1. Introduction
Mountain pine beetle,
Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae, Scolytinae), is one of the most serious bark beetle pests of lodgepole pine,
Pinus contorta Douglas, in western North America [
1]. Because of the beetle’s wide-ranging distribution (from northern Mexico, throughout the western United States and into southwestern Canada), local populations of the beetle may encounter very different environmental conditions and/or host populations. However, when mountain pine beetle outbreaks occur, they can extend over years, during which time the beetle can cause extensive tree mortality [
2,
3]. Mountain pine beetle attacks occur more frequently in stands with higher basal area and stand density index measurements [
4]. Further, successfully attacked trees tend to be dominant or co-dominant trees with a larger diameter then adjacent trees [
4,
5,
6].
Conifer resistance to attack by bark beetles or other bole-infesting organisms is comprised of a three stage response of wound cleansing, infection containment and wound healing [
7,
8]. The first step of this sequence, wound cleansing, involves the flow of constitutive resins at the point of attack(s). When exuded in high enough concentrations, the resins can trap and kill attacking beetles before they colonize the tree. Bark beetles attacking ponderosa pine have been shown to preferentially attack trees with fewer resin ducts [
9] and lodgepole pines that successfully resisted attack by mountain pine beetle had more resin ducts in recent growth then did trees that were killed by the beetle [
10].
Many of the forested areas in the Intermountain West are deficient in multiple nutrients including nitrogen (N), potassium (K), sulfur (S) and/or boron (B) [
11,
12]. Lodgepole pine on these sites responds to N-only or N plus K and/or S fertilization with an increase in N content of a tree’s inner bark tissue [
13], a change in foliar chemistry and significantly increased growth [
14,
15]. However, a significant decline in K/N ratios can occur when trees are fertilized with N-only treatments which can result in susceptibility to insect pests [
11]. Further, while N fertilization did increase the growth rate of mature stands of lodgepole pine, it did not prevent attack by mountain pine beetle until a threshold of tree growth efficiency was exceeded [
14]. Higher N content within a host pine can result in higher N content of bark beetles reared in those trees [
13,
16]. Higher N content trees produce larger adults [
16] and the increased N availability expressed in the beetles results in increased progeny production and adult emergence [
17].
The nutrient deficiencies of Intermountain West forest soils provide a basis for understanding the interaction between lodgepole pine and mountain pine beetle based upon several theories that examine chemically-mediated interactions between insects and their host plant. For example, optimal defense theory assumes that defenses against herbivores are costly to a plant because they divert resources away from growth [
18]. Similarly, the fundamental principle of the growth-differentiation balance hypothesis is the existence of a physiological tradeoff between growth and differentiation processes (including the maintenance of resistance/defense chemicals and mechanisms) [
19,
20]. The carbon-nutrient balance theory hypothesizes that nutrient availability to a plant is a major factor in limiting plant growth [
21]. Further, a low availability of nutrient resources should favor the production and maintenance of substantive constitutive defenses so as to limit herbivory in resource-poor environments [
22,
23,
24]. While resin flow is a physical (as opposed to chemical) resistance parameter, one of the toxic components of resin are monoterpenes which are C-based secondary metabolites. Prior studies have demonstrated a decrease in C-based secondary metabolites following fertilization [
25,
26] and loblolly pine,
P. taeda L. has its lowest measurements of resin flow during periods of rapid tree growth [
27].
The primary objective of the current project was to determine the impact of N fertilization (with and without the addition of other nutrients) plus thinning on a constitutive defense (resin flow) of the tree against bole-attacking bark beetles. While accomplishing the primary objective, we also wanted to determine if changes in resin flow on the tree bole was related to changes in foliar nutrient content following fertilization and thinning.
2. Methods
Two experiments were conducted, both on the University of Idaho’s Experimental Forest located in Latah County, Idaho, U.S.A. Both stands were mixed-conifer forests, consisting of mature (at least 40 year old) lodgepole pine, ponderosa pine (
P. ponderosa (Douglas), Douglas-fir (
Pseudotsuga menziesii (Mirb.) Franco) and grand fir (
Abies grandis (Douglas ex D. Don) Lindley). The dominant vegetation series for both stands is characterized as
Abies grandis with
Thuja plicata (Donn ex D. Don) occurring locally in microsites. Soil parent material is derived from granitic geology with a thin mantle of ash and loess and has a soil taxonomic classification of Vitrandic Fragixeralf. Site topography is expressed as a northeasterly aspect with slopes <10% at an elevation of 975 m. Within an experiment, the lodgepole pines selected for investigation were all dominant or co-dominant trees that were similar in size (
Table 1) based upon dbh measurements (Experiment 1:
f = 0.14;
df = 2, 23; (
p >
f) = 0.08694 and Experiment 2:
f = 0.65;
df = 7, 33; (
p >
f ) = 0.7086).
Table 1.
Treatment and tree diameters (cm + SEM) for the lodgepole pines in two nitrogen fertilization experiments. Individual trees in Experiment 1 received 1 of 3 treatments (0, 200 or 400 kg/ha of N applied as urea). Trees in Experiment 2 were in stands that either received no thinning treatment or were thinned to a stand density of approximately 40 trees/ha. Individual trees were then fertilized with N applied as urea at 0, 227, 454 or 227 (plus additional nutrients) kg/ha.
Table 1.
Treatment and tree diameters (cm + SEM) for the lodgepole pines in two nitrogen fertilization experiments. Individual trees in Experiment 1 received 1 of 3 treatments (0, 200 or 400 kg/ha of N applied as urea). Trees in Experiment 2 were in stands that either received no thinning treatment or were thinned to a stand density of approximately 40 trees/ha. Individual trees were then fertilized with N applied as urea at 0, 227, 454 or 227 (plus additional nutrients) kg/ha.
| Nitrogen (kg/ha) | Experiment 1 | Experiment 2 |
|---|
| Stand Thinned | n | dbh (cm) | n | dbh (cm) |
|---|
| 0 | no | 8 | 30.6 ± 1.5 | 5 | 28.6 ± 1.7 |
| yes | | | 5 | 29.3 ± 0.7 |
| 227 | no | 9 | 29.7 ± 2.1 | 5 | 28.3 ± 0.9 |
| yes | | | 5 | 30.8 ±1.8 |
| 454 | no | 9 | 29.1 ± 2.0 | 5 | 29.6 ± 2.5 |
| yes | | | 5 | 26.3 ± 1.4 |
| 227 ± (K, S, B) | no | | | 5 | 28.8 ± 1.8 |
| | | | 5 | 27.3 ± 2.2 |
Twenty-six lodgepole pines were used in the first experiment. Trees were a minimum of 30.5 m apart and were randomly assigned to one of three fertilization treatments (eight trees were non-fertilized controls, and 18 trees received N fertilizer that was applied as urea). Of the trees that were fertilized, nine received the equivalent of 227 kg N and nine received the equivalent of 454 kg N per ha. Using a hand spreader, urea was applied to individual trees at the designated rates on 15 October 2006 to an area of ground corresponding to twice the diameter of the crown.
Resin flow from each tree was measured the following summer (July 2007) using calibrated receptacles to capture resin flowing from the wounded inner bark for 48 h [
28]. Among treatment resin flow was compared using analysis of variance techniques with a protected Fisher’s least significant difference test [
29].
Forty lodgepole pines were used in the second experiment. Twenty of the experimental trees had the trees in the stand directly surrounding them removed during the summer (June–August, 2007). Densities prior to thinning were approximately 500 trees/ha and the post-thinning densities were approximately 200–225 trees/ha. All experimental trees were a minimum of 30.5 m apart and were randomly assigned to one of four fertilization treatments (non-fertilized controls, low (227 kg/ha) and high (454 kg/ha) N or a blend of 227 kg/ha of N plus potassium (193 kg/ha), sulfur (102 kg/ha) and boron (3.4 kg/ha). Treatments (urea with/without amendments) were applied during October 2007. Treatment amounts were calculated and applied based upon twice the area of green crown for the individual trees.
To determine individual tree growth, all of the study trees were measured for dbh in August 2007 and again in July 2008. Resin flow from each tree was measured during July 2008 [
28]. Current-year foliage was removed from two limbs per tree at a minimum height of 10 m using a 12-gauge shotgun with steel shot in July 2008. Foliage was collected from trees just prior to the dormant season (October 2008). Current-year lateral shoot growth was collected from the third whorl from the top of each selected tree using a pruning pole or by removing limbs using a shotgun with steel shot. Samples were placed in plastic bags, stored in an ice-cooled container, and transported to the laboratory for deep-freeze storage until tissue processing in the laboratory. Foliar tissue was oven dried at 70 °C for 12 h, ground, packaged and sent to Harris Laboratories (Lincoln, NE, USA). Tissue concentrations of N, K, S and B were measured. Nitrogen was measured by micro Kjeldahl (a digestive process technique) while the other nutrients were measured using colorimetric inductively coupled plasma techniques.
Soil ion exchange resins were installed immediately post-thinning, but prior to fertilization. The objective of ion exchange resins was to assess soil solution nutrient flux in the fine root zone following treatment. Measurements of soil solution nutrient flux enabled the detection of fertilizer or slash nutrient leaching into the rooting zone of treated trees, thereby providing an empirical validation mechanism for any treatment induced shift in foliar nutrient concentrations. Treatment effect on nutrient flux is indicated by shifts in ion content per surface area of ion exchange resin (µg cm−2) relative to a control. Exchange resins were placed at mineral soil surface, 15, 30 and 45 cm within the crown drip line of two randomly selected trees per fertilizer × thinning treatment to obtain a mean fine rooting zone nutrient flux. Following one growing season, ion exchange resins were exhumed and ions stripped using the shake extraction technique in 60 mL 2N HCl acid. Extract solutions were analyzed for nitrogen (NO3 + NH4), potassium (K), sulfur (S) and boron (B) either through gas diffusion conductivity (N) or inductively coupled plasma emission (K, S, B). Treatment means were calculated as the average ion content across the four soil depths and two replicated treated trees.
All statistical analyses were conducted using Statistix [
29]. Tree growth and resin flow was compared among treatment groups using analysis of variance techniques. Correlation analysis was used to determine the relationship between tree growth and resin flow, regardless of treatment. Foliar chemistry was compared among treatments using analysis of variance techniques and correlation analyses were used to determine if there was a relationship between resin flow and foliar chemistry. Deviation of ion exchange resin nutrient content from the control was assessed using analysis of variance and the least square means test.
3. Results
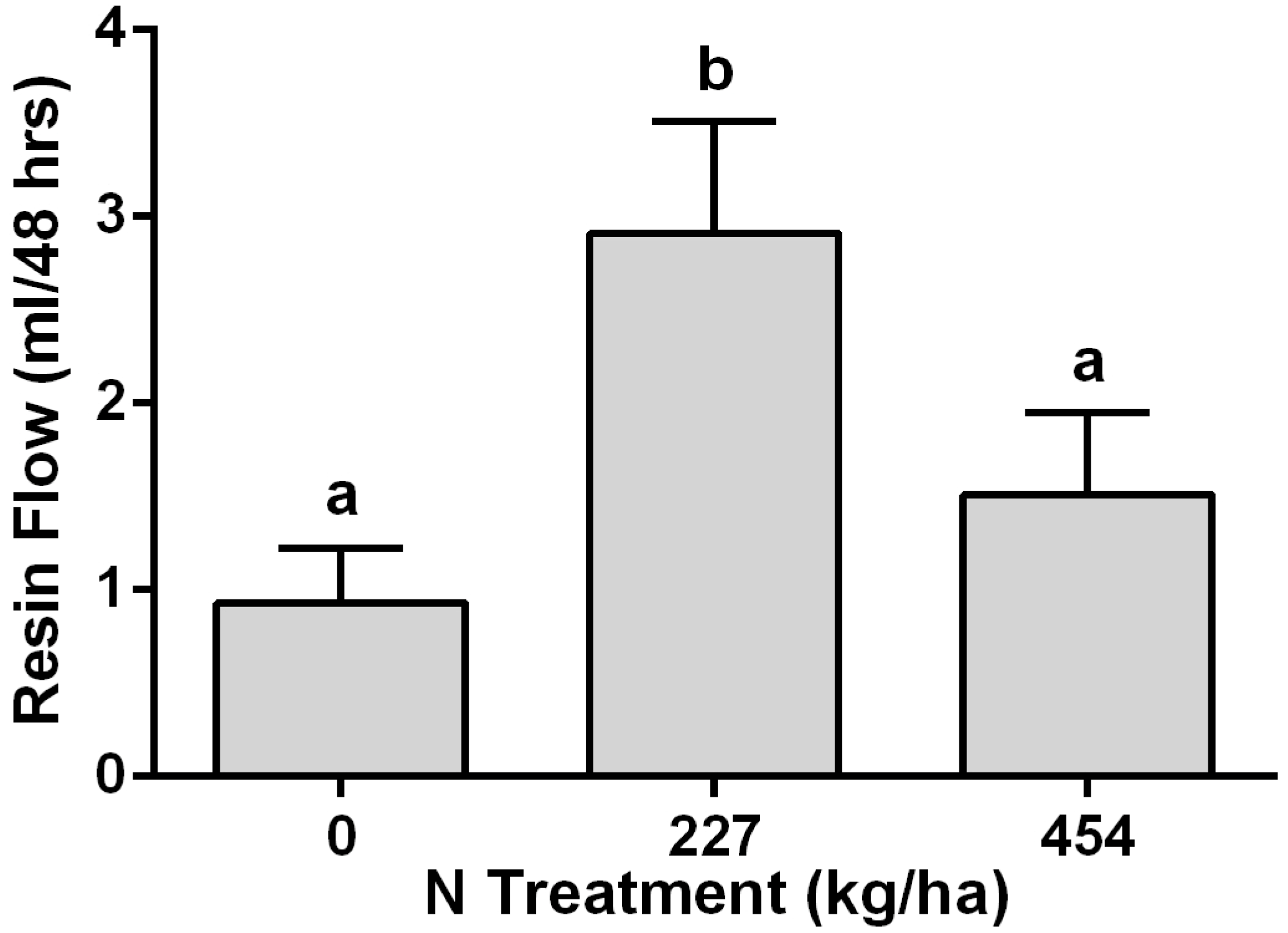
Resin Flow: There was a significant increase in resin flow measurements one year following fertilization in the first experiment (
Figure 1). Resin flow was significantly higher in trees that had received the low-N (227 kg/ha) treatment compared with those that had received no additional N or the high-N (454 kg/ha) treatments (
f = 4.63;
df = 2, 23; (
p >
f) = 0.0205). There was no significant difference in resin flow between the control trees and those that had received the high-N treatments.
Figure 1.
Mean (+SEM) resin flow (mL) during a 48-h period for lodgepole pines that had been individually fertilized with nitrogen applied as urea.
Figure 1.
Mean (+SEM) resin flow (mL) during a 48-h period for lodgepole pines that had been individually fertilized with nitrogen applied as urea.
When fertilization was coupled with a thinning treatment in the second experiment, there were no significant differences in resin flow measurements between trees for fertilized
versus fertilized and thinned one year following treatments (
Table 2): no additional N (
t = 0.58;
df = 7; (
p >
t) = 0.5819), low-N (
t = 0.46;
df = 8; (
p >
t) = 0.6604), high-N (
t = 0.88;
df = 7; (
p >
t) = 0.4105), or the complete fertilization (low-N + K + S + B) (
t = 1.73;
df = 7; (
p >
t) = 0.1280) treatment.
Tree Growth: Tree growth was not measured in the first (no thin) experiment. However, in the second experiment, there were no differences in tree growth one year following fertilization and thinning (
Table 2) in any of the four fertilization categories: no additional N (
t = 0.58;
df = 7; (
p >
t) = 0.5819), low-N (
t = 0.87;
df = 8; (
p >
t) = 0.4071), high-N (
t = 0.16;
df = 7; (
p >
t) = 0.8759) or complete fertilization (low N + K + S + B) (
t = 0.43;
df = 7; (
p >
t) = 0.6787) treatments.
Table 2.
Resin flow (mL/24 h ± SEM) and growth (cm dbh ± SEM) of lodgepole pines the summer following treatment (thinning and fertilization of individual trees with N at low (227 kg/ha), high (454 kg/ha), or a complete blend (low N plus K, B and S).
Table 2.
Resin flow (mL/24 h ± SEM) and growth (cm dbh ± SEM) of lodgepole pines the summer following treatment (thinning and fertilization of individual trees with N at low (227 kg/ha), high (454 kg/ha), or a complete blend (low N plus K, B and S).
| Thin | Resin Flow (mL/24 h ± SEM) | Growth (cm dbh ± SEM) |
|---|
| Control | Low N | High N | Complete | Control | Low N | High N | Complete |
|---|
| Yes | 0.36 ± 0.04 | 0.51 ± 0.69 | 1.06 ± 0.55 | 0.50 ± 0.21 | 0.18 ± 0.13 | 0.36 ± 0.18 | 0.22 ± 0.15 | 0.38 ± 0.12 |
| No | 0.46 ± 0.15 | 0.77 ± 0.44 | 0.55 ± 0.29 | 1.76 ± 0.73 | 0.32 ± 0.15 | 0.17 ± 0.12 | 0.19 ± 0.08 | 0.33 ± 0.05 |
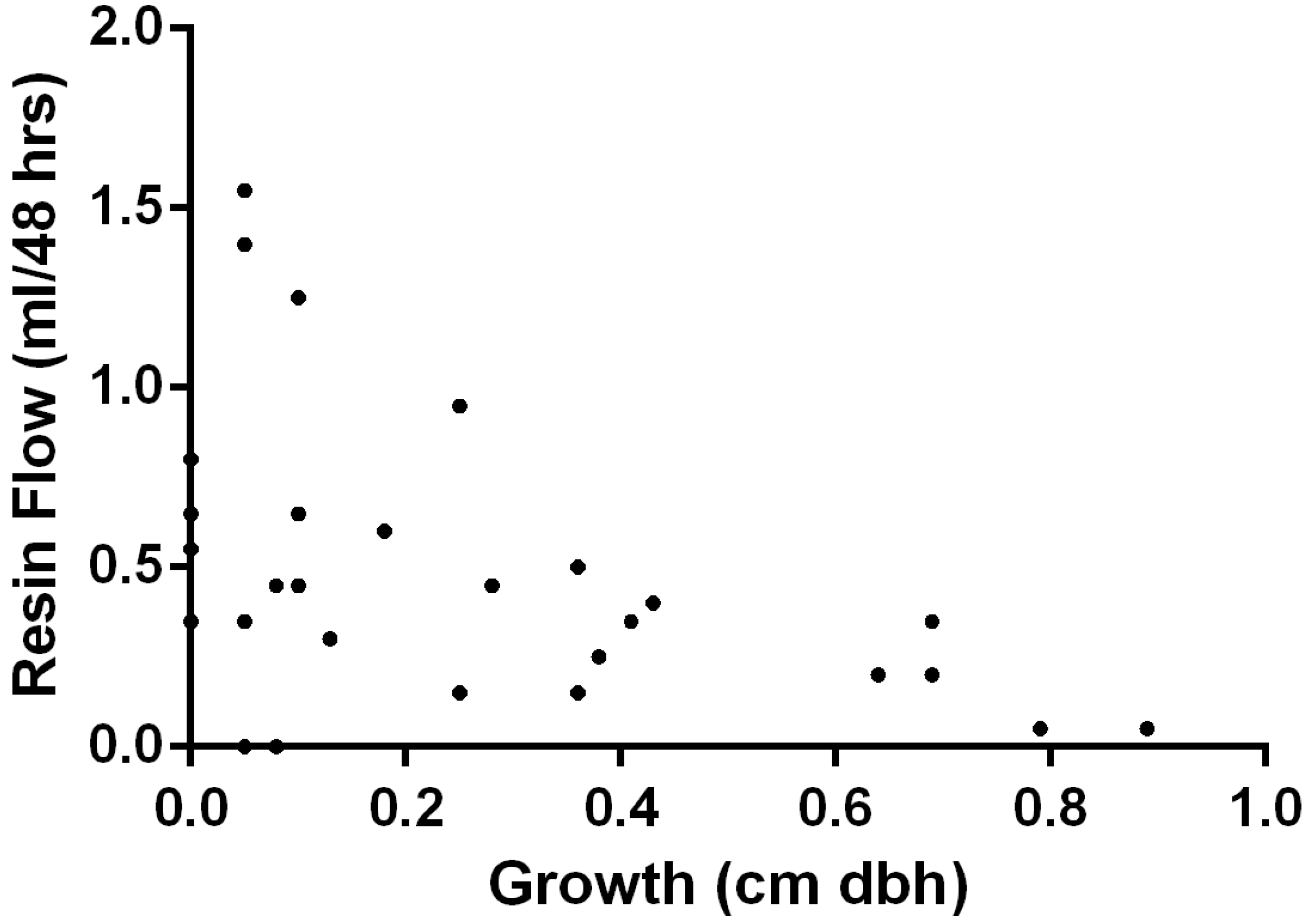
Tree Growth and Resin Flow: The measurements of tree growth and resin flow, regardless of treatment, were combined to determine if there was a relationship between growth and resin flow. When examining all of the trees in the experiment, there was a significant and negative correlation (
k = −0.4461; (
p >
k) = 0.0142) between tree growth and resin flow (
Figure 2).
Figure 2.
Association (k = −0.4461; (p > k) = 0.0142) of resin flow (mL/48 h) and annual growth (cm of diameter at breast height) for lodgepole pines that had received various combinations of fertilization and stand thinning treatments.
Figure 2.
Association (k = −0.4461; (p > k) = 0.0142) of resin flow (mL/48 h) and annual growth (cm of diameter at breast height) for lodgepole pines that had received various combinations of fertilization and stand thinning treatments.
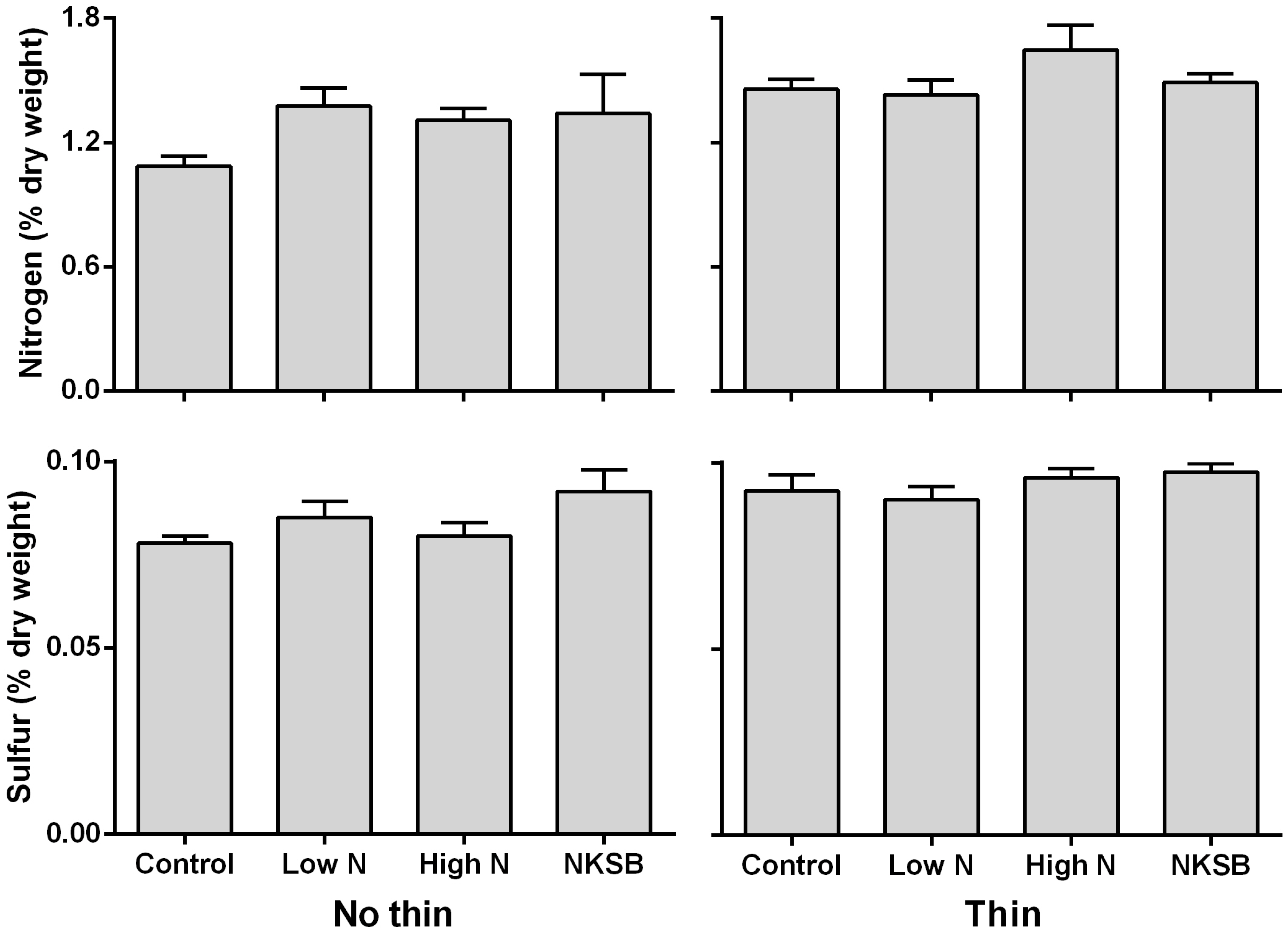
Foliar Chemistry: The N concentration (measured as %age dry weight) in new foliage was significantly influenced by fertilization and thinning (
f = 2.55;
df = 7, 29; (
p >
f) = 0.0354), with the highest measures of foliar N concentration occurring in the thinned treatments, regardless of fertilization (
Figure 3). In addition, the percentage of N in the current year’s foliage was significantly higher in trees that had received the thinning treatment with no fertilization (
t = 5.01;
df = 7; (
p >
t) = 0.0016) or the thinning treatment with the complete fertilizer (
t = 2.37;
df = 7; (
p >
t) = 0.0495) compared with the fertilization treatment without thinning. There were no significant differences in the percentage of N in new foliage of trees in the thinned
versus non-thinned treatments when fertilized with either low (
t = 0.41;
df = 8; (
p >
t) = 0.6946) or high (
t = 0.69;
df = 7; (
p >
t) = 0.5150) levels of N alone. There were no significant differences in K concentration in the current year’s foliage for trees in the thinned
versus non-thinned treatments that had received no additional N (
t = 1.05;
df = 7; (
p >
t) = 0.3276), low-N (
t = 0.80;
df = 8; (
p >
t) = 0.4489), high-N (
t = 0.26;
df = 7; (
p >
t) = 0.8048) or complete fertilization (
t = 1.09;
df = 7; (
p >
t) = 0.3102) treatments.
Figure 3.
Nitrogen and Sulfur content (% ± SEM) of the current year’s foliage following thinning and fertilization (Con = control, Low N = 227 kg N/ha, High N = 454 kg N/ha and Compl = low N + K + S + B) treatments the prior year.
Figure 3.
Nitrogen and Sulfur content (% ± SEM) of the current year’s foliage following thinning and fertilization (Con = control, Low N = 227 kg N/ha, High N = 454 kg N/ha and Compl = low N + K + S + B) treatments the prior year.
Of the two other nutrients examined, foliar S concentration (measured as %age dry weight) was significantly influenced by fertilization and thinning (
f = 3.14;
df = 7, 29; (
p >
f) = 0.0135), with the highest measures of foliar S concentration occurring in the thinned treatments, regardless of fertilization (
Figure 3). As with foliar N concentration, the percentage of S in the current year’s foliage was significantly higher in trees that had received the thinning treatment with no fertilization (
t = 3.04;
df = 7; (
p >
t) = 0.0190) or the thinning treatment with the complete fertilizer (
t = 3.53;
df = 7; (
p >
t) = 0.0096) compared with the fertilization treatment without thinning. There were no significant differences in the percentage of S in the new foliage of trees in the thinned
versus non-thinned treatments when fertilized with either low (
t = 0.80;
df = 8; (
p >
t) = 0.4468) or high (
t = 0.79;
df = 7; (
p >
t) = 0.4557) levels of N alone.
There were no significant differences in the concentration of B in the foliage one year following treatment for trees in the thinned versus non-thinned treatments regardless of fertilization treatment: no additional N (t = 0.77; df = 7; (p > t) = 0.4639), low-N (t = 1.00; df = 8; (p > t) = 0.3488), high-N (t = 0.39; df = 7; (p > t) = 0.7059) or complete fertilization (t = 0.20; df = 7; (p > t) = 0.8485) treatments.
Foliar Chemistry and Resin Flow: There was no significant correlation between resin flow and foliar content of any of the four nutrients examined: N (k = 0.0502; (p > k) = 0.7829), K (k = 0.0764; (p > k) = 0.6758), S (k = 0.0775 (p > k) = 0.6714) or B (k = 0.0584; (p > k) = 0.7492).
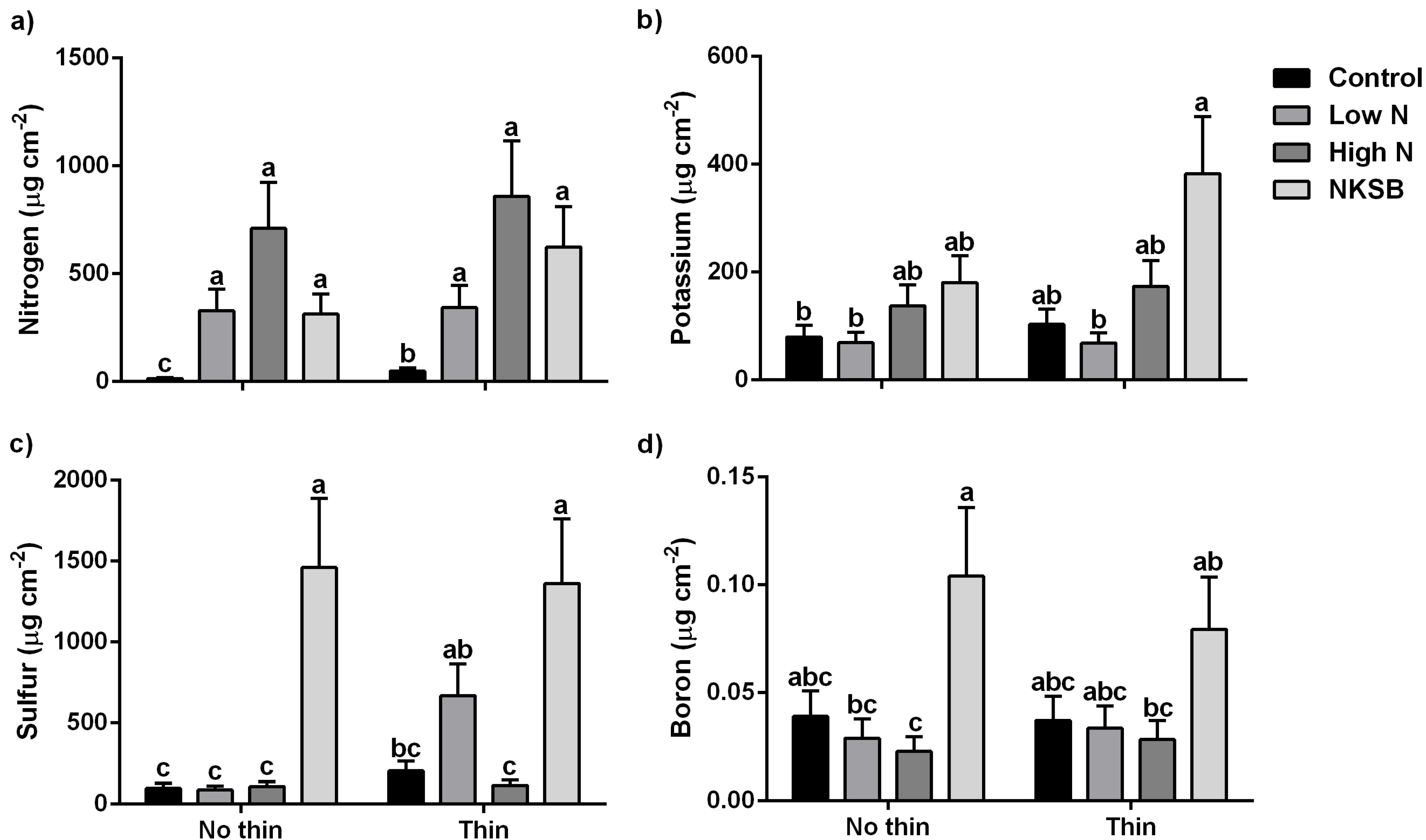
Ion Exchange Resins: Fertilization, and to a lesser degree thinning, significantly increased soil solution N over the control (
f = 12.39;
df = 7, 29; (
p >
f) < 0.0001) (
Figure 4). Due to variation in soil solution N, no significant difference between low-N and high-N with or without thinning were detected. Complete fertilization (including K) with thinning significantly increased soil solution K relative to the no-thin control (
f = 5.06;
df = 7, 29; (
p >
f) < 0.0001). However, there was no significant difference between the remaining fertilizer and thinning treatments. Soil solution S was significantly higher following a complete treatment regardless of thinning treatment (
f = 8.88;
df = 7, 29; (
p >
f) < 0.0001) or after a low-N × thin treatment (
f = 12.39;
df = 7, 29; (
p >
f) < 0.0001). A no-thin complete fertilizer blend showed a significant increase in soil solution B relative to other no-thin fertilizer treatments (
f = 2.75;
df = 7, 29; (
p >
f) = 0.0028); however, B was too variable in solution to show any clear treatment patterns. Overall, one year post-treatment there was no clear thinning effect on soil solution N, K, S or B.
Figure 4.
Soil ion exchange resin nutrient content (µg cm−2) for (a) N; (b) K; (c) S; and (d) B under various combinations of fertilization and stand thinning treatments. Data are presented as log back-transformed means (±SEM) of each nutrient by treatment. Different letters represent significant treatment effect at α = 0.05 level.
Figure 4.
Soil ion exchange resin nutrient content (µg cm−2) for (a) N; (b) K; (c) S; and (d) B under various combinations of fertilization and stand thinning treatments. Data are presented as log back-transformed means (±SEM) of each nutrient by treatment. Different letters represent significant treatment effect at α = 0.05 level.
4. Discussion
Many forest soils in the Intermountain West are deficient in multiple nutrients [
11,
12]. In the current study, thinning resulted in higher concentrations of N and S in the foliage but no differences in foliar K or B one year following treatment. The highest levels of foliar N and S consistently occurred in the trees from thinned treatments, regardless of fertilization. Increased foliar levels of N and S may indicate a greater availability of these nutrients in the absence of competition. While thinning did not show a strong effect on soil nutrient pools, we cannot make the assumption there was no thinning effect. Limitations due to sample size, sampling area and the short duration of the sampling period may be partially responsible. There was a strong thinning effect on foliar N and S concentration. The higher levels of N and S in thinned treatments suggests that either slash nutrient leaching across the rooting area of the treated tree was greater than what was evidenced by the exchange resins, or that increasing foliar levels of N and S indicate greater availability of these nutrients in the absence of competition. Exchange resins and foliar analyses demonstrated a similar lack of treatment effect on K and B. For K, this is not unexpected because K is fairly immobile in soil [
30] meaning any treatment effect may not be detectable until several years after the thinning and fertilization treatments. Also, soil variability was too great to show a B treatment effect, suggesting that the application rate of 3.4 kg B/ha was not high enough to differentiate its effect in soil solution against the native soil solution concentration. However, volcanic ash anion adsorption mechanisms may additionally contribute to this lack of treatment differentiation. Forested soils influenced by weathered fine textured volcanic ash have been demonstrated to absorb >60% of applied B from soil solution [
31].
We did not measure inner bark N content during the current study, but prior work has demonstrated that N fertilization results in higher concentrations of N in the inner bark of lodgepole pine [
13], changes in foliar chemistry and increased growth [
14,
15]. However, the increased growth rate did not prevent attack by mountain pine beetle in the fertilized stands [
14]. In addition, higher N content within a host pine can result in higher N content of bark beetles reared in those trees [
13,
16], the production of larger adults [
16] that can result in beetle populations having increased progeny production and adult emergence [
17].
Resin flow represents the first step of tree resistance to attack by bole-invading bark beetles and their associated fungi [
7] and a rapid flow can prevent the initial attacking beetles from entering a tree and establishing an attraction site from which aggregation pheromones can be released [
32]. Further, the concentration of resin ducts in recent growth has been demonstrated to affect the ability of bark beetles to successfully colonize host trees [
10]. Therefore, tree or stand-level treatments that result in an increase in resin flow should also result in an increase in tree resistance to bark beetle attack. In our first test, a low-N fertilization treatment, without stand thinning, increased resin flow over the non-fertilized trees or trees that received a high-N fertilization treatment. During our second test, in the absence of a thinning treatment, it was again the trees that received a low-N fertilization treatment (whether N alone or in combination with K, S and B) that had the highest resin flows. However, there was not a significant increase in resin flow when comparing trees within any individual fertilization treatment between thinned and non-thinned areas. Our measurements occurred one year following treatment and therefore, the positive effects of fertilization may be masked when fertilization and thinning treatments are combined by factors such as differences in annual climate (temperature and/or precipitation). Further, there was no relationship between foliar levels of N, K, S or B and resin flow.
In the current study, we did not observe an increase in growth for any of the fertilization treatments when compared with their respective fertilization plus thinning groups. However, growth was measured one year following treatment application which may not have allowed enough time for differences to be discernable. We chose a one-year sample period because silvicultural treatments used to reduce mountain pine beetle colonization of host trees are not always applied multiple years in advance of beetle attack.
A scenario of increased growth corresponding to decreased plant resistance agrees with several theories detailing the interaction between plants and their herbivorous insects. Several of these theories assume that defenses against herbivores are costly to a plant because they divert resources away from growth [
18]. For example, carbon-nutrient balance theory hypothesizes that nutrient availability to a plant is a major factor in limiting plant growth [
21] and that low availability of nutrient resources should favor the production and maintenance of substantive constitutive defenses [
22,
23,
24]. Tree resistance can be related to growth but under growth-limiting field conditions, individual components of the constitutive resistance may be mutually exclusive in their expression [
33]. Resin flow is primarily a physical parameter of tree constitutive resistance parameters, but the resin does contain toxic, C-based components secondary metabolites (monoterpenes). Prior studies have demonstrated a decrease in C-based secondary metabolites following fertilization [
25,
26] and loblolly pine has its lowest resin flow during periods of rapid tree growth [
27]. Our results suggest that a similar reduction in resin flow accompanies increased growth in lodgepole pine. Similarly, the fundamental principle of the growth-differentiation balance hypothesis is the existence of a physiological tradeoff between growth and differentiation processes (including the maintenance of resistance/defense chemicals and mechanisms) [
17]. While growth has been associated with conifer resistance to bark beetle attack [
34] we measured a reduction in resin production associated with increased growth one year following thinning and fertilization.
Large outbreaks of bark beetles, including mountain pine beetle in lodgepole pine [
35], have been recorded. However, the magnitude of bark beetle outbreaks has increased in recent years [
3]. Some species (including mountain pine beetle) are causing increasing mortality in novel hosts or known hosts that occur in less suitable environments [
35,
36,
37]. The increasing magnitude and extent of bark beetle outbreaks has been placed into a context of thresholds and feedback processes that included scales at the tree and forest stand levels [
3]. Beetles must overcome a threshold of tree resistance parameters (including resin flow) to succeed in colonizing a tree and for reproduction to be successful [
38,
39]. Within this context, increasing available nutrient levels to trees through stand manipulations may result in modified resistance parameters that must be considered when making management decisions.



{kind=link}
{kind=link}
{kind=link}
{kind=link}



