1. Introduction
Northern white-cedar (
Thuja occidentalis L.) is a species of high commercial and ecological value that is native to North America. The current annual wood harvest for northern white-cedar in Ontario, Quebec, and New England ranges from 275,000 to 350,000 m
3 [
1]. Northern white-cedar is also an important habitat for many bird and mammal species [
1], especially as an important winter browse and protective cover species for white-tailed deer (
Odocoileus virginianus Zimmerman) [
2]. Since the middle of the 19th century, a decline in northern white-cedar populations has been noted over most of its natural range [
3,
4]. This decline has been linked to an increase in white-tailed deer populations [
2,
3], but could also be related to management practices.
Natural northern white-cedar regeneration can be established through a variety of approaches; partial cutting is especially effective [
1,
5]. However, its development may be hampered by browsing pressure, since northern white-cedar is highly palatable. Herbivore preferences can limit northern white-cedar regeneration and, at the same time, favor less palatable species [
6]. Browsing damage is mostly caused by white-tailed deer, but also by moose (
Alces alces Gray) and snowshoe hare (
Lepus americanus Erxleben) [
7,
8].
Northern white-cedar forms monocultures on wet and dry sites, but grows best and produces higher quality logs on mesic mineral soils, where it coexists with many other gymnosperms and deciduous angiosperms [
1]. Even though much of the high-quality northern white-cedar resource is found in mixedwood mesic sites, most of the available research has been conducted in monocultures growing on wet sites [
1,
9]. Light partial cutting strategies, such as selection cuts, have been suggested as a means to manage northern white-cedar [
1,
5]. However, tree growth response to partial cuts is influenced by the proportion of basal area in conifer species and the cedar response decreases with increasing softwood proportion [
10]. Such practices have been shown to be effective in establishing northern white-cedar and increasing seedling establishment [
5]. However, northern white-cedar in mixedwood stands has to compete with a variety of species, many which present faster height growth. Management practices in mixedwood stands usually target more abundant or valuable species, such as yellow birch (
Betula alleghaniensis Britt). Since yellow birch requires more light, partial cutting aimed at regenerating this species may lead to too severe cuts for northern white-cedar. In such cases, cedar is likely to be outperformed. Even when low-intensity cuts leads to proper northern white-cedar seedling establishment, differential browsing in mixedwood stands may hinder regeneration response of northern white-cedar to partial cutting. Since natural northern white-cedar regeneration is strongly dependent on understory establishment, it becomes necessary to examine how partial harvesting in mixedwood stands affects seedling and sapling abundance and growth. Few studies have analyzed the growth response of pre-established northern white-cedar seedlings and saplings, which may play a crucial role in stand renewal [
11,
12]. No study is available for mixedwood stands.
The main objective of this retrospective study was to quantify the height distribution and the height and diameter growth increments of natural northern white-cedar regeneration in mixedwood stands, in the presence or absence of deer browsing. We measured the abundance of northern white-cedar by height class, and modelled the effects of time and canopy light transmittance on the height increment of individual northern white-cedar stems that were 0 to 200 cm tall. In addition, we determined the diameter growth response of pre-established regeneration (≥3 years old) to partial harvesting, over a 10-year period. Results are discussed regarding sustainable mixedwood stand under different contexts of browsing pressure.
2. Methods
2.1. Study Site
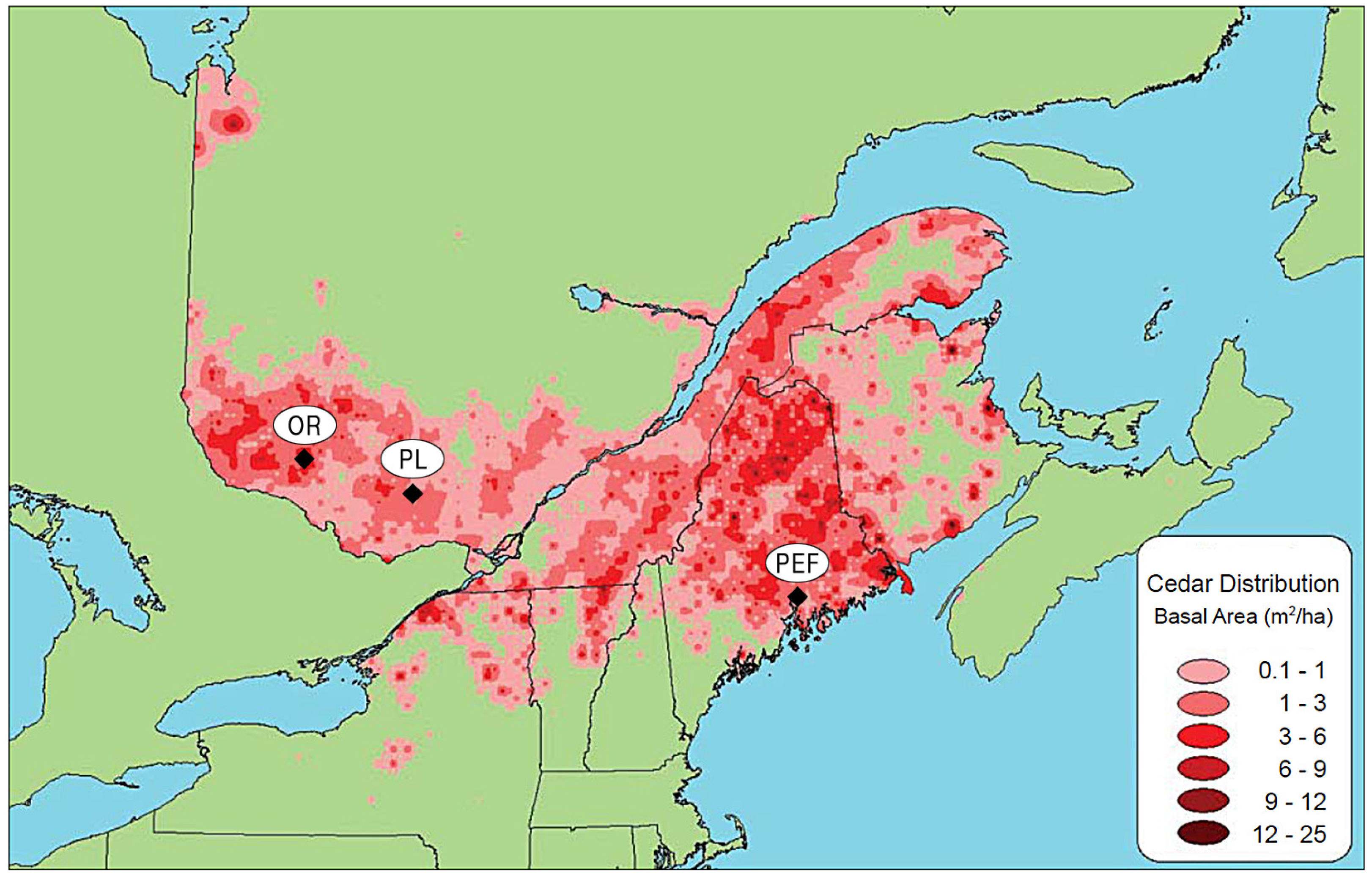
The study encompassed a wide area, which included a significant portion of the geographic range of northern white-cedar and a variety of deer densities (
Figure 1;
Table 1). Moose and hare were also present. Herbivore densities were determined on the basis of hunting surveys rather than direct inventories [
13]. Consequently, they were interpreted only in a descriptive fashion because they surveyed only a part of the entire population. Three sites were selected: in the first, located in the Outaouais region of western Quebec, Canada (hereafter called “OR”), no deer or evidence of browsing were observed on northern white-cedar regeneration. In the second, in the Papineau-Labelle wildlife reserve of western Quebec, Canada (hereafter called “PL”), deer density was high and browsing was frequently observed on northern white-cedar regeneration. The third site, on the Penobscot Experimental Forest in Maine, USA (hereafter called “PEF”), had the highest deer density; browsing was observed on 90% of large northern white-cedar seedlings and small saplings [
6].
Figure 1.
Location of the 3 study sites showing the northern white-cedar basal area in Quebec, Atlantic provinces, and northeastern United-States: Outaouais Region (OR), Papineau-Labelle wildlife reserve (PL), Penobscot Experimental Forest (PEF). Reproduced with permission from Éric Forget, Nova Sylva [
1].
Figure 1.
Location of the 3 study sites showing the northern white-cedar basal area in Quebec, Atlantic provinces, and northeastern United-States: Outaouais Region (OR), Papineau-Labelle wildlife reserve (PL), Penobscot Experimental Forest (PEF). Reproduced with permission from Éric Forget, Nova Sylva [
1].
Table 1.
Ecological characteristics of the 3 study sites: Outaouais Region (OR site), Papineau-Labelle wildlife reserve (PL site), and Penobscot Experimental Forest (PEF site).
Table 1.
Ecological characteristics of the 3 study sites: Outaouais Region (OR site), Papineau-Labelle wildlife reserve (PL site), and Penobscot Experimental Forest (PEF site).
| Parameter | OR site | PL site | PEF site |
|---|
| Geographical coordinates | 46°34′ N, 77°27′ W | 46°10′ N, 75°20′ W | 44°52′ N, 68°38′ W |
| Herbivore density per km2 a |
| Deer | Absent (0.0) | High (5.5) | High (7.0–8.0) |
| Hare | Low (0.6) | Low (0.6) | High (15.2) |
| Moose | Low (0.3) | Low (0.1) | Low (0.3) |
| Climate b |
| Bioclimatic domain or forest region | Balsam fir- Yellow birch | Sugar maple- Yellow birch | Acadian Forest |
| Elevation (m) | 400 | 320 | 75 |
| Mean annual temperature (°C) | 2.55 ± 0.42 | 2.83 ± 0.45 | 6.68 ± 0.34 |
| Total annual precipitation (mm) | 1037 ± 115 | 1138 ± 115 | 1066 ± 137 |
| Mean annual snowfall (cm) | 336 ± 63 | 357 ± 72 | 289 ± 78 |
| Growing season (days) | 147 ± 16 | 157 ± 17 | 183 ± 15 |
| Soil |
| Texture of B-horizon | Sandy loam | Sandy loam | Clay loam |
| Sand (%) | 70.2 | 61.8 | 35.6 |
| Silt (%) | 21.4 | 27.2 | 37.2 |
| Clay (%) | 8.4 | 11.0 | 27.2 |
| pH Humus | 3.25 ± 0.47 | 3.15 ± 0.46 | 3.28 ± 0.31 |
| pH B-horizon | 3.95 ± 0.29 | 3.95 ± 0.49 | 4.45 ± 0.53 |
| Organic layer (cm) | 9 | 7 | 9 |
| Soil depth(cm) | 50–100 | 50–100 | 50–100 |
| Drainage | Good | Good | Good to poor |
During the summer of 2005, 16 mixedwood stands were selected: 4 on the OR site, 4 on the PL site, and 8 on the PEF site. Stand selection guidelines were: (i) management by partial cutting regime; (ii) stand composition representative of stands growing on mesic sites; and (iii) mean proportion of merchantable basal area of northern white-cedar of 8% to 66% (diameter at breast height (DBH) ≥ 9.1 cm;
Table 2). On the OR and PL sites, diameter-limit cuts had been conducted between 1971 and 1983. The PEF had been established by the United States Forest Service in 1950 for stand-level silvicultural experiments [
17]. On the PEF, we used 2 stands for each partial cutting regime, which included selection cuttings with 5-, 10-, and 20-year cutting cycles, and fixed diameter-limit cutting with a harvest interval of about 20 years, based on stand volume increment [
17]. Fixed diameter-limit cutting was a partial harvesting according to species-specific size thresholds, above which merchantable trees were cut [
18]. Even if frequency and intensity of partial harvesting differed among sites, all treatments could potentially create conditions suitable for successful regeneration and recruitment of northern white-cedar [
1,
5,
12]. In all of these stands, management did not focus on northern white-cedar; rather, the main target species was yellow birch in the PL and OR sites, and red spruce (
Picea rubens Sarg.) for the PEF [
19].
Table 2.
Composition of the 3 study sites: Outaouais Region (OR site), Papineau-Labelle wildlife reserve (PL site), and Penobscot Experimental Forest (PEF site).
Table 2.
Composition of the 3 study sites: Outaouais Region (OR site), Papineau-Labelle wildlife reserve (PL site), and Penobscot Experimental Forest (PEF site).
| Parameter | OR site | PL site | PEF site |
|---|
| Mean basal area (m2/ha) | 34.5 | 37.2 | 23.5 |
| Mean density of seedlings (seedlings/ha) | 50,139 | 49,491 | 30,189 |
| Last partial harvesting | 1971, 1974 | 1979, 1981, 1983 | 1994, 1995, 1997, 1998, 1999, 2002 |
| Composition of the overstory (%) a | Northern white-cedar | 30 | 29 | 8 |
| Balsam fir | 13 | 16 | 22 |
| Red/white spruce | 4 | 10 | 23 |
| Hemlock | 0 | 12 | 31 |
| Yellow birch | 23 | 15 | 0 |
| Red maple | 11 | 6 | 9 |
| Sugar maple | 3 | 6 | 0 |
| Paper birch | 0 | 2 | 3 |
| Composition of the understory (%) b | Northern white-cedar | 17 | 7 | 12 |
| Balsam fir | 16 | 31 | 43 |
| Red/white spruce | 2 | 2 | 9 |
| Hemlock | 0 | 3 | 20 |
| Yellow birch | 20 | 20 | 0 |
| Red maple | 16 | 17 | 8 |
| Sugar maple | 28 | 17 | 1 |
| Paper birch | 1 | 1 | 8 |
The PEF is warmer and has a longer growing season than the 2 Quebec sites, while the OR site is the coldest (
Table 1). On all 3 sites, soil texture ranges from clay loam to sandy loam, with good to poor drainage. At the time of the survey, mean stand basal area ranged from 23.5 to 37.2 m
2/ha (
Table 2). Dominant canopy species included mixed northern conifers (northern white-cedar, balsam fir (
Abies balsamea (L.) Mill.), white spruce (
Picea glauca (Moench) Voss), and red spruce); hemlock (
Tsuga canadensis (L.) Carrière) was present only on the PL and PEF sites. The most common hardwoods were yellow birch, paper birch (
Betula papyrifera Marsh.), red maple (
Acer rubrum L.), and sugar maple (
Acer saccharum Marsh.). Spatial position and spacing of trees (horizontal structure) were heterogeneous, with an intimate mix of species and aggregations of northern white-cedar. Vertical structure was multicohort, as a result of past harvestings and natural disturbances. No major natural disturbance was reported, except for the 2 most recent spruce budworm (
Choristoneura fumiferana (Clem.)) outbreaks [
10,
20]. The understory was mostly composed of balsam fir, maples, northern white-cedar, yellow birch, and hemlock (except on the OR site). Mean shrub cover was 41%–60%, mainly composed of mountain maple (
Acer spicatum Lamb.) and squashberry viburnum (
Viburnum edule (Michx.) Raf.).
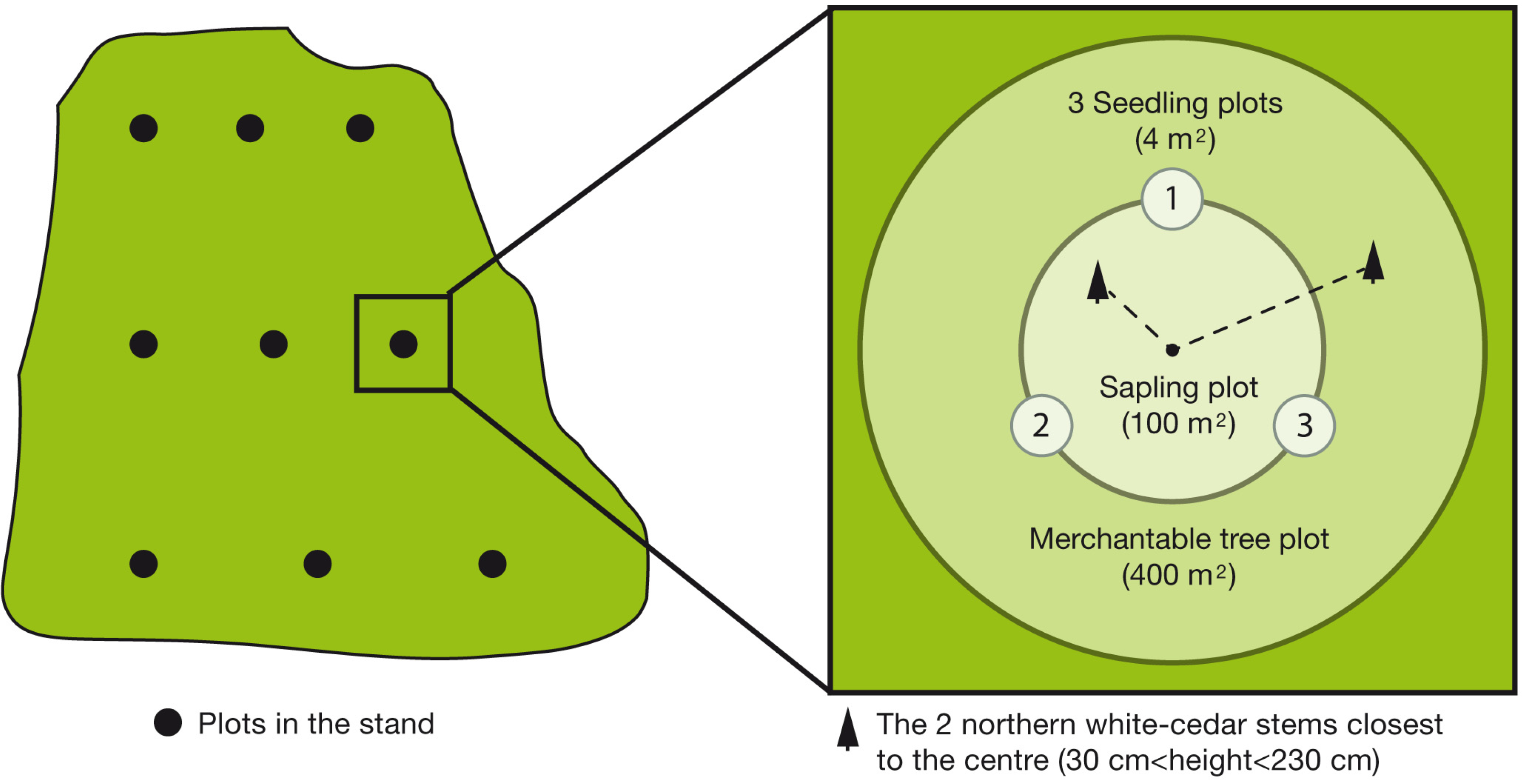
2.2. Design and Data Collection
On the OR and PL sites, we established 9 plots per stand consisting of 2 concentric circles measuring 400 m
2 and 100 m
2 (
n = 72 plots (36 plots per site)) to inventory merchantable trees and saplings by species and diameter class (
Figure 2). For details, see Ruel
et al. [
10]. Regeneration monitoring of northern white-cedar by 5-cm height classes was done in 3 equally spaced, circular 4-m
2 plots on the circumference of the sapling plots (
n = 216 seedling plots). For the PEF, the same approach was used, with 3 circular 4-m
2 seedling plots established on the circumference of each of the 45 sapling plots measuring 100-m
2 (
n = 135 seedling plots). Plots were established on a systematic grid according to compartment area (see Kenefic
et al. [
19] et Larouche
et al. [
6]). The same 5-cm height classes were used to monitor regeneration.
Figure 2.
Schematic representation of experimental design used to inventory merchantable trees, saplings, and seedlings on the 3 study sites, and selection of northern white-cedar stems on the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site).
Figure 2.
Schematic representation of experimental design used to inventory merchantable trees, saplings, and seedlings on the 3 study sites, and selection of northern white-cedar stems on the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site).
The 2 northern white-cedar stems closest to the center of each sapling plot were selected and numbered on the OR and PL sites (n = 144 stems). On the OR site, they were 130 to 230 cm tall. On the PL site, very few stems taller than 130 cm were available, so we had to lower the height criterion from 130 to 30 cm. We did not find enough seedlings and small saplings taller than 30 cm on the PEF to conduct stem analyses for this site.
We marked ground level and the north side on each stem, and assessed browsing on each tree by visual estimation of the percentage of total foliage consumed into classes (unbrowsed; 1%–25%; 26%–50%; 51%–75%; 76%–100%). After measuring ground-level diameter (GLD) and total height, we excavated the roots of each selected stem to bring them in laboratory for stem analyses.
In order to characterize the microenvironment, we estimated the canopy transmittance from a point 150 cm above the soil level after stem harvesting, on cloudy days in July 2005, using hemispherical digital photographs of the forest canopy. Canopy transmittance is the percentage of light transmitted through canopy trees in the openings. Detailed methods for canopy transmittance measurements are found in Hébert
et al. [
21].
2.3. Stem Analyses
We sectioned harvested stems and sampled cross-sectional discs at 10-cm intervals from ground level to top height. Discs were dried and sanded, and rings were counted with a microscope. If the difference between 2 successive discs was more than 10 years, we counted rings on supplementary cross-sectional discs taken from the middle of the section. We counted 25% of the discs twice, and obtained a mean difference of 0.61 year between both counts. For advance regeneration (≥3 years old at the time of partial harvesting; n = 58 stems), discs collected at ground level were analyzed in detail with a tree-ring measuring system (TA Uni-slide System, Velmex Inc., Bloomfield, NY, USA; precision = ± 0.1 mm) to separately measure growth for 4 perpendicular radii corresponding to the cardinal points. For 2 discs where butt rot was present, only 2 radii were used to measure growth. Annual growth was averaged for each disc and multiplied by 2 to estimate diameter increment. Since the 2005 growing season (when samples were taken) was incomplete, it was excluded from the diameter growth analysis.
2.4. Statistical Analyses
Regarding regeneration abundance variable, it was difficult to fit a good model using the Poisson or the negative binomial distributions because of the excess of zero counts. Even in categorizing the response variable, we couldn’t perform the Fisher’s exact test, and the Chi-Square test was not valid since a lot of cells had less than 5 expected counts. Hence, we only present descriptive statistics for regeneration density by height class.
A model was constructed to determine the time needed to reach a given height. Age 0 was set at ground level. The history of browsing pressure for each stem was unknown and, thus, could not be included in a model using retrospective data. Analysis was performed using the NLMIXED procedure (SAS 9.1, SAS Institute Inc., Cary, NC, USA) with height of the disc as the independent variable, and age of the disc as the dependent variable. The analysis treated sites, stands, and plots as random effects, and took into account multiple measurements (3 to 20 discs per stem) using another random effect. Only significant variables were included in the final model. Assumptions of normality, homoscedasticity, and independence were met. A power function (Equation 1) was used, and adjusted correlation coefficients were computed using a regression (PROC REG; SAS 9.1) of predicted values as a function of observed values.
where
T = age of the disc (years);
H = height of the disc (cm);
a and
b are parameters to be estimated.
Mean height increment for the 3 last complete growing seasons (2002–2004) was also modelled as a function of canopy transmittance using the GLM procedure (SAS 9.1). Assumptions of normality, variance homogeneity, and independence were met. The following quadratic model form was used (Equation 2).
where
HI = mean height increment of the 3 last complete growing seasons (cm);
CT = canopy transmittance (%);
a,
b, and
c are parameters to be estimated.
GLD response to partial harvesting was evaluated on individual stems to determine if the pre-established regeneration responded to the canopy opening. We identified the moment at which growth became significantly greater than before treatment, and how long this response persisted over a 10-year period. We analyzed 3-year periods instead of single years, to filter out the variability in diameter growth due to annual changes in temperature and precipitation [
22,
23]. Because the exact month of the partial harvest was unknown, we did not include the year of harvesting (year = 0) in the diameter growth analysis. After harvesting (years = 1 to 10), we used the moving average method to find differences generated by adding and dropping only one year at a time [
24]. This method is frequently used in dendrochronology to search for the moment of the release of stems following disturbance of the canopy [
23,
25]. We constructed models for each individual seedling and post-harvesting period to compare the 8 post-harvesting periods with the pre-harvesting period (mean diameter increment of years −3 to −1). We used the MIXED procedure (SAS 9.1) to build a model which included random effects (site, stand, and plot) and the fixed effect of total stem height before harvesting. Assumptions of normality, independence of residuals, and variance homogeneity were met. A significant difference between estimates (α = 0.05) indicates a positive or negative difference compared to the pre-harvesting period.
3. Results
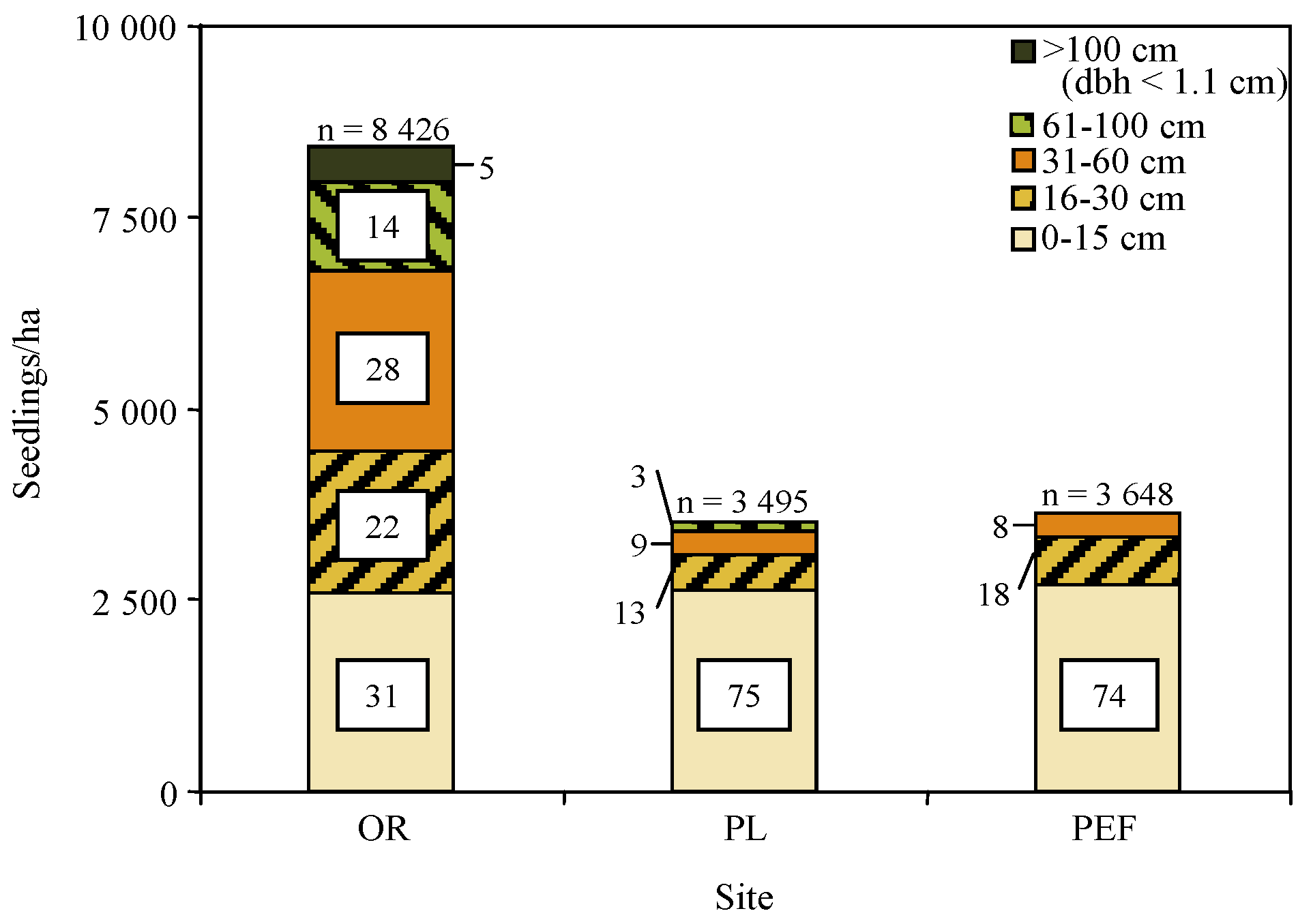
Visual analysis of
Figure 3 shows that the abundance of northern white-cedar regeneration on the OR site was twice as high as on the PL and PEF sites. On the OR site, seedlings in all height classes were present, including 5% of seedlings >100 cm. The density of seedlings smaller than 15 cm was the same on all sites, but the PL and PEF sites had fewer seedlings in other height classes: in these two sites, approximately 75% of seedlings were ≤15 cm tall, and seedlings >100 cm were totally absent. Some seedlings >30 cm were found at the PL and PEF sites, but no seedlings >60 cm were found at PEF.
Figure 3.
Height distribution of northern white-cedar regeneration on three sites selected throughout the native range of northern white-cedar: Outaouais Region (OR), with no deer or evidence of browsing, Papineau-Labelle wildlife reserve (PL), with high deer density and browsing, and Penobscot Experimental Forest (PEF), with the highest deer density and most intense browsing. Numbers indicate the percentage of seedlings by height class; n = total number of seedlings measured.
Figure 3.
Height distribution of northern white-cedar regeneration on three sites selected throughout the native range of northern white-cedar: Outaouais Region (OR), with no deer or evidence of browsing, Papineau-Labelle wildlife reserve (PL), with high deer density and browsing, and Penobscot Experimental Forest (PEF), with the highest deer density and most intense browsing. Numbers indicate the percentage of seedlings by height class; n = total number of seedlings measured.
Of the 144 stems from the OR and PL sites used in our analyses (
Table 3), only the 58 stems older than three years at ground level at the time of harvest were considered pre-established. The fact that 86 sampled stems were not present before partial harvesting reveals that establishment of new northern white-cedar regeneration occurred following partial harvesting. In 2005, 28.5% of northern white-cedar harvested stems from the OR and PL sites presented browsing marks.
Table 3.
Descriptive characteristics of northern white-cedar harvested stems on the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site) (a) just before the last partial cutting that occurred between 1971 and 1983; and (b) in 2005, when they were harvested during the current study.
Table 3.
Descriptive characteristics of northern white-cedar harvested stems on the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site) (a) just before the last partial cutting that occurred between 1971 and 1983; and (b) in 2005, when they were harvested during the current study.
| (a) Before partial cutting (pre-established stems ≥3 years old) |
| Parameter | N a | Mean | ± | SD b | Minimum | Maximum |
| Number of stems | 58. | | | | | |
| Height (cm) | | 23.4 | ± | 20.3 | 2.5 | 79.6 |
| GLD (cm) c | | 3.0 | ± | 2.2 | 0.3 | 10.0 |
| Age (years) c | | 12.2 | ± | 7.1 | 3.0 | 34.0 |
| (b) In 2005 (all harvested stems) |
| Parameter | N a | Mean | ± | SD b | Minimum | Maximum |
| Number of stems | 144 | | | | | |
| Height (cm) | | 127.2 | ± | 62.7 | 34.2 | 281.0 |
| GLD (cm) c | | 18.9 | ± | 12.7 | 3.1 | 47.6 |
| Age (years) c | | 28.6 | ± | 14.2 | 8.0 | 65.0 |
| Browsing d |
| Unbrowsed | 103 | | | | | |
| 1%–25% | 26 | | | | | |
| 26%–50% | 9 | | | | | |
| 51%–75% | 5 | | | | | |
| 76%–100% | 1 | | | | | |
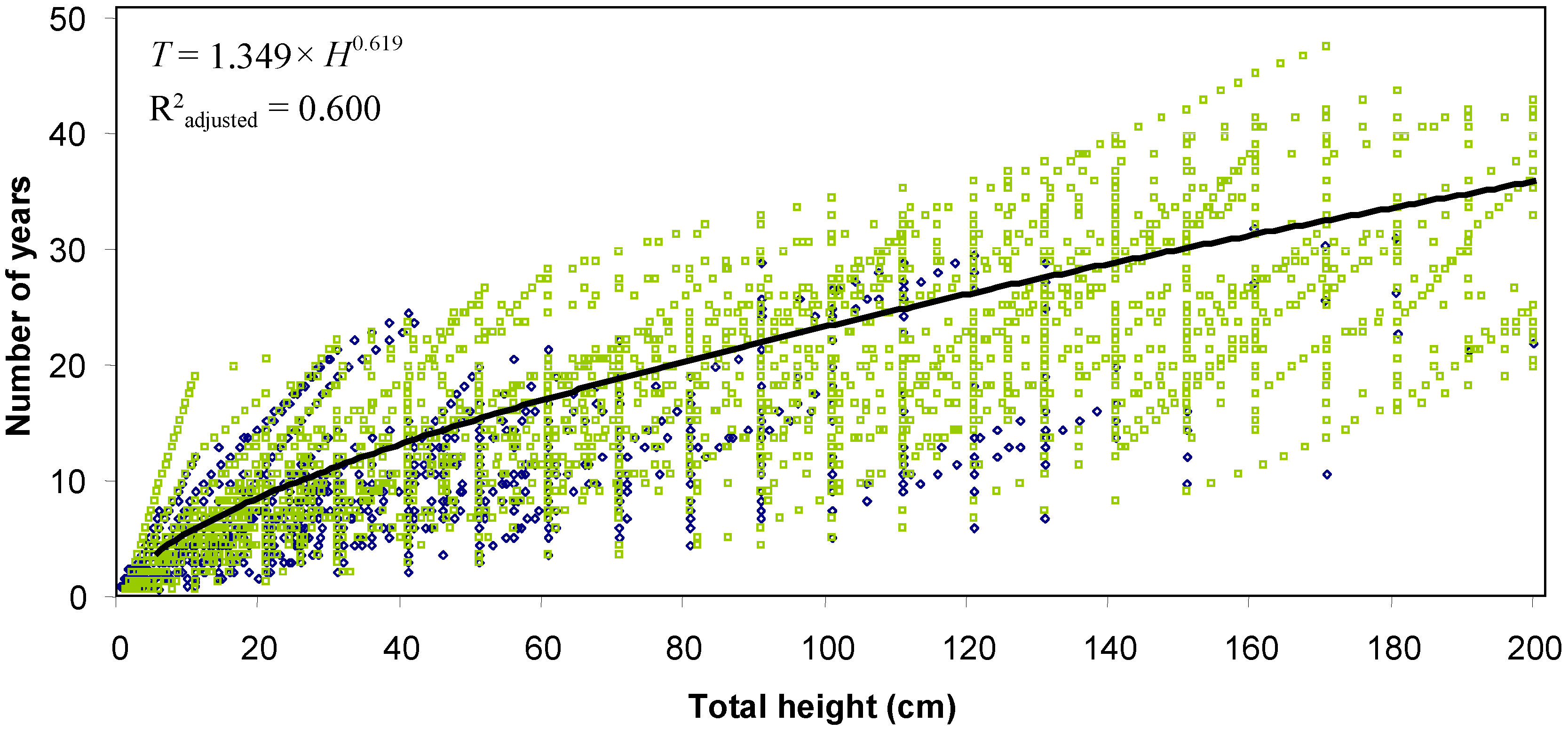
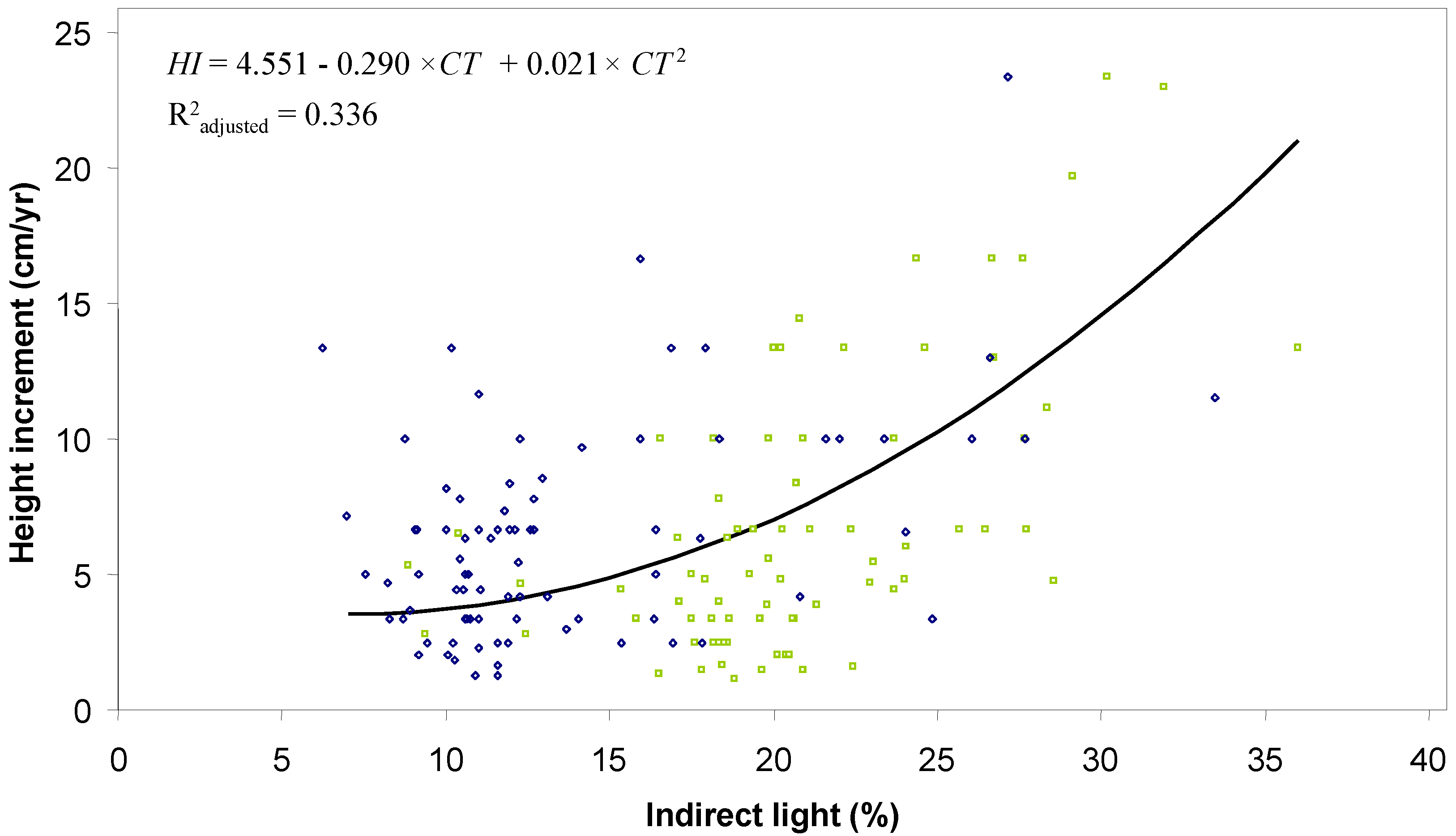
The age-height relationship shows that northern white-cedar stems need 11 years to reach a height of 30 cm, 28 years to reach 130 cm, and 36 years to reach 2 m (
Table 4;
Figure 4). The mean height increment of the last three complete growing seasons was influenced by canopy transmittance (
Table 4;
Figure 5). The model is applicable only for canopy transmittance values ranging from 7% to 36%. When transmittance values increase from 10% to 20%, height increment rises by a total of 3.3 cm/yr. By contrast, when transmittance values increase from 20% to 30%, height increment doubles to reach a total of 7.5 cm/yr. Total height and browsing pressure in 2005 did not influence the relationship between height increment and canopy transmittance.
Table 4.
Estimates and significance tests for parameters of Equations (1) and (2).
Table 4.
Estimates and significance tests for parameters of Equations (1) and (2).
| Equation 1 | Equation 2 |
|---|
| Parameter | Estimate | SE a | DF b | t value | Parameter | Estimate | SE a | DF b | t value |
|---|
| a | 1.349 | 0.056 | 149 | 24.01 | a | 4.551 | 3.907 | 1 | 1.16 |
| b | 0.619 | 0.006 | 149 | 96.26 | b | −0.290 | 0.372 | 132 | −0.78 |
| | | | | | c | 0.021 | 0.009 | 132 | 2.37 |
Figure 4.
Relationship between age (T) and height (H) calculated from Equation (1) for northern white-cedar harvested stems of the Outaouais Region (OR site; green squares) and Papineau-Labelle wildlife reserve (PL site; blue diamonds); n = 144 stems.
Figure 4.
Relationship between age (T) and height (H) calculated from Equation (1) for northern white-cedar harvested stems of the Outaouais Region (OR site; green squares) and Papineau-Labelle wildlife reserve (PL site; blue diamonds); n = 144 stems.
Figure 5.
Relationship between height increment (HI) and canopy transmittance (CT) calculated from Equation (2) for northern white-cedar harvested stems of the Outaouais Region (OR site; green squares) and Papineau-Labelle wildlife reserve (PL site; blue diamonds); n = 144 stems.
Figure 5.
Relationship between height increment (HI) and canopy transmittance (CT) calculated from Equation (2) for northern white-cedar harvested stems of the Outaouais Region (OR site; green squares) and Papineau-Labelle wildlife reserve (PL site; blue diamonds); n = 144 stems.
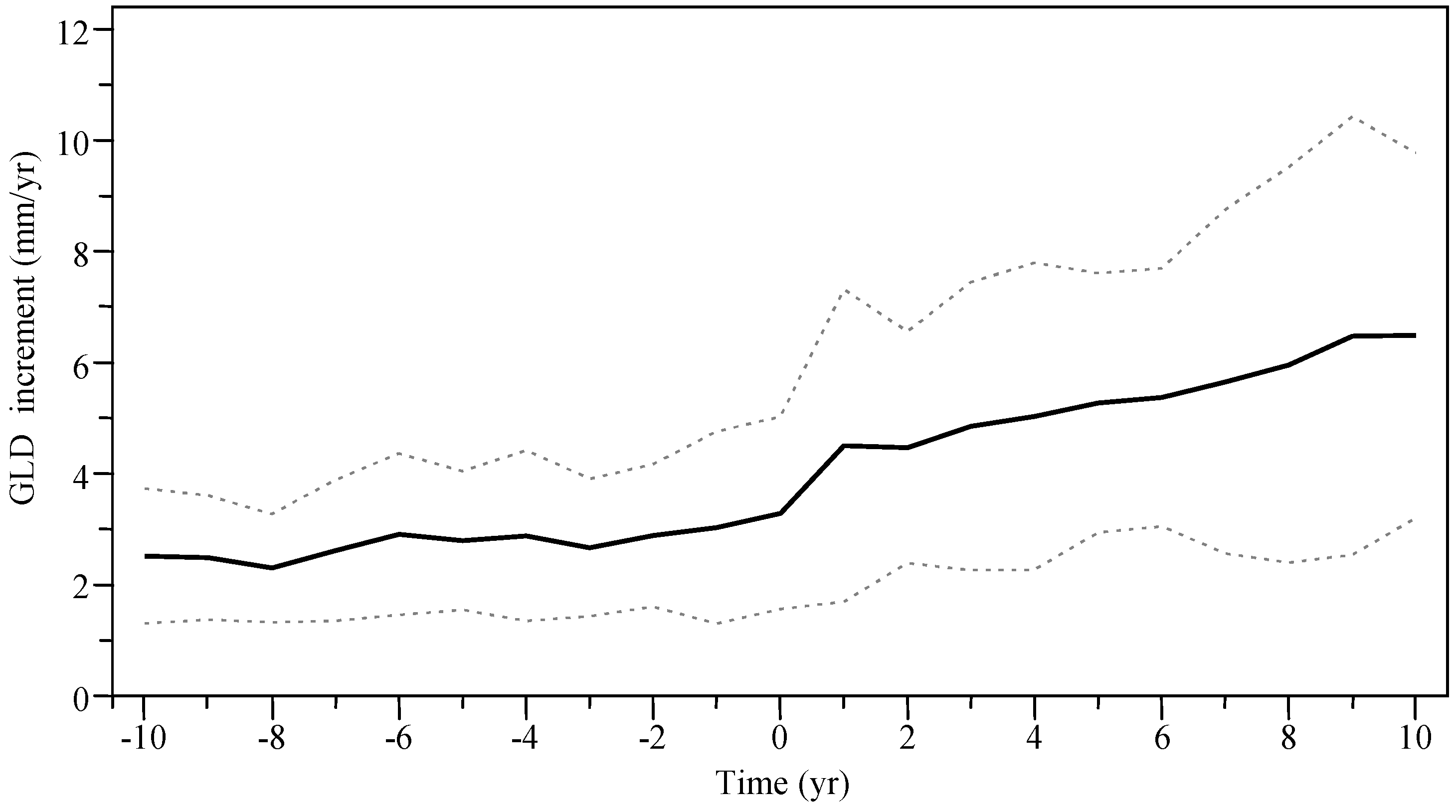
Before partial cutting, mean annual GLD increment ranged from 2.5 to 3.0 mm/yr and progressively increased with time since harvesting (mean slope = 0.06), up to 6.5 ± 3.3 mm/yr after 10 years (mean slope = 0.22;
Figure 6). In the first post-harvesting period (years 1 to 3), 43.1% of stems showed a significant increase in GLD increment compared to the pre-harvesting increment (
Figure 7). For subsequent 3-year periods (years 2 to 10), an additional 1.7% to 10.3% of stems showed an increase in GLD increment. After the first significant 3-year period, GLD increment of most stems remained greater than before harvesting for the rest of the data sequence analyzed. About 20% of stems showed no GLD increment response to partial harvesting; for 5.2% of stems, we observed at least one period where GLD increment significantly decreased compared to the pre-harvesting period.
Figure 6.
Mean annual GLD increment for stems ≥3 years at time of harvest (year 0) of the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site) (n = 58 stems). Dashed lines indicate standard error (SE).
Figure 6.
Mean annual GLD increment for stems ≥3 years at time of harvest (year 0) of the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site) (n = 58 stems). Dashed lines indicate standard error (SE).
Figure 7.
Distribution of northern white-cedar harvested stems showing a significant difference in GLD increment compared to the pre-cutting period (years −3 to −1) of the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site). Time of harvest = year 0.
Figure 7.
Distribution of northern white-cedar harvested stems showing a significant difference in GLD increment compared to the pre-cutting period (years −3 to −1) of the Outaouais Region (OR site) and Papineau-Labelle wildlife reserve (PL site). Time of harvest = year 0.
4. Discussion
Northern white-cedar can naturally regenerate with success under a wide variety of stand compositions and partial cutting regimes, with or without deer browsing. Despite the fact that the study spanned a very large area (
Figure 1) and that frequency and intensity of partial cutting differed between stands, all study sites had similar densities of seedlings <15 cm in 2005. Northern white-cedar regeneration is not particularly difficult to establish, provided there are seed trees in close proximity (<60 m) and the forest floor is disturbed, partially shaded, and constantly moist [
5]. The shade tolerance of northern white-cedar seedlings allows the formation of a seedling bank waiting for an opening in the canopy, as is the case for balsam fir [
20]. The observed density of small seedlings is in accordance with Forester
et al. [
26] who found a mean of 1800 northern white-cedar seedlings <22 cm by hectare in lowland northern white-cedar stands in Wisconsin, USA. Moreover, the density of these small seedlings was not affected per browsing pressure. This can be explained by the greater vulnerability to deer browsing of 30 to 130 cm seedlings, compared to smaller seedlings and taller saplings [
27]. Northern white-cedar is mostly a winter food, thus the snow cover generally protects the smallest seedlings, while the foliage of saplings >130cm is more difficult to reach, even considering that deer can browse to heights of up to 200 cm [
1,
3].
In contrast to establishment and GLD growth, height development and the ability of seedlings to reach larger sizes are strongly influenced by browsing pressure. In fact, the only site on which northern white-cedar seemed able to reach larger sizes was the OR site, where deer were absent. In addition, the PL and PEF sites, which were both subject to heavy browsing, had very similar height class distribution patterns, regardless of differences in climate, stand composition, and partial cutting regime. Browsing has already been suggested as a predominant factor to explain the low recruitment of northern white-cedar seedlings to larger size classes, but its impact can be very variable, depending on local herbivore densities [
1,
3,
26,
28]. Deer are the most important predators of northern white-cedar seedlings and saplings, especially in winter, and may cause mortality by recurrent browsing [
29]. Hare could also decrease the density of northern white-cedar regeneration, but do not seem to have played a major role in the present study, since northern white-cedar regeneration was quite comparable on the PL and PEF sites, which had different hare densities but high deer densities. Moose density was almost the same in all regions. Like hare, moose browses on northern white-cedar only when other food sources are scarce [
30], which was not the case in our mixedwood stands.
Recruitment of regeneration to larger sizes is also a function of height growth. No significant effect of browsing on height growth was detected, possibly because surviving stems were those that experienced low browsing pressure: on 95.8% of them, less than 50% of total foliage was consumed (
Table 3). The absence of older and taller seedlings on the PL and PEF sites could indicate that advance regeneration had reached a size where heavy browsing eliminated them instead of reducing the growth rate of the surviving stems [
6]. Browsing was frequent on the PEF; in 2005, it affected 90% of large seedlings (height > 30 cm) and small saplings [
6]. Depending on snow depth, stems may be browsed until they reach a height of 200 cm [
3]. Based on the age–height relationship in
Figure 4, northern white-cedar stems could therefore remain vulnerable to browsing damage for at least 36 years.
Northern white-cedar needs 11 years to reach a height of 30 cm, a value in the same range as balsam fir, white spruce, black spruce, and hemlock in northeastern North America (5 to 15 years) [
31,
32,
33]. Height growth rate increases once northern white-cedar regeneration gets older and larger. At a height of 50 cm, northern white-cedar stems are predicted to grow about 5 cm/yr, which is less than balsam fir (18 cm/yr), red spruce (12 cm/yr), and hardwood species [
12,
34]. In the presence of these species, northern white-cedar could then become overtopped. Northern white-cedar can survive extended periods of suppression [
8,
12,
35], and we know that merchantable stems of this species can respond well to release [
10]. Hence, the growth of northern white-cedar to maturity could involve a succession of suppression—release periods.
Height growth was a function of canopy transmittance, a factor that is heavily influenced by the partial cutting regime used and the harvesting intensity. Small northern white-cedar seedlings generally intercept a low percentage of light because of their overtopped position. This gives a false impression that northern white-cedar is a very slow-growing species. Nevertheless, the observed height increments (10 to 20 cm/yr with a canopy transmittance of 25% to 36%) are in the same range as those observed for balsam fir and red spruce [
34]. Moreover, our results agree with those obtained by Logan [
36] in a controlled environment, who predicted an increase of almost 6 cm/yr for northern white-cedar seedlings (>30 cm high) when light transmittance increased from 13% to 25%. Consequently, it seems reasonable to think that partial cutting could be used to favor northern white-cedar height growth.
In the same way, the GLD increment of half of the pre-established northern white-cedar regeneration responded quickly (≤5 years) and consistently to partial cutting. This result lends additional support to the use of partial cutting to release northern white-cedar advance regeneration, a suggestion made by Heitzman
et al. [
2] and Fraver [
35]. However, partial harvesting does not necessarily create uniform growing conditions, and sometimes leaves the local density of the overstory and understory unchanged. This is particularly true for diameter-limit cutting. In the present study, this single intervention did not release all overtopped stems, which may explain why the GLD increment of about 25% of stems did not increase after cutting. Another possible explanation is that pre-established stems of faster-growing tree species, such as birches, red maple, and balsam fir, together with the shrub stratum, were also present at the moment of partial cutting and benefited more than northern white-cedar from the additional growing space.
5. Conclusions
This study has shown that northern white-cedar can regenerate in a variety of stands and under many partial cutting regimes. However, recruitment to large seedling or sapling size classes can be quite variable and seriously compromised by high deer densities. This could have an important impact on future stand composition. According to our data, northern white-cedar remains vulnerable to browsing for more than 35 years, which poses major challenges regarding the renewal of the northern white-cedar component.
Northern white-cedar advance regeneration showed a rapid response in diameter increment (GLD) after harvesting. Height growth was strongly influenced by canopy light transmittance. Based on these results, we recommend that managers use partial cutting to open up the stand sufficiently to increase canopy transmittance on the forest floor to 20%–35% of full sunlight, and carefully retain pre-established northern white-cedar regeneration (both seedlings and saplings). This should be enough to induce a response for northern white-cedar, without stimulating the growth of other less shade-tolerant species growing in the same mixedwood stands. If deer density is high (>5.5 deer/km2, as in our study), measures limiting deer access could be necessary to ensure the continued development of the northern white-cedar regeneration.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





