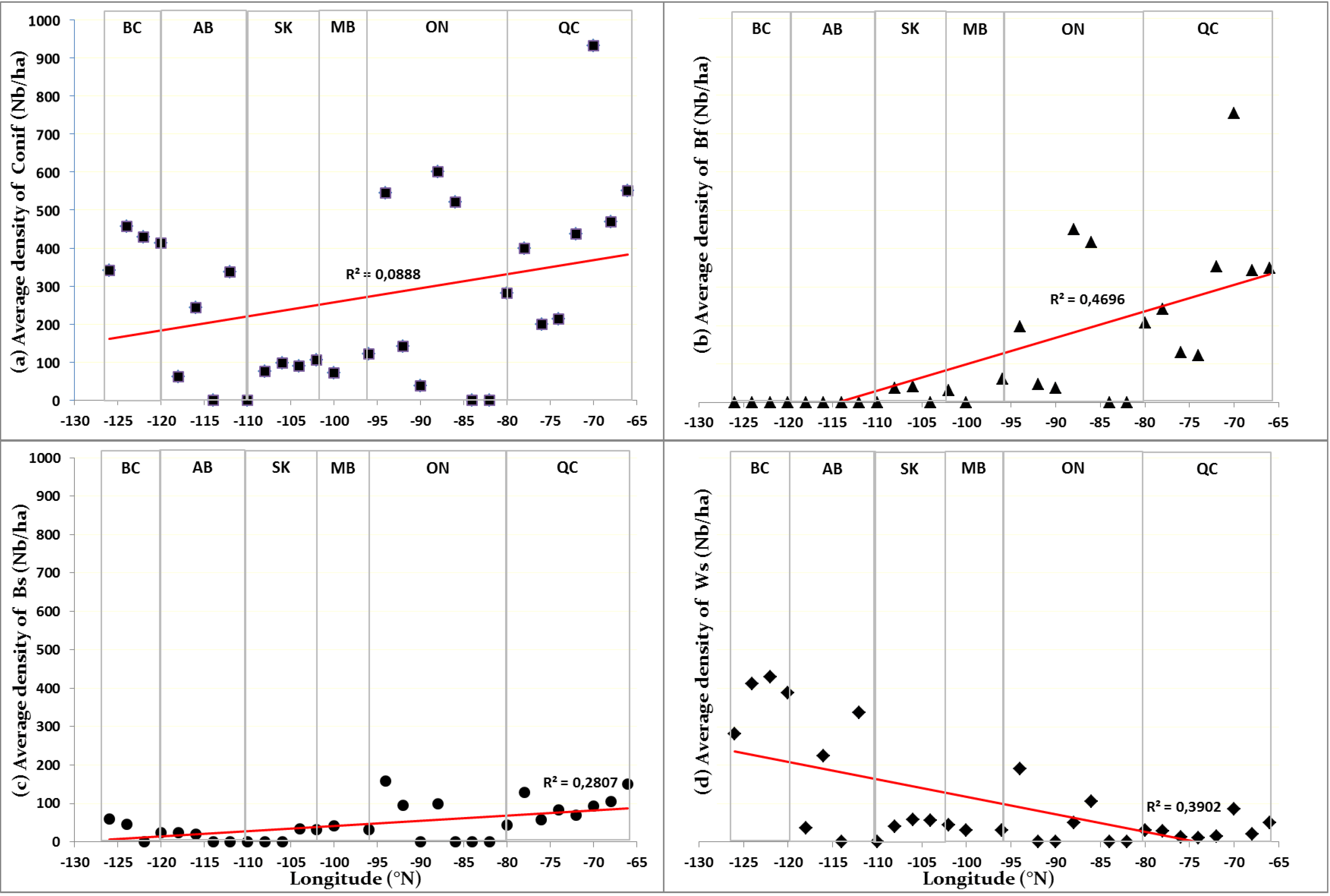
Overall, there were no significant differences in the abundance of conifer recruitment (
Figure 2a) and the proportion of aspen forests having coniferous recruitment (
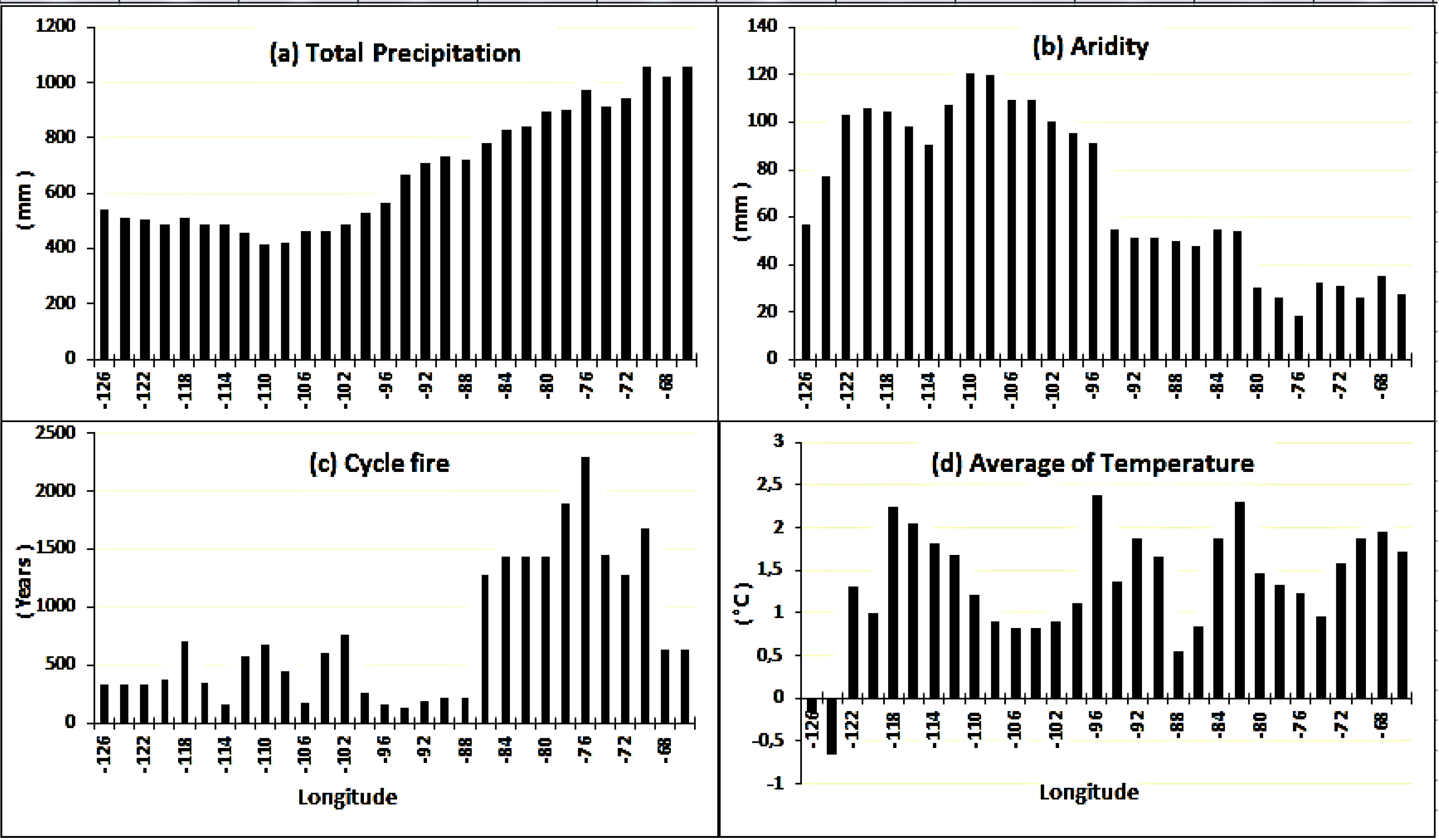
Figure 3) between the eastern and western extremes of the gradient, despite having contrasting fire regimes and climate. The higher total annual precipitation (
Appendix – Figure A1a) and the longer fire cycle (
Appendix – Figure A1c) in the east should, in principle, promote the aging of forests and an increase in shade-tolerant conifers [
51]. Yet, only about a third of aspen stands examined (32%) in QC contained conifer regeneration. This low proportion could be attributed to the intensification of forest management activities that have been undertaken in recent decades. Indeed, logging activity has increased in the southern boreal forest to the extent that it has become a major stand-replacing disturbance [
6]. In contrast, conifer recruitment was particularly low in the central part of the gradient (corresponding to MB and SK provinces) probably due to low annual precipitation (
Appendix – Figure A1a) and the aridity being higher (
Appendix – Figure A1b) in the region. Indeed, this portion of the gradient is usually characterized by a dry continental climate and a relatively short fire cycle (
Appendix – Figure A1c). Conifer recruitment is thus affected both by severe drought conditions that reduce the growth and survival of conifer seedlings and by the recurrence of severe fires, which leads to frequent rejuvenation of forest stands and promotes the establishment of pioneer species at the expense of late-successional species [
26]. Fire affects both propagule availability and substrate suitability for regeneration, which, in turn, impact regeneration and subsequent stand dynamics [
15,
18]. Mostly in the north, fire severity strongly affects the availability of regeneration microsites and, thus, has an important controlling influence on the density of initial conifer regeneration [
15,
26]. Whereas in the south, the pervasive presence of bluejoint grass,
Calamagrostis canadensis (Michx.), in the aspen understory could be an additional limiting factor for coniferous regeneration, because of its adverse effects (competition) on the growth and survival of conifer seedlings [
52].
The differences in conifer recruitment across Canada were most significant in terms of individual species. Among the major factors that guarantee the presence of a species in a forest landscape are favorable climatic conditions for its reproduction and growth, seed availability, the presence of suitable sites for its establishment and resilience to disturbances [
53].
The abundance of Bf regeneration in the eastern portion of the gradient would be associated with a usually long return interval between fires and the maritime climate that characterizes this part of the Canadian boreal forest [
11]. Conversely in aspen forests of western Canada, the recruitment of Bf would be limited because of the dry climate for germination and the fire size for seed dispersal. This species is more sensitive to drought than Ws [
54] and possesses large seeds compared to other conifer species, thereby limiting its potential for long-distance seed dispersal [
19]. Bf regenerates primarily by seed; its reproduction is severely limited by fire, because the presence of residual seed trees is required for site recolonization after disturbance [
4,
19]. In regions that are affected by large fires in a cyclical manner (the case of the western Canada boreal forest), it is possible that Bf was gradually eliminated from the landscape over time (subsisting only in some small sectors naturally protected against fires), since this species is not fire-adapted. However, its native range extends towards western Canada in scattered stands through north-central Manitoba and Saskatchewan to the Peace River Valley in northwestern Alberta, then south for approximately 640 km (400 mi) to central Alberta and east and south to southern Manitoba [
55].
Bs regeneration is still present in western aspen forests despite recurrent and severe fires in this portion of the gradient. This presence is facilitated by its adaptation to fire. It is generally able to re-establish itself quickly and amply after a fire event, as it has semi-serotinous cones that open under the effect of heat [
29,
51,
56]. It is a transcontinental tree species across Canada [
55].
The dominance of Ws regeneration in the aspen forests of western Canada seems contrary to the autecology of this coniferous species, given the predominance of short fire cycles (
Appendix – Figure A1c) and less favorable climatic conditions (low precipitation annual and drier) in this portion of the gradient. First, the abundance of Ws regeneration in western aspen stands could be explained by the greater availability of seeds of this species in this portion of the gradient. Indeed, Ws is ubiquitous in the forest landscapes of western Canada [
57]. It is likely that the existing spatial patterns of the Ws, Bs and Bf regeneration abundance are a legacy of the regions’ post-glacial history of vegetation. The overall distribution of conifer species is likely a consequence (biological legacy) of vegetation responses to environmental changes during the late Quaternary [
58]. Interactive maps of plant species distributions during the Holocene have been constructed from pollen diagrams, which reinforce this biogeographic hypothesis [
59]. These maps indicate that during deglaciation, Ws began recolonizing the North American continent from the west, while Bf started in the east and Bs moved southward from refugia in the north. These species became adapted to their environmental conditions of the early periods of migration. The distribution of coniferous regeneration that is observed in aspen stands along the longitudinal gradient, therefore, could not be only interpreted as a response to pressure exercised by climate and disturbance regimes, but should actually reflect migration routes by which these species invaded the North American continent during the post-glacial period.
Ws regeneration is everywhere along the longitudinal gradient. The longevity, low mortality rate, periodic dispersion of large seed crops (
i.e., masting) and shade-tolerance may explain the persistence of white spruce as a major species of the boreal forest [
60]. White spruce regeneration, however, is very variable. Particularly in the west, this species seems to enjoy the absence of Bf and the low presence of Bs to increase in abundance. However, Ws is less dominant in eastern Canada, probably due to competition with Bf. Moreover, Ws is less shade-tolerant than Bf [
61].
Regarding the modeling of conifer recruitment across Canada, climate and anthropogenic proxies appear clearly as the determining factors. Despite its ecological impact on the regeneration of late-successional species [
62], the fire regime (variable FC) did not appear in almost all of the selected predictive models. This is probably due to the fact that FC is very much correlated with DI (
r = 0.68). Across Canada, the risk of forest fires is closely associated with drought severity and fire activity [
63,
64,
65,
66]. Therefore, the inclusion of DI in models masks the effect of fire, thereby rendering the variable FC non-significant. Temperature and soil moisture directly influence the success of conifer regeneration. Unfavorable weather conditions (e.g., drought) would compromise the processes of germination and establishment of conifer species [
36,
54,
58]. Hydric stress or excessive temperatures during germination and seedling development can adversely affect their survival and incur significant mortality that is likely to limit or inhibit regeneration. For example, balsam fir seedlings in open spaces can undergo a high mortality rate when surface temperatures exceed 46 °C or when there is a drought or frost [
55].
Furthermore, the variable “road density” (DensRoad25Km) mainly (or “distance to agricultural area” (DistAgri)) appears in almost all of the best models, with a negative effect on the abundance and occurrence of conifer recruitments. Increasing this variable suggests an increasing pressure from human activities and their impact on forest ecosystems. The consequences of this anthropization on forest characteristics are reflected in particular by increasing pressure timber harvesting due to population growth and fire occurrence following the introduction of new sources of ignition (risks of human-caused fires). These human actions lead to significant changes in the disturbance regime, with impacts ranging from local to global scales. Because tree species respond in different ways to changes in disturbance regime, alteration in fire regimes will affect forest composition. The cumulative effect of fires and forest harvesting usually favor early-successional species at the expense of late-successional tolerant conifers species. The recurrence of severe fires locally affects the seed bank of shade-tolerant species, which show a low dispersion capacity of seeds and a slow growth rate; unlike the shade-intolerant deciduous, which have traits better adapted to the new frequently disturbed environments [
67]. Disturbances (fire and cut) occurring quite closely over time can therefore create local extinguishing pockets of shade-tolerant species, by accentuating the constraint of dispersion. The decrease in coniferous species could be explained partly by the removal in large numbers of seed trees as a result of cuts [
68]. However, in a study aimed at describing and understanding the contribution of human activities to the transformations of forest landscapes since the early nineteenth century in the Bas-Saint-Laurent in eastern Quebec [
69], researchers observed a dramatic increase in the absolute and relative frequency of deciduous species. This increase was accompanied by a decrease in coniferous trees that dominated pre-industrial forests, such as spruce (
Picea spp.) and cedar (
Thuja occidentalis L); only balsam fir seems more resistant. Human activities were a major factor in this dominance of deciduous species.
In addition to logging, forest clearing for farmland in the north-east of North America was accompanied by bush fires, which escaped in the surrounding forests. The study findings showed that 90% of the fire surface is less than 2 km from colonization zones. These results imply a strong connectivity between fires and colonization compared to a random distribution of fires, an indication of the importance of anthropogenic fires in landscapes. The current location of aspen stands seems to match the fire occurrence of colonization in this region.



{kind=link}
{kind=link}
{kind=link}
{kind=link}


 ), white spruce (
), white spruce (  ), black spruce (
), black spruce (  ) and balsam fir (
) and balsam fir (  ) respectively.
) respectively.