
Carbon Budgets for Caribbean Mangrove Forests of Varying Structure and with Phosphorus Enrichment

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Site


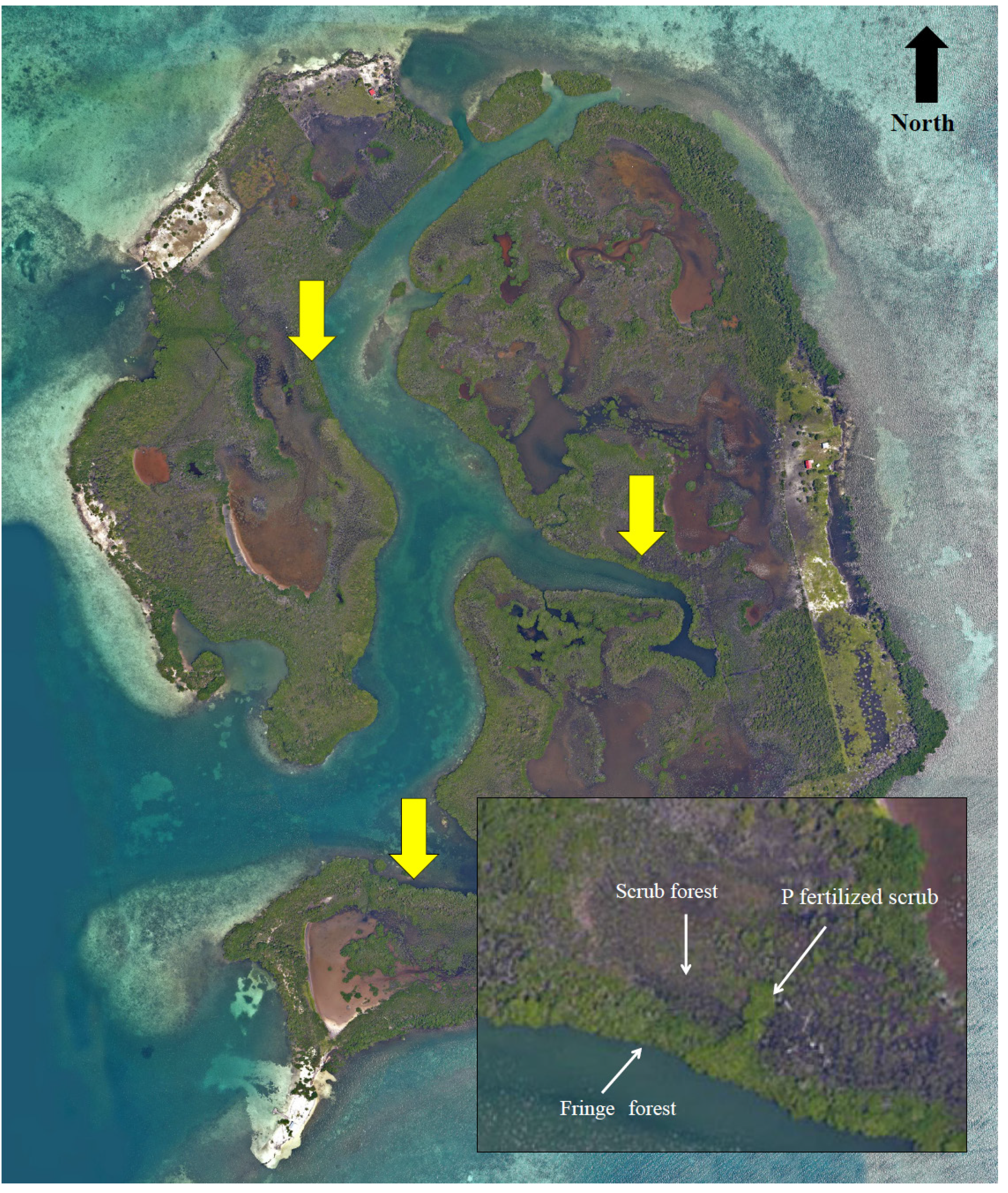{kind=link}
{kind=link}
| Forest Attributes | Forest Type | ||
|---|---|---|---|
| Fringe | Scrub | P Fertilized Scrub | |
| Density of trees No. m−2 (N = 5) | 0.51 ± 0.23 | 0.23 ± 0.09 | 0.23 ± 0.09 |
| Tree Height (N = 9) | 3.3 ± 0.2 | 1.19 ± 0.10 | 2.24 ± 0.22 |
| LAI m2·m−2 (N = 9) | 1.82 ± 0.07 | 0.39 ± 0.06 | 1.70 ± 0.77 |
| Direct measure of canopy leaf area (m2·m−2) (N = 9) | 2.62 ± 0.30 | 1.17 ± 0.33 | 4.61 ± 0.94 |
| Surface area stems (m2 tree) | 1.3 ± 0.3 | 0.06 ± 0.01 | 0.46 ± 0.09 |
| Aboveground root length (m length per tree) (N = 9) | 62 ± 20 | 7.4 ± 4.0 | 39 ± 16 |
| Surface area aboveground roots (m2 root area m−2) | 5.4 ± 1.7 | 0.59 ± 0.29 | 2.8 ± 1.1 |
| Biomass of macroalgae g·m−2 (N = 9) | 40 ± 20 | 8 ± 8 | 24 ±13 |
| Surface area of habitat for macroalgae (m2 root area m−2) (N = 9) | 0.68 ± 0.14 | 0.22 ± 0.02 | 0.53 ± 0.08 |
| Leaf photosynthetic carbon gain µmol·m−2·s−1 (N = 9) | 10.3 ± 0.4 | 7.4 ± 0.1 | 12.1 ± 0.7 |
| Leaf respiration µmol·m−2·s−1 (N = 9) | 1.41 ± 0.06 | 1.61 ± 0.04 | 1.98 ± 0.12 |
2.2. Primary Productivity and Respiration of the Canopy, Mats and Macroalgae
2.3. Other Outputs: Litter Export, Herbivory, Respiration of Woody Tissues and Soils
2.4. Data Analysis
3. Results and Discussion
3.1. Gross Primary Productivity
| Budget Components | Forest Type | ||
|---|---|---|---|
| Fringe | Scrub | Fertilized P Scrub | |
| INPUTS | |||
| Mangrove trees | 378.0 | 89.0 | 555.0 |
| Epiphytic algal community | 38.0 | 7.5 | 22.0 |
| Microbial mat | 11.0 | 36.5 | 11.0 |
| TOTAL INPUTS | 427.0 | 133.0 | 588.0 |
| OUTPUTS | |||
| Canopy respiration | 132.0 | 51.0 | 286.0 |
| Soil respiration | 47.0 | 8.6 | 81.5 |
| Aboveground root respiration | 108.0 | 29.0 | 168.0 |
| Stem respiration | 18.6 | 3.2 | 36.5 |
| Algal respiration | 8.2 | 1.6 | 4.8 |
| Wood production | 11.6 | 0.1 | 15.0 |
| Herbivory | 1.2 | 0.4 | 1.6 |
| Burial in soil | 17.6 | 2.8 | 12.9 |
| Export (litter) | 13.1 | 0.8 | 13.1 |
| TOTAL OUTPUTS | 357.3 | 97.5 | 619.4 |
| Net Ecosystem Production (NEP) (% GPP) | 69.6 (16.3) | 35.5 (26.7) | −31.4 (−5.3%) |
| % C of GPP buried | 4.1 | 2.1 | 2.2 |
| Production:Respiration ratio (P:R) | 1.19 | 1.36 | 0.95 |
3.2. Autotrophic Respiration
3.3. Soil Respiration
| Budget Components | Forest Type | ||
|---|---|---|---|
| Fringe | Scrub | P fertilized Scrub | |
| Soil Respiration | 61.2 | 34.7 | 92.7 |
| Root respiration (stock × respiration rate) | 64.2 | 6.9 | 102.4 |
| Microbial respiration (by difference) (% of soil respiration) | −3.0 (−5%) | 27.8 (80%) | −9.7 (−10%) |
3.4. Carbon Burial

3.5. Variation in Net Ecosystem Production
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global carbon sequestration in tidal, saline wetland soils. Glob. Biogeochem. Cycles 2003, 17, 1111. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling and storage in mangrove forests. Annu. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Bjork, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B. A Blueprint for Blue Carbon: Towards an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marba, N.; et al. Estimating global “Blue Carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [Green Version]
- Siikamäki, J.; Sanchirico, J.N.; Jardine, S.L. Global economic potential for reducing carbon dioxide emissions from mangrove loss. Proc. Natl. Acad. Sci. USA 2012, 109, 14369–14374. [Google Scholar] [CrossRef]
- Jennerjahn, T.C.; Ittekkot, V. Relevance of mangroves for the production and deposition of organic matter along tropical continental margins. Naturwissenschaften 2002, 89, 23–30. [Google Scholar] [CrossRef]
- Dittmar, T.; Hertkorn, N.; Kattner, G.; Lara, R.J. Mangroves, a major source of dissolved organic carbon to the oceans. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castaneda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. Carbon sequestration in mangrove forests. Carbon Manag. 2012, 3, 313–322. [Google Scholar] [CrossRef]
- Cebrian, J.; Duarte, C.M. Plant growth-rate dependence of detrital carbon storage in ecosystems. Science 1995, 5217, 1606–1608. [Google Scholar] [CrossRef]
- Alongi, D.M. Energetics of Mangrove Ecosystems; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Feller, I.C.; McKee, K.L.; Whigham, D.F.; O’Neill, J.P. Nitrogen vs. phosphorus limitation across and ecotonal gradient in a mangrove forest. Biogeochemistry 2003, 62, 145–175. [Google Scholar] [CrossRef]
- McKee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation. Glob. Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Wooller, M.J.; Behling, H.; Smallwood, B.J.; Fogel, M. Mangrove ecosystem dynamics and elemental cycling at Twin Cays, Belize, during the Holocene. J. Quat. Sci. 2004, 19, 703–711. [Google Scholar] [CrossRef]
- Maher, D.T.; Eyre, B.D. Carbon budgets for three autotrophic Australian estuaries: Implications for global estimates of the coastal air-water CO2 flux. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Alongi, D.M. Mangrove forests: Resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 2002, 76, 1–13. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Ruess, R.W.; Feller, I.C. CO2 efflux from cleared mangrove peat. PLoS ONE 2011, 6, e21279. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, J.B.; Heider, C.; Norfolk, J.; Payton, F. Carbon stocks of intact mangroves and carbon emissions arising from their conversion in the Dominican Republic. Ecol. Appl. 2014, 24, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S. Mineral nutrition of wild plants. Annu. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Tilman, D. Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol. Monogr. 1987, 57, 189–214. [Google Scholar] [CrossRef]
- Feller, I.C.; Lovelock, C.E.; Piou, C. Growth and nutrient conservation in Rhizophora mangle in response to fertilization along latitudinal and tidal gradients. Smithson. Contrib. Mar. Sci. 2009, 38, 345–358. [Google Scholar]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. The biocomplexity of mangrove ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–416. [Google Scholar] [CrossRef]
- Ruetzler, K.; Feller, I.C. Caribbean mangrove swamps. Sci. Am. 1996, 274, 94–99. [Google Scholar] [CrossRef]
- Macintyre, I.G.; Littler, M.M.; Littler, D.S. Holocene history of Tobacco Range, Belize, Central America. Atoll Res. Bull. 1995, 43, 1–18. [Google Scholar] [CrossRef]
- Woodroffe, C.D. Mangrove vegetation of Tobacco Range and nearby mangrove ranges, central Belize barrier reef. Atoll Res. Bull. 2005, 427, 1–35. [Google Scholar]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Herrera-Silveira, J.A. Carbon stocks of tropical coastal wetlands within the karstic landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef] [PubMed]
- Middleton, B.A.; McKee, K.L. Degradation of mangrove tissues and implications for peat formation in Belizean island forests. J. Ecol. 2001, 89, 818–828. [Google Scholar] [CrossRef]
- Rodriguez, W.; Feller, I.C. Mangrove landscape characterization and change in Twin Cays, Belize using aerial photography and IKONOS satellite data. Atoll Res. Bull. 2004, 513, 1–24. [Google Scholar] [CrossRef]
- Lee, R.Y.; Joye, S.B. Patterns and controls on nitrogen fixation and denitrification in intertidal soils of a tropical oceanic mangrove island. Mar. Ecol. Prog. Ser. 2006, 307, 127–41. [Google Scholar] [CrossRef]
- Koltes, K.; Tschirky, J.; Feller, I.C. Carrie Bow Cay, Belize. In Caribbean Coastal Marine Productivity (CARICOMP): Caribbean Coral Reef, Seagrass and Mangrove Sites Characteristics; Coastal Region and Small Island Papers 3; Kjerfve, B., Ed.; UNESCO: Paris, France, 1998; pp. 79–94. [Google Scholar]
- Feller, I.C. Effects of nutrient enrichment on growth and herbivory of dwarf red mangrove Rhizophora mangle. Ecol. Monogr. 1995, 65, 477–505. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Ball, M.C.; Choat, B.; Engelbrecht, B.M.J.; Holbrook, N.M.; Feller, I.C. Linking physiological processes with mangrove forest structure: Phosphorus deficiency limits canopy development; hydraulic conductivity and photosynthetic carbon gain in dwarf Rhizophora mangle. Plant Cell Environ. 2006, 29, 793–802. [Google Scholar] [CrossRef]
- Cheeseman, J.C.; Lovelock, C.E. Photosynthetic characteristics of dwarf and fringe Rhizophora mangle in a Belizean mangrove. Plant Cell Environ. 2004, 27, 768–780. [Google Scholar] [CrossRef]
- Ball, M.C.; Cowan, I.R.; Farquhar, G.D. Maintenance of leaf temperature and the optimisation of carbon gain in relation to water loss in a tropical mangrove forest. Aust. J. Plant Physiol. 1988, 15, 263–267. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Clough, B.F. Influence of solar radiation and leaf angle on leaf xanthophyll concentrations in mangroves. Oecologia 1992, 91, 518–525. [Google Scholar] [CrossRef]
- Lee, R.Y.; Porubsky, W.P.; Feller, I.C.; McKee, K.L.; Joye, S.B. Porewater biogeochemistry and soil metabolism in dwarf red mangrove habitats (Twin Cays; Belize). Biogeochemistry 2008, 87, 181–198. [Google Scholar] [CrossRef]
- Joye, S.B.; Lee, R.Y. Benthic microbial mats: Important sources of fixed nitrogen and carbon to the Twin Cays, Belize ecosystem. Atoll Res. Bull. 2004, 528, 1–24. [Google Scholar] [CrossRef]
- Genty, B.; Briantas, J.-M.; Baker, N.R. The relationship between quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Feller, I.C.; Chamberlain, A.H. Herbivore responses to nutrient enrichment and landscape heterogeneity in a mangrove ecosystem. Oecologia 2007, 153, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, J.B.; Heider, C.; Cole, T.; Dwire, K.A.; Donato, D.C. Ecosystem carbon pools of Micronesian mangrove forests: Implications of land use and climate change. Wetlands 2011, 31, 343–352. [Google Scholar] [CrossRef]
- Ryan, M.G.; Gower, S.T.; Hubbard, R.M.; Waring, R.H.; Gholz, H.L.; Cropper Jr, W.P.; Running, S.W. Woody tissue maintenance respiration of four conifers in contrasting climates. Oecologia 1995, 101, 133–140. [Google Scholar] [CrossRef]
- Reich, P.B.; Tjoelker, M.G.; Machado, J.L.; Oleksyn, J. Universal scaling of respiratory metabolism, size and nitrogen in plants. Nature 2006, 439, 457–461. [Google Scholar] [CrossRef]
- Hoque, A.T.M.R.; Sharma, S.; Suwa, R.; Hagihara, A. Seasonal variation in the size-dependent respiration of mangroves Kandelia obovata. Mar. Ecol. Prog. Ser. 2010, 404, 31–37. [Google Scholar] [CrossRef]
- Mori, S.; Yamaji, K.; Ishida, A.; Prokushkinc, S.G.; Masyaginac, O.V.; Hagiharad, A.; Hoqued, A.T.M.R.; Suwad, R.; Osawae, A.; Nishizono, T.; et al. Mixed-power scaling of whole-plant respiration from seedlings to giant trees. Proc. Natl. Acad. Sci. 2010, 107, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Lovelock, C.E. Soil respiration in tropical and subtropical mangrove forests. Ecosystems 2008, 11, 342–354. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Ruess, R.W. Variable effects of nutrient enrichment on soil respiration in mangrove forests. Plant Soil 2015, 379, 135–148. [Google Scholar] [CrossRef]
- McKee, K.L. Biophysical controls on accretion and elevation change in Caribbean mangrove ecosystems. Estuar. Coast. Shelf Sci. 2011, 91, 475–483. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Ruess, R.W.; Feller, I.C. Root respiration in Rhizophora mangle over variation in forest stature and nutrient availability. Tree Physiol. 2006, 26, 1601–1606. [Google Scholar] [CrossRef]
- McKee, K.L.; Feller, I.C.; Popp, M.; Wanek, W. Mangrove isotopic fractionation (δ15N and δ13C) across a nitrogen versus phosphorus limitation gradient. Ecology 2002, 83, 1065–75. [Google Scholar]
- Ellison, A.M.; Farnsworth, E.J. Spatial and temporal variability in growth of Rhizophora mangle saplings on coral cays: Links with variation in insolation, herbivory, and local sedimentation rate. J. Ecol. 1996, 84, 717–731. [Google Scholar] [CrossRef]
- Lugo, A.E. Old-growth mangrove forests in the United States. Conserv. Biol. 1997, 11, 11–20. [Google Scholar] [CrossRef]
- Kitaya, Y.; Yabuki, K.; Kiyota, M.; Tani, A.; Hirano, T.; Aiga, I. Gas exchange and oxygen concentration in pneumatophores and prop roots of four mangrove species. Tree Struct. Funct. 2002, 16, 155–158. [Google Scholar] [CrossRef]
- Schmitz, N.; Egerton, J.J.G.; Lovelock, C.E.; Ball, M.C. Light-dependent maintenance of hydraulic function in mangrove branches: Do xylary chloroplasts play a role in embolism repair? New Phytol. 2012, 195, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Material not intended for publication: C.E. Lovelock (The University of Queensland, St Lucia, QLD). Transmittance of light through epidermal tissues of aboveground roots of Rhizophora sp., 2013.
- Drew, M.C.; Saglio, P.H.; Pradet, A. Larger adenylate energy charge and ATP/ADP ratios in aerenchymatous roots of Zea mays in anaerobic media as a consequence of improved internal oxygen transport. Planta 1985, 165, 51–58. [Google Scholar] [CrossRef] [PubMed]
- King, J.S.; Hanson, P.J.; Bernhardt, E.; DeAngelis, P.; Norby, R.J.; Pregitzer, K.S. A multiyear synthesis of soil respiration responses to elevated atmospheric CO2 from four forest FACE experiments. Glob. Chang. Biol. 2004, 10, 1027–1042. [Google Scholar] [CrossRef]
- Adame, M.F.; Lovelock, C.E. Carbon and nutrient exchange of mangrove forests with the coastal ocean. Hydrobiologia 2011, 663, 23–50. [Google Scholar] [CrossRef]
- Day, J.W.; Coronado-Molina, C.; Vera-Herrera, F.R.; Twilley, R.; Rivera-Monroy, V.H.; Alvarez-Guillen, H.; Day, R.; Conner, W. A 7 year record of above-ground net primary production in a southeastern Mexican mangrove forest. Aquat. Bot. 1996, 55, 39–60. [Google Scholar] [CrossRef]
- Feller, I.C. The role of herbivory by wood-boring insects in mangrove ecosystems in Belize. Oikos 2002, 97, 167–76. [Google Scholar] [CrossRef]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How mangrove forests adjust to rising sea level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Castaneda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado-Molina, C.; Ewe, S.M. Patterns of root dynamics in mangrove forests along environmental gradients in the Florida Coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Hogberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–90. [Google Scholar] [CrossRef] [PubMed]
- Keuskamp, J.A.; Schmitt, H.; Laanbroek, H.J.; Verhoeven, J.T.; Hefting, M.M. Nutrient amendment does not increase mineralisation of sequestered carbon during incubation of a nitrogen limited mangrove soil. Soil Biol. Biochem. 2013, 57, 822–829. [Google Scholar] [CrossRef]
- Alongi, D.M.; de Carvalho, N.A.; Amaral, A.L. Uncoupled surface and below-ground soil respiration in mangroves: implications for estimates of dissolved inorganic carbon export. Biogeochemistry 2012, 109, 151–162. [Google Scholar]
- Anon. Carbon Dioxide Information and Analysis Center. Available online: http://cdiac.ornl.gov/trends/emis/blz.html (accessed on 1 September 2015).
- Murray, M.R.; Zismana, S.A.; Furleya, P.A.; Munro, D.M.; Gibson, J.; Ratter, J.; Bridgewater, S.; Minty, C.D.; Place, C.J. The mangroves of Belize Part 1. Distribution, composition and classification. For. Ecol. Manag. 2003, 174, 265–279. [Google Scholar] [CrossRef]
- Material not intended for publication: C.E. Lovelock (The University of Queensland, St Lucia, QLD). Estimates of Gross Primary Production for mangrove forests in Moreton Bay, derived from assessment of LAI and photosynthetic rates of leaves; and estimates of carbon burial derived from assessment of soil accretion and soil carbon density, 2013.
- Lovelock, C.E.; Adame, M.F.; Bennion, V.; Hayes, M.; O’Mara, J.; Reef, R.; Santini, N. Contemporary rates of carbon sequestration through vertical accretion of sediments in mangrove forests and saltmarshes of South East Queensland, Australia. Estuar. Coasts 2013. [Google Scholar] [CrossRef]
- Alongi, D.M. Dissolved iron supply limits early growth of estuarine mangroves. Ecology 2011, 91, 3229–3241. [Google Scholar] [CrossRef]
- Leopold, A.; Marchand, C.; Debordeb, J.; Chaduteau, C.; Allenbach, M. Influence of mangrove zonation on CO2 fluxes at the sediment-air interface (New Caledonia). Geoderma 2012, 202–203, 62–70. [Google Scholar] [CrossRef]
- Bouillon, S.; Middleburg, J.J.; Dehairs, F.; Borges, A.V.; Abril, G.; Flindt, M.R.; Ulomi, S.; Kristensen, E. Importance of intertidal sediment processes and porewater exchange on the water column biogeochemistry in a pristine mangrove creek (Ras Dege, Tanzania). Biogeosciences 2007, 4, 311–322. [Google Scholar] [CrossRef]
- Urish, D.W.; Wright, R.M.; Feller, I.C.; Rodriguez, W. Dynamic hydrology of a mangrove island: Twin Cays, Belize. Smithson. Contrib. Mar. Sci. 2009, 38, 473–490. [Google Scholar]
- Feller, I.C.; (Smithsonian Environmental Research Center, Edgewater, MD, USA). Aerial photographs from light aircraft. 2011. [Google Scholar]
- Feller, I.C.; Baker, R.; (Smithsonian Environmental Research Center, Edgewater, MD, USA). Stable isotope composition of seagrass at Twin Cays. 2013. [Google Scholar]
- Lin, G.; Banks, T.; Sternberg, L.S.L.O. Variation in δ13C values for the seagrass Thalassia testudinum and its relations to mangrove carbon. Aquat. Bot. 1991, 40, 333–341. [Google Scholar] [CrossRef]
- Raven, J.A.; Johnston, A.M.; Kübler, J.E.; Korb, R.; McInroy, S.G.; Handley, L.L.; Scrimgeour, C.M.; Walker, D.I.; Beardall, J.; Vanderklift, M.; et al. Mechanistic interpretation of carbon isotope discrimination by marine macroalgae and seagrasses. Funct. Plant Biol. 2002, 29, 355–378. [Google Scholar] [CrossRef]
- Barr, J.G.; Engel, V.; Fuentes, J.D.; Zieman, J.C.; O’Halloran, T.L.; Smith, T.J.; Anderson, G.H. Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef]
- Maher, D.T.; Santos, I.R.; Golsby-Smith, J.; Gleeson, J.; Eyre, B.D. Groundwater-derived dissolved inorganic and organic carbon exports from a mangrove tidal creek: The missing mangrove carbon sink? Limnol. Oceanogr. 2013, 58, 475–488. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lovelock, C.E.; Simpson, L.T.; Duckett, L.J.; Feller, I.C. Carbon Budgets for Caribbean Mangrove Forests of Varying Structure and with Phosphorus Enrichment. Forests 2015, 6, 3528-3546. https://doi.org/10.3390/f6103528
Lovelock CE, Simpson LT, Duckett LJ, Feller IC. Carbon Budgets for Caribbean Mangrove Forests of Varying Structure and with Phosphorus Enrichment. Forests. 2015; 6(10):3528-3546. https://doi.org/10.3390/f6103528
Chicago/Turabian StyleLovelock, Catherine E., Lorae T. Simpson, Lisa J. Duckett, and Ilka C. Feller. 2015. "Carbon Budgets for Caribbean Mangrove Forests of Varying Structure and with Phosphorus Enrichment" Forests 6, no. 10: 3528-3546. https://doi.org/10.3390/f6103528