3.1. Results
At age 19 there were only minor differences in race means for standing-tree AWV across the Meunna, Southport and Florentine sites (3.43–3.75 km s
−1;
Table 4). Race means at Tarraleah, which was measured at an earlier age (13 and 14 years), were lower (3.25–3.45 km s
−1). At Tarraleah, the only site where both core and wedge basic density were assessed, basic density in the radial cores was lower than that in the wedge sectors, but this was confounded by age (nine years for cores
vs. 13 and 14 for wedge sectors) and by sampling height (0.9 m
vs. 5.8 m) differences. The Meunna site, where thinning and pruning had been undertaken, had the largest diameter trees (
Table 4). Of the unpruned and unthinned sites, Southport, with a soil developed from a sandstone parent material, was clearly the least productive, although survival at age 19 years was much higher at this site (90%) when compared to Florentine (65%) (
Table 2). While survival at Tarraleah at age 14 years (86%) was higher than Florentine (65%) (the other cool moist site) at age 19 years, analysis of earlier measurements showed that at age 14 years the Florentine site had a very similar survival rate (87%) to that of the Tarraleah site.
Significant differences were observed among the races for AWV and DBH at all sites and for wedge basic density at Southport (
Table 4). The interaction between site and race was not significant for standing tree AWV or core basic density and, despite a significant interaction (
p < 0.001), there were no changes in race ranking across sites for DBH (
Figure 2). For sites and traits where significant differences were observed among races, the Southern race had the highest values and Connor’s Plain the lowest values (
Figure 2).
All traits were observed to be under significant within-race additive genetic control (
Table 4). Estimated narrow-sense heritabilities were generally moderate to high for standing tree AWV (0.39–0.74), except at Meunna (0.16); low (0.09–0.23) for DBH; and moderate for both core and wedge basic density (0.35–0.43). Estimated coefficients of additive variation (
Table 4) were relatively stable across all trials and low for standing tree AWV, core basic density and wedge basic density (2.6%–4.6%), and moderate for DBH (10.0%–14.4%). Similar trends were observed in evolvability: 1.0%–4.1% for AWV, 2.6%–2.9% for basic density and 3.2%–6.1% for DBH.
Figure 2.
Race means at the four trial sites for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m). Some traits were assessed at different ages across sites (see
Table 1). The significance of the race by site interaction effect is shown in parenthesis (
ns not significant,
* p < 0.05,
** p < 0.01,
*** p < 0.001).
Figure 2.
Race means at the four trial sites for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m). Some traits were assessed at different ages across sites (see
Table 1). The significance of the race by site interaction effect is shown in parenthesis (
ns not significant,
* p < 0.05,
** p < 0.01,
*** p < 0.001).
All inter-site additive genetic correlations (
rg) involving the same trait were strong (0.61–0.99,
Table 5) and positive, and were significantly different from zero (
p < 0.05). AWV inter-site correlations averaged 0.83, and were not significantly different from one. This compares with an average of 0.80 for DBH, where sample sizes were greater and inter-site correlations were generally significantly different from one. Within sites, estimated additive genetic correlations between standing tree AWV and DBH were significantly different from zero and positive at Florentine and Meunna (
rg 0.71 and 0.51, respectively), but were weak (
rg 0.18 and 0.20, respectively) and not significant at Southport and Tarraleah (
Table 6). A two-tailed likelihood ratio test showed the genetic correlations between AWV and DBH were significantly different among sites (
p = 0.023).
Table 4.
Estimated race means, significance of the difference among races, within-race additive variance, narrow-sense heritability (
![Forests 05 00744 i012]()
) and coefficient of additive variance (
![Forests 05 00744 i011]()
) and evolvability expressed as a percentage for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) at the four
Eucalyptus nitens progeny trial sites (standard errors shown in parentheses).
Table 5.
Inter-site additive genetic correlations among the four Eucalyptus nitens progeny trial sites for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) (standard errors shown in parentheses). The significance shown is for the test from one. All correlations were highly significantly different from zero (p < 0.001) except for the standing tree AWV correlation between Southport and Meunna (0.61), which was not significant at the p < 0.05 level.
Table 5.
Inter-site additive genetic correlations among the four Eucalyptus nitens progeny trial sites for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) (standard errors shown in parentheses). The significance shown is for the test from one. All correlations were highly significantly different from zero (p < 0.001) except for the standing tree AWV correlation between Southport and Meunna (0.61), which was not significant at the p < 0.05 level.
| Trait | Site | Florentine | Meunna | Southport |
|---|
| AWV (km s−1) | Meunna | 0.86 (0.26) ns | | |
| | Southport | 0.87 (0.19) ns | 0.61 (0.36) ns | |
| Tarraleah | 0.95 (0.13) ns | 0.68 (0.25) ns | 0.99 (0.11) ns |
| Core BD (kg m−3) | Tarraleah | | 0.76 (0.18) ns | |
| Wedge BD (kg m−3) | Tarraleah | | | 0.70 (0.19) ns |
| DBH (cm) | Meunna | 0.79 (0.11) * | | |
| | Southport | 0.71 (0.11) ** | 0.69 (0.10) ** | |
| | Tarraleah | 0.93 (0.08) ns | 0.84 (0.07) ** | 0.86 (0.08) * |
As expected, the additive genetic correlation between earlier age core basic density and later age wedge basic density at Tarraleah was strong (
rg 0.99 ± 0.24) and significantly different from zero (
p < 0.001). Additive genetic correlations between standing tree AWV and basic density were positive (
rg 0.70–0.87) and significantly different from zero (
p < 0.001), with the exception of Meunna (
rg 0.15) which was not significantly different from zero (
Table 6). Furthermore, despite the substantially weaker correlation observed at Meunna, the additive genetic correlations between AWV and basic density were not significantly different across sites. When this correlation was forced to be equal across sites, it was 0.65 (± 0.20) across Meunna and Tarraleah for core basic density and 0.73 (± 0.08) across Tarraleah and Southport for wedge basic density. Additive genetic correlations between DBH and basic density traits were weak and not significantly different from zero (
rg −0.18–0.15).
3.2. Discussion
In this comprehensive study of the genetic control of AWV in eucalypts, we have shown that standing tree AWV in
E. nitens is under strong genetic control and genetic differences are relatively stable across diverse growing environments at the race and within-race levels. This conclusion is based on the growth of open-pollinated progenies from the Central Victorian races of
E. nitens grown on sites covering the range of altitudes (120 m to 600 m), over which most plantations of the species are grown on the island of Tasmania [
26]. The weakest inter-site additive genetic correlation for AWV was between Southport and Meunna (
rg 0.61). These trials were grown under highly divergent silvicultural regimes, which exacerbated substantial climatic and soil differences between the sites. However, even in this extreme case, the additive genetic correlation across sites was not significantly different from one.
Table 6.
Inter-trait additive genetic (rg) and phenotypic (rp) correlations among acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) within each of the four Eucalyptus nitens progeny trial sites (standard errors shown in parentheses). The significance shown is for the test from zero. Trial age in years of each trait assessment is shown as subscripted text.
Table 6.
Inter-trait additive genetic (rg) and phenotypic (rp) correlations among acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) within each of the four Eucalyptus nitens progeny trial sites (standard errors shown in parentheses). The significance shown is for the test from zero. Trial age in years of each trait assessment is shown as subscripted text.
| Site | Trait | AWV (km s−1) | DBH (cm) |
|---|
| | | (Meunna19 Southport19 Florentine19 Tarraleah13,14) | | |
| | | rg | rp | rg | rp |
| Meunna | Core BD9 (kg m−3) | 0.15 (0.35) ns | 0.24 (0.08) ** | −0.18 (0.21) ns | −0.02 (0.05) ns |
| | DBH19 (cm) | 0.51 (0.19) ** | 0.19 (0.03) *** | | |
| Southport | Wedge BD19 (kg m−3) | 0.87 (0.17) *** | 0.37 (0.04) *** | 0.15 (0.23) ns | −0.15 (0.05) ** |
| | DBH19 (cm) | 0.18 (0.21) ns | 0.04 (0.04) ns | | |
| Tarraleah | Core BD9 (kg m−3) | 0.78 (0.22) ** | 0.63 (0.23) * | 0.12 (0.18) ns | 0.11 (0.05) * |
| | Wedge BD14 (kg m−3) | 0.70 (0.10) *** | 0.23 (0.03) *** | 0.24 (0.15) ns | −0.04 (0.04) ns |
| | DBH14 (cm) | 0.20 (0.13) ns | 0.08 (0.03) * | | |
| Florentine | DBH19 (cm) | 0.71 (0.12) *** | 0.21 (0.03) *** | | |
For the three sites that were neither pruned nor thinned, the additive genetic correlations for AWV across sites were very strong (
rg 0.87 to 0.99), despite marked differences in site productivity (see DBH in
Table 4). Our average additive genetic correlation across sites for standing tree AWV (
rg 0.83) was comparable to that observed for other traits in our study (DBH 0.80 and density 0.73). Our average for DBH was 13% higher than the average of DBH across-site genetic correlation estimates reported in the world-wide literature analyses (106 studies) of Hamilton and Potts [
6], which is not surprising given that G × E has been tested with a greater diversity of race material across wider environmental ranges in some studies [
7].
Where significant G × E at the race level was detected it did not usually result in a change in rank order (
Figure 2) and in no case was a change in rank observed in a trait where significant differences among races were evident at the aberrant site, suggesting that scale changes are the predominant drivers of G × E interaction for the traits studied [
17]. Thus rank changes in performance would not be expected if the same genotypes established in these trials were deployed across similar cool-moist and warm-moist climate zones in Tasmania, which represent the majority of the Tasmanian
E. nitens plantation estate [
26]. Notably, the AWV of the Southern race was higher than the other two Central Victorian races across all sites. This is consistent with the Southern race’s previously reported superior performance for several traits relevant to veneer [
24], as well as solid wood and pulpwood [
35] traits.
The genetic stability of AWV rankings across sites (
Table 5) bodes well for the use of AWV as a cost effective predictor of MOE in standing trees of
E. nitens, given that strong genetic correlations between standing tree AWV and static MOE in plywood products has previously been observed at Tarraleah [
24,
25]. To our knowledge there are no other published studies of G × E for AWV in eucalypts, although direct measures of MOE in
Pinus radiata from two sites in southern NSW; one a cool moist site the other a warmer drier site (mean annual temperatures and rainfall, 10.9 °C, 1270 mm and 12.4 °C, 930 mm respectively) showed no significant G × E effect was present [
40]. However, the stability of MOE predictions derived from AWV across sites is not well understood [
41], and requires further investigation before AWV is more widely adopted as a common selection criterion in
E. nitens breeding programs.
The lower AWV observed at the Tarraleah site was most likely due to the younger assessment age compared to the other sites, which were of the same age and had similar AWVs. An increase in AWV with age is consistent with previous studies that have found that MOE increases with cambial age [
42]. In studies of ten year-old
E. nitens, Chauhan and Walker [
43] found that the MOE of outerwood was significantly greater (about 56%) than the MOE of corewood. This trend was also evident in a study of the closely related species
E. globulus [
44] and other temperate eucalypt species [
45], where radial variation in MOE was predicted from studies of micro-fibril angle and density. Basic density has also been shown to increase with cambial age and height in eucalypts [
46] and variation in sampling position (5.8 m for the wedges compared with 0.9 m for cores) and sampling age (14 years for wedges and 6 years for cores) is likely to explain the greater density of wedges compared with cores in our study.
The level of genetic control of standing tree AWV (mean
![Forests 05 00744 i012]()
= 0.43;
Table 4) within races was nearly three-fold higher than that for DBH (0.15). The evolvability of AWV (
Table 4) was on average similar to that observed for basic density but slightly lower than that for DBH, indicating that lower genetic gains would be expected in AWV and basic density than in DBH, assuming a fixed selection intensity applied to a single trait under phenotypic selection. The later-age estimates of narrow-sense heritabilities for standing tree AWV from the Florentine and Southport trial (
Table 4) were high and similar to previous estimates for
E. dunnii (0.42),
E. pilularis (0.65) [
47] and
Pinus radiata (0.67) [
48]. The highest narrow-sense heritability we obtained was from the Tarraleah trial, which was expected because it combined data from two previous studies, which reported high heritabilities, each of which was based on trees from three different replicates of this trial [
24,
25]. High heritabilities have also been shown in studies of direct and indirect measures of stiffness and strength in
Pinus radiata [
49]. The lower narrow-sense heritability at Meunna, compared to the other three
E. nitens trials studied is difficult to explain. It is possible that thicker bark on larger stems at Meunna could have increased measurement error and decreased heritability at this site. It is further possible that thinning and pruning at this site reduced the expression of additive genetic variation in AWV [
50]. In any case, the very strong inter-site genetic correlations observed for all traits (
Table 5) suggests that genetic gains achieved through selection at any one site would, to a large extent, be transferable to environments and silvicultural regimes represented by the other sites [
20].
The strong, highly significant and positive additive genetic correlations between standing tree AWV and basic density (assessed using either wedges or cores) observed at the Tarraleah and Southport sites (
Table 6), supports the use of standing tree AWV as a selection criterion for the improvement of basic density in
E. nitens. A strong and positive additive genetic correlation between these traits has also been observed in other species (e.g. 0.57 in
Pinus radiata [
51]). Consistent with the observations for G × E and narrow-sense heritability, the main deviant inter-trait additive genetic correlation involving AWV and density in the present study occurred at the thinned and pruned Meunna site where the correlation was close to zero, albeit not significantly different from the strong correlations observed at Tarraleah and Southport. Although positive and significantly different from zero, phenotypic correlations between AWV and basic density in the present study were generally weaker than those observed in
E. dunnii by Dickson
et al. [
52] (0.67 at age nine years and 0.71 at age 25 years).
The Tarraleah site had the lowest rainfall but was also the coolest site with deep, fertile soil derived from Tertiary basalt, which would contribute to reducing evaporation and improving water holding capacity and therefore the observed good growth. The site of lowest productivity was Southport, with the lowest DBH at age 19 years. This site had the lowest effective rainfall (annual precipitation minus annual evaporation) (
Table 1). The soil at Southport, which has poor moisture retention, being derived from Triassic sandstone, may have also contributed to the slower growth there. In studies on
E. nitens in Chile, the soil type was shown to have a major impact on site productivity [
53]. Meunna had the largest diameter trees and had the highest rainfall and temperature on deep, fertile soil derived from Tertiary basalt, although growth would have been substantially increased by thinning at age 4 years. Our study indicates that there is a weak positive additive genetic correlation between AWV and DBH, but suggests that the strength of this favourable relationship is not stable across sites. Other studies in
E. nitens [
24] and
Pinus taeda [
54] have shown no significant genetic correlation between AWV and DBH. The positive genetic correlations of AWV with both basic density and DBH (
Table 6) occur despite the absence of a significant correlation between DBH and density, and indicates that past selection for fast growth and high density in pulpwood breeding programs is unlikely to have adversely affected MOE [
24].



{kind=link}
{kind=link}

 ) [3], open-pollinated narrow-sense heritability (
) [3], open-pollinated narrow-sense heritability (  ) [4], percentage coefficient of additive variation (%CVadd) [5], and a dimensionless measure of evolvability [38] were estimated for each trait from univariate site analyses as follows:
) [4], percentage coefficient of additive variation (%CVadd) [5], and a dimensionless measure of evolvability [38] were estimated for each trait from univariate site analyses as follows:




 and
and  are the variance components for ADD(RACE) and RESIDUAL respectively.
are the variance components for ADD(RACE) and RESIDUAL respectively.
 is the ADD(RACE) covariance component between traits 1 and 2, and
is the ADD(RACE) covariance component between traits 1 and 2, and 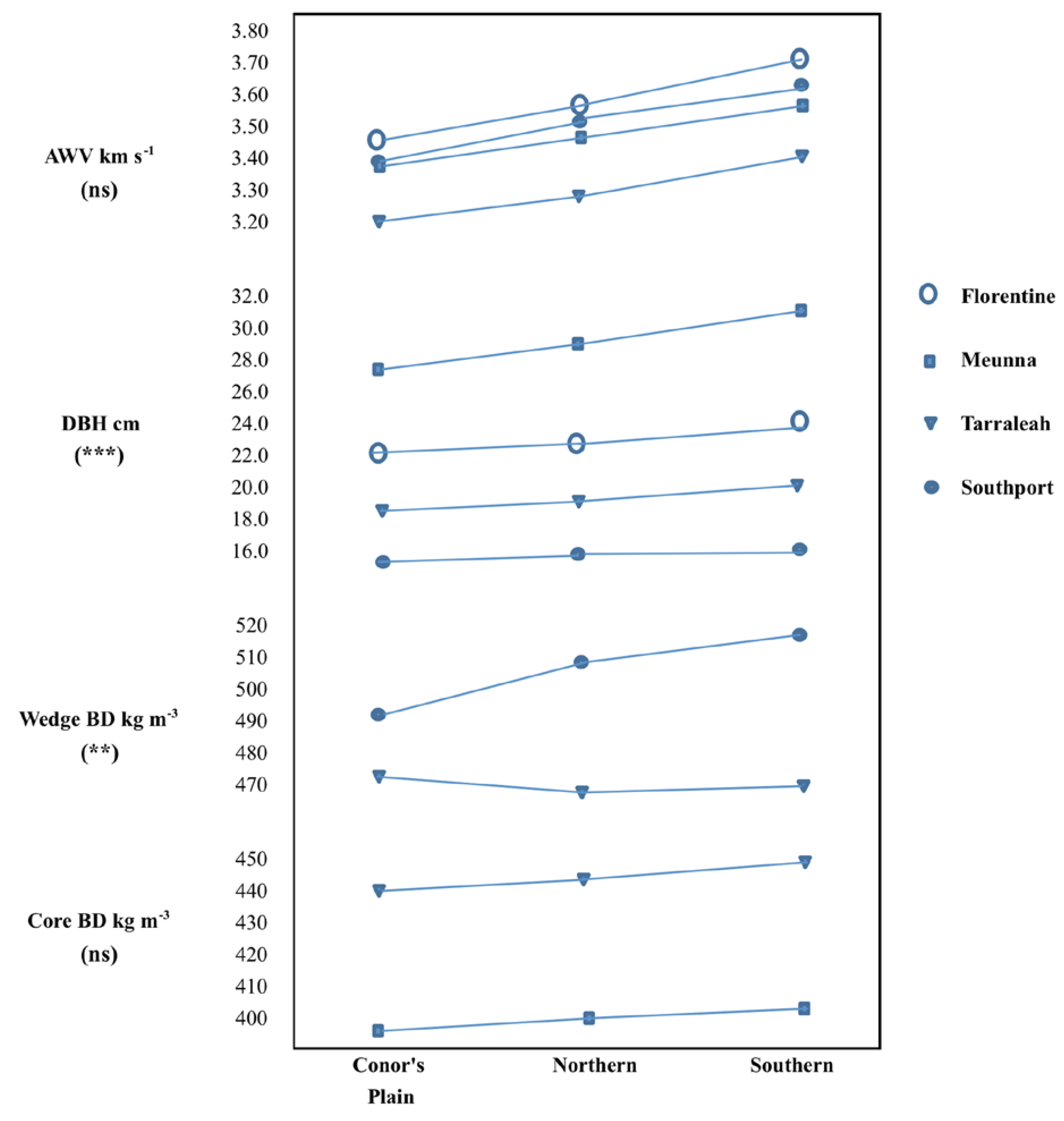
 ) and coefficient of additive variance (
) and coefficient of additive variance (  ) and evolvability expressed as a percentage for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) at the four Eucalyptus nitens progeny trial sites (standard errors shown in parentheses).
) and evolvability expressed as a percentage for acoustic wave velocity (AWV), basic density (BD) and diameter at breast height (DBH; 1.3 m) at the four Eucalyptus nitens progeny trial sites (standard errors shown in parentheses).